Дендрит, аксон и синапс, строение нервной клетки
Клеточная мембрана
Этот элемент обеспечивает функцию барьера, отделяя внутреннюю среду от находящейся снаружи нейроглии. Тончайшая пленка состоит из двух слоев белковых молекул и находящихся между ними фосфолипидов. Строение мембраны нейрона предполагает наличие в ее структуре специфических рецепторов, отвечающих за узнавание раздражителей. Они обладают выборочной чувствительностью и при необходимости «включаются» при наличии контрагента. Связь внутренней и наружной сред происходит через канальцы, пропускающие ионы кальция или калия. При этом они открываются или закрываются под действием белковых рецепторов.
Благодаря мембране клетка имеет свой потенциал. При передаче его по цепочке происходит иннервация возбудимой ткани. Контакт мембран соседствующих нейронов происходит в синапсах. Поддержание постоянства внутренней среды – это важная составляющая жизнедеятельности любой клетки. И мембрана тонко регулирует концентрацию в цитоплазме молекул и заряженных ионов. При этом происходит транспорт их в необходимых количествах для протекания реакций метаболизма на оптимальном уровне.
Классификация
Структурная классификация
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Многие морфологи считают, что униполярные нейроны в теле человека и высших позвоночных не встречаются.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация
Морфологическое строение нейронов многообразно. При классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона;
- количество и характер ветвления отростков;
- длину аксона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов.
По количеству отростков выделяют следующие морфологические типы нейронов:
- униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
- псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
- биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
- мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
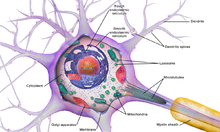
Строение нейронов
Схема нейрона
Тело клетки
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), ограниченной снаружи мембраной из липидного бислоя. Липиды состоят из гидрофильных головок и гидрофобных хвостов. Липиды располагаются гидрофобными хвостами друг к другу, образуя гидрофобный слой. Этот слой пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: в форме глобул на поверхности, на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20—30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии.(Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов).
В теле нейрона выявляется развитый синтетический аппарат, гранулярная эндоплазматическая сеть нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Основные статьи: Дендрит и Аксон
Схема строения нейрона
Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу.
Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом для образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Основная статья: Синапс
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г.
Литература
- Поляков Г. И., О принципах нейронной организации мозга, М: МГУ, 1965
- Косицын Н. С. Микроструктура дендритов и аксодендритических связей в центральной нервной системе. М.: Наука, 1976, 197 с.
- Немечек С. и др. Введение в нейробиологию, Avicennum: Прага, 1978, 400 c.
- Мозг (сборник статей: Д. Хьюбел, Ч. Стивенс, Э. Кэндел и дp. — выпуск журнала Scientific American (сентябрь 1979)). М. :Миp, 1980
- Савельева-Новосёлова Н. А., Савельев А. В. Устройство для моделирования нейрона. А. с. № 1436720, 1988
- Савельев А. В. Источники вариаций динамических свойств нервной системы на синаптическом уровне // журнал “Искусственный интеллект”, НАН Украины. — Донецк, Украина, 2006. — № 4. — С. 323—338.
Строение нейрона
На рисунке приведено строение нейрона. Он состоит из основного тела и ядра. От клеточного тела идет ответвление многочисленных волокон, которые именуются дендритами.
Мощные и длинные дендриты называются аксонами, которые в действительности намного длиннее, чем на картинке. Их протяженность варьируется от нескольких миллиметров до более метра.
Аксоны играют ведущую роль в передаче информации между нейронами и обеспечивают работу всей нервной системы.
Место соединения дендрита (аксона) с другим нейроном называется синапсом. Дендриты при наличии раздражителей могут разрастись настолько сильно, что станут улавливать импульсы от других клеток, что приводит к образованию новых синаптических связей.
Синаптические связи играют существенную роль в формировании личности человека. Так, личность с устоявшимся позитивным опытом будет смотреть на жизнь с любовью и надеждой, человек, у которого нейронные связи с негативным зарядом, станет со временем пессимистом.
Волокна
Вокруг нервных отростков независимо располагаются глиальные оболочки. В комплексе они формируют нервные волокна. Ответвления в них называются осевыми цилиндрами. Существуют безмиелиновые и миелиновые волокна. Они отличаются по строению глиальной оболочки. Безмиелиновые волокна имеют достаточно простое устройство. Подходящий к глиальной клетке осевой цилиндр прогибает ее цитолемму. Цитоплазма смыкается над ним и формирует мезаксон — двойную складку. Одна глиальная клетка может содержать несколько осевых цилиндров. Это «кабельные» волокна. Их ответвления могут переходить в расположенные по соседству глиальные клетки. Импульс проходит со скоростью 1-5 м/с. Волокна данного типа обнаруживаются в ходе эмбриогенеза и в постганглионарных участках вегетативной системы. Миелиновые сегменты толстые. Они расположены в соматической системе, иннервирующей мускулатуру скелета. Леммоциты (глиальные клетки) проходят последовательно, цепью. Они формируют тяж. В центре проходит осевой цилиндр. В глиальной оболочке присутствуют:
- Внутренний слой нервных клеток (миелиновый). Он считается основным. На некоторых участках между слоями цитолеммы присутствуют расширения, образующие миелиновые насечки.
- Периферический слой. В нем присутствуют органеллы и ядро – нейрилемма.
- Толстая базальная мембрана.
Внутреннее строение нейронов
Ядро нейрона
обычно крупное, округлое, с мелкодисперсным
хроматином, 1-3 крупными ядрышками. Это
отражает высокую интенсивность
процессов транскрипции в ядре нейрона.
Клеточная оболочка
нейрона способна генерировать и проводить
электрические импульсы. Это достигается
изменением локальной проницаемости
её ионных каналов для Na+ и К+, изменением
электрического потенциала и быстрым
перемещением его по цитолемме (волна
деполяризации, нервный импульс).
В цитоплазме нейронов
хорошо развиты все органоиды общего
назначения. Митохондрии
многочисленны и обеспечивают высокие
энергетические потребности нейрона,
связанные со значительной активностью
синтетических процессов, проведением
нервных импульсов, работой ионных
насосов. Они характеризуются быстрым
изнашиванием и обновлением (рис 8-3).
Комплекс
Гольджи очень
хорошо развит. Не случайно эта органелла
впервые была описана и демонстрируется
в курсе цитологии именно в нейронах.
При световой микроскопии он выявляется
в виде колечек, нитей, зёрнышек,
расположенных вокруг ядра (диктиосомы).
Многочисленные лизосомы
обеспечивают постоянное интенсивное
разрушение изнашиваемых компонентов
цитоплазмы нейрона (аутофагия).
Р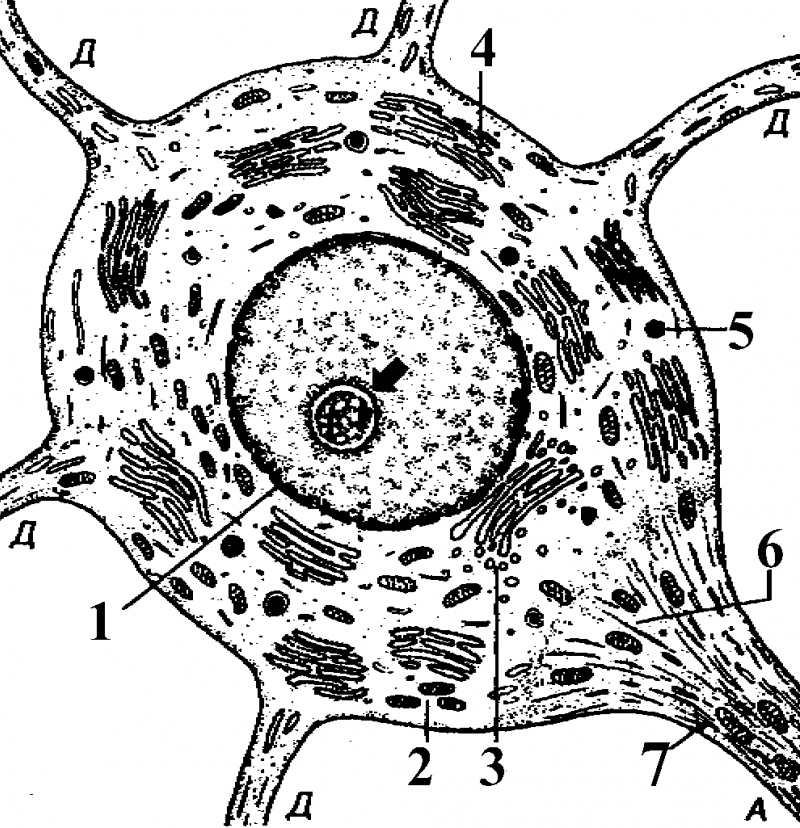 ис.
ис.
8-3. Ультрастуктурная организация
тела нейрона.
Д. Дендриты. А.
Аксон.
1. Ядро (ядрышко
показано стрелкой).
2. Митохондрии.
3. Комплекс
Гольджи.
4. Хроматофильная
субстанция (участки гранулярной
цитоплазмотической сети).
5. Лизосомы.
6. Аксонный
холмик.
7. Нейротрубочки,
нейрофиламенты.
(По В. Л. Быкову).
Для нормального
функционирования и обновления структур
нейрона в них должен быть хорошо развит
белоксинтезирующий аппарат (рис.
8-3). Гранулярная
цитоплазматическая сеть
в цитоплазме нейронов образует скопления,
которые хорошо окрашиваются основными
красителями и видны при световой
микроскопии в виде глыбок хроматофильного
вещества
(базофильное, или тигровое вещество,
субстанция Ниссля). Термин субстанция
Ниссля
сохранился в честь учёного Франца
Ниссля, впервые ее описавшего. Глыбки
хроматофильного вещества расположены
в перикарионах нейронов и дендритах,
но никогда не встречаются в аксонах,
где белоксинтезирующий аппарат развит
слабо (рис. 8-3). При длительном раздражении
или повреждении нейрона эти скопления
гранулярной цитоплазматической сети
распадаются на отдельные элементы, что
на светооптическом уровне проявляется
исчезновением субстанции Ниссля
(хроматолиз,
тигролиз).
Цитоскелет
нейронов хорошо развит, образует
трёхмерную сеть, представленную
нейрофиламентами (толщиной 6-10 нм) и
нейротрубочками (диаметром 20-30 нм).
Нейрофиламенты и нейротрубочки
связаны друг с другом поперечными
мостиками, при фиксации они склеиваются
в пучки толщиной 0,5-0,3 мкм, которые
окрашиваются солями серебра.На
светооптическом уровне они описаны под
названием нейрофибрилл.
Они образуют
сеть в перикарионах нейроцитов, а в
отростках лежат параллельно (рис. 8-2).
Цитоскелет поддерживает форму клеток,
а также обеспечивает транспортную
функцию – участвует в транспорте веществ
из перикариона в отростки (аксональный
транспорт).
Включения
в цитоплазме нейрона представлены
липидными каплями, гранулами
липофусцина
– «пигмента
старения» – жёлто-бурого цвета
липопротеидной природы. Они представляют
собой остаточные тельца (телолизосомы)
с продуктами непереваренных структур
нейрона. По-видимому, липофусцин
может накапливаться и в молодом возрасте,
при интенсивном функционировании и
повреждении нейронов. Кроме того, в
цитоплазме нейронов черной субстанции
и голубого пятна ствола мозга имеются
пигментные включения меланина.
Во многих нейронах головного мозга
встречаются включения гликогена.
Нейроны не способны к делению, и с
возрастом их число постепенно уменьшается
вследствие естественной гибели. При
дегенеративных заболеваниях (болезнь
Альцгеймера, Гентингтона, паркинсонизм)
интенсивность апоптоза возрастает и
количество нейронов в определённых
участках нервной системы резко
уменьшается.
Нервные клетки
Чтобы обеспечивать множественные связи, нейрон имеет особое строение. Кроме тела, в котором сосредоточены главные органеллы, присутствуют отростки. Часть их короткие (дендриты), обычно их несколько, другой (аксон) – он один, и его длина в отдельных структурах может достигать 1 метра.
Строение нервной клетки нейрона имеет такой вид, чтобы обеспечивать наилучший взаимообмен информацией. Дендриты сильно ветвятся (как крона дерева). Своими окончаниями они взаимодействуют с отростками других клеток. Место их стыка называют синапсом. Там происходит прием-передача импульса. Его направление: рецептор – дендрит – тело клетки (сома) – аксон – реагирующий орган или ткань.
Внутреннее строение нейрона по составу органелл сходно с другими структурными единицами тканей. В нем присутствует ядро и цитоплазма, ограниченная мембраной. Внутри располагаются митохондрии и рибосомы, микротрубочки, эндоплазматическая сеть, аппарат Гольджи.
Синапсы
С их помощью клетки нервной системы соединяются между собой. Существуют разные синапсы: аксо-соматические, -дендритические, -аксональные (главным образом тормозного типа). Также выделяют электрические и химические (первые выявляются достаточно редко в организме). В синапсах различают пост- и пресинаптическую части. Первая содержит мембрану, в которой присутствуют высокоспецифичные протеиновые (белковые) рецепторы. Они реагируют только на определенные медиаторы. Между пре- и постсинаптической частями расположена щель. Нервный импульс достигает первой и активирует особые пузырьки. Они переходят к пресинаптической мембране и попадают в щель. Оттуда они влияют на рецептор постсинаптической пленки. Это провоцирует ее деполяризацию, передающуюся, в свою очередь, посредством центрального отростка следующей нервной клетки. В химическом синапсе передача информации осуществляется только по одному направлению.
Развитие
Закладка нервной ткани происходит на третьей неделе эмбрионального периода. В это время формируется пластинка. Из нее развиваются:
- Олигодендроциты.
- Астроциты.
- Эпендимоциты.
- Макроглия.
В ходе дальнейшего эмбриогенеза нервная пластинка превращается в трубку. Во внутреннем слое ее стенки располагаются стволовые вентрикулярные элементы. Они пролиферируют и отходят кнаружи. В этой области часть клеток продолжает делиться. В результате они разделяются на спонгиобласты (компоненты микроглии), глиобласты и нейробласты. Из последних формируются нервные клетки. В стенке трубки выделяется 3 слоя:
- Внутренний (эпендимный).
- Средний (плащевой).
- Внешний (краевой) – представлен белым мозговым веществом.
На 20-24 неделе в краниальном сегменте трубки начинается образование пузырей, которые являются источником формирования головного мозга. Оставшиеся отделы служат для развития спинного мозга. От краев нервного желоба отходят клетки, участвующие в образовании гребня. Он располагается между эктодермой и трубкой. Из этих же клеток формируются ганглиозные пластинки, служащие основой для миелоцитов (пигментных кожных элементов), периферических нервных узлов, меланоцитов покрова, компонентов APUD-системы.
Классификация
Нейроны разделяют на виды в зависимости от типа медиатора (посредника проводящего импульса) выделяемого на окончаниях аксона. Это может быть холин, адреналин и пр. От места расположения в отделах ЦНС они могут относиться к соматическим нейронам или к вегетативным. Различают воспринимающие клетки (афферентные) и передающие обратные сигналы (эфферентные) в ответ на раздражение. Между ними могут находиться итернейроны, отвечающие за обмен информацией внутри ЦНС. По типу ответной реакции клетки могут тормозить возбуждение или, наоборот, повышать его.
По состоянию их готовности различают: «молчащие», которые начинают действовать (передают импульс) только при наличии определенного вида раздражения, и фоновые, что постоянно осуществляют мониторинг (непрерывная генерация сигналов). В зависимости от типа воспринимаемой от сенсоров информации меняется и строение нейрона. В этой связи их классифицируют на бимодальные, с относительно простым ответом на раздражение (два взаимосвязанных вида ощущения: укол и — как результат — боль, и полимодальные. Это более сложная структура – полимодальные нейроны (специфическая и неоднозначная реакция).
Что такое нейрон нейронные связи
В переводе с греческого нейрон, или как его еще называют неврон, означает «волокно», «нерв». Нейрон – это специфическая структура в нашем организме, которая отвечает за передачу внутри него любой информации, в быту называемая нервной клеткой.
Нейроны работают при помощи электрических сигналов и способствуют обработке мозгом поступающей информации для дальнейшей координации производимых телом действий.
Эти клетки являются составляющей частью нервной системы человека, предназначение которой состоит в том, чтобы собрать все сигналы, поступающие из вне или от собственного организма и принять решение о необходимости того или иного действия. Именно нейроны помогают справиться с такой задачей.
Каждый из нейронов имеет связь с огромным количеством таких же клеток, создаётся своеобразная «паутина», которая называется нейронной сетью. Посредством данной связи в организме передаются электрические и химические импульсы, приводящие всю нервную систему в состояние покоя либо, наоборот, возбуждения.
К примеру, человек столкнулся с неким значимым событием. Возникает электрохимический толчок (импульс) нейронов, приводящий к возбуждению неровной системы. У человека начинает чаще биться сердце, потеют руки или возникают другие физиологические реакции.
Мы рождаемся с заданным количеством нейронов, но связи между ними еще не сформированы. Нейронная сеть строится постепенно в результате поступающих из вне импульсов. Новые толчки формируют новые нейронные пути, именно по ним в течение жизни побежит аналогичная информация. Мозг воспринимает индивидуальный опыт каждого человека и реагирует на него. К примеру, ребенок, схватился за горячий утюг и отдернул руку. Так у него появилась новая нейронная связь.
Стабильная нейронная сеть выстраивается у ребенка уже к двум годам. Удивительно, но уже с этого возраста те клетки, которые не используются, начинают ослабевать. Но это никак не мешает развитию интеллекта. Наоборот, ребенок познает мир через уже устоявшиеся нейронные связи, а не анализирует бесцельно все вокруг.
Даже у такого малыша есть практический опыт, позволяющий отсекать ненужные действия и стремиться к полезным. Поэтому, например, так сложно отучить ребенка от груди — у него сформировалась крепкая нейронная связь между приложением к материнскому молоку и удовольствию, безопасности, спокойствию.
Познание нового опыта на протяжении всей жизни приводит к отмиранию ненужных нейронных связей и формированию новых и полезных. Этот процесс оптимизирует головной мозг наиболее эффективным для нас образом. Например, люди, проживающие в жарких странах, учатся жить в определенном климате, а северянам нужен совсем другой опыт для выживания.
Составляющие
Глиоцитов в системе в 5-10 раз больше, чем нервных клеток. Они выполняют разные функции: опорную, защитную, трофическую, стромальную, выделительную, всасывающую. Кроме этого, глиоциты обладают способностью к пролиферации. Эпендимоциты отличаются призматической формой. Они составляют первый слой, выстилают мозговые полости и центральный спинномозговой отдел. Клетки участвуют в продуцировании спинномозговой жидкости и обладают способностью всасывать ее. Базальная часть эпендимоцитов имеет коническую усеченную форму. Она переходит в длинный тонкий отросток, пронизывающий мозговое вещество. На его поверхности он формирует глиальную отграничительную мембрану. Астроциты представлены многоотросчатыми клетками. Они бывают:
- Протоплазматическими. Они расположены в сером мозговом веществе. Эти элементы отличаются наличием многочисленных коротких разветвлений, широких окончаний. Часть последних окружает кровеносные капиллярные сосуды, участвует в формировании гематоэнцефалического барьера. Другие отростки направлены к нейронным телам и по ним осуществляется перенос питательных веществ из крови. Они также обеспечивают защиту и изолируют синапсы.
- Волокнистыми (фиброзными). Эти клетки находятся в белом веществе. Их окончания слабоветвящиеся, длинные и тонкие. На концах у них присутствуют разветвления и формируются отграничительные мембраны.
Олиодендроциты представляют собой мелкие элементы с отходящими короткими хвостами, расположенными вокруг нейронов и их окончаний. Они формируют глиальную оболочку. Посредством нее передаются импульсы. На периферии эти клетки называют мантийными (леммоцитами). Микроглия является частью макрофагальной системы. Она представлена в виде мелких подвижных клеток с малоразветвленными короткими отростками. В элементах содержится светлое ядро. Они могут формироваться из кровяных моноцитов. Микроглия восстанавливает строение нервной клетки, подвергшейся повреждениям.
Нейроглия
Невроны не способны делиться, потому и появилось утверждение, что нервные клетки не восстанавливаются. Именно поэтому их следует оберегать с особой тщательностью. С основной функцией «няни» справляется нейроглия. Она находится между нервными волокнами.
Эти мелкие клетки отделяют нейроны друг от друга, удерживают их на своем месте. У них длинный список функций. Благодаря нейроглии сохраняется постоянная система установленных связей, обеспечивается расположение, питание и восстановление нейронов, выделяются отдельные медиаторы, фагоцитируется генетически чужое.
Таким образом, нейроглия выполняет ряд функций:
- опорную;
- разграничительную;
- регенераторную;
- трофическую;
- секреторную;
- защитную и т.д.
В ЦНС нейроны составляют серое вещество, а за границами мозга они скапливаются в специальные соединения, узлы – ганглии. Дендриты и аксоны создают белое вещество. На периферии именно благодаря этим отросткам строятся волокна, из которых и состоят нервы.
Строение нейрона
Плазматическая
мембранаокружает нервную клетку.
Она состоит из белковых и липидных
компонентов, находящихся в
жидкокристаллическом состоянии(модель
мозаичной мембраны): двуслойность
мембраны создается липидами, образующими
матрикс, в котрый частично или полностью
погружены белковые комплексы.
Плазматическая мембрана регулирует
обмен веществ между клеткой и ее средой,
а также служит структурной основой
электрической активности.
Ядроотделено
от цитоплазмы двумя мембранами, одна
из которых примыкает к ядру, а другая к
цитоплазме. Обе они местами сходятся,
образуя поры в ядерной оболочке, служащие
для транспорта веществ между ядром и
цитоплазмой. Ядро контролирует
дифференцировку нейрона в его конечную
форму, которая может быть очень сложной
и определяет характер межклеточных
связей. В ядре нейрона обычно находится
ядрышко.
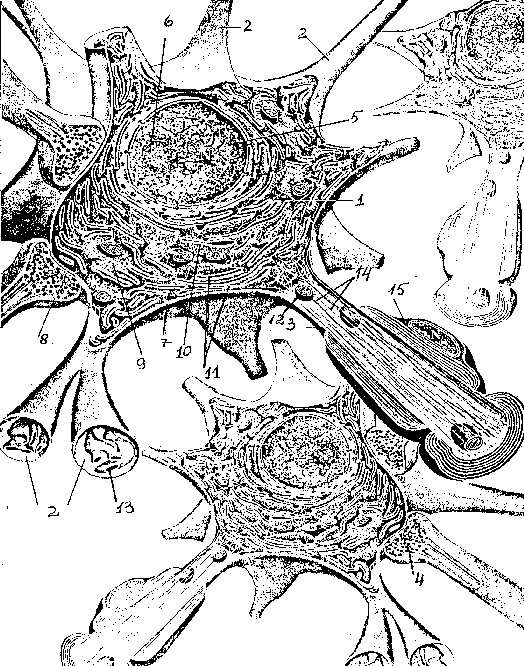
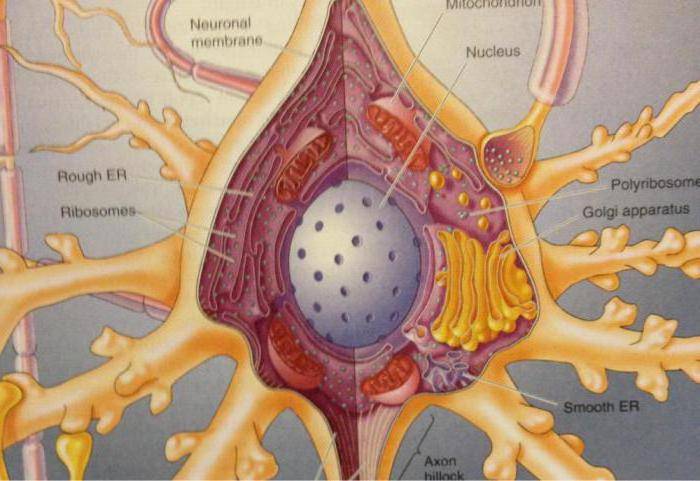
Рис. 1. Строение
нейрона (с изменениями по ):
1 — тело (сома), 2 —
дендрит, 3 — аксон, 4 — аксонная терминаль,
5 — ядро,
6 — ядрышко, 7 —
плазматическая мембрана, 8 — синапс, 9 —
рибосомы,
10 — шероховатый
(гранулярный) эндоплазматический
ретикулум,
11 — субстанция
Ниссля, 12 — митохондрии, 13 — агранулярный
эндоплазматический ретикулум, 14 —
микротрубочки и нейрофиламенты,
15
— миелиновая оболочка, образованная
шванновской клеткой
Рибосомы производят
элементы молекулярного аппарата для
большей части клеточных функций:
ферменты, белки-переносчики, рецепторы,
трансдукторы, сократительные и опорные
элементы, белки мембран. Часть рибосом
находится в цитоплазме в свободном
состоянии, другая часть прикрепляется
к обширной внутриклеточной мембранной
системе, являющейся продолжением
оболочки ядра и расходящейся по всей
соме в форме мембран, каналов, цистерн
и пузырьков (шероховатый эндоплазматический
ретикулум). В нейронах близ ядра
образуется характерное скопление
шероховатого эндоплазматического
ретикулума (субстанция Ниссля),
служащее местом интенсивного синтеза
белка.
Аппарат Гольджи
— система уплощенных мешочков, или
цистерн — имеет внутреннюю, формирующую,
сторону и наружную, выделяющую. От
последней отпочковываются пузырьки,
образующие секреторные гранулы. Функция
аппарата Гольджи в клетках состоит в
хранении, концентрировании и упаковке
секреторных белков. В нейронах он
представлен более мелкими скоплениями
цистерн и его функция менее ясна.
Лизосомы —заключенные в мембрану структуры,не
имеющие постоянной формы, — образуют
внутреннюю пищеварительную систему. У
взрослых особей в нейронах образуются
и накапливаютсялипофусциновые
гранулы, происходящие из лизосом. С
ними связывают процессы старения, а
также некоторые болезни.
Митохондрии
имеют гладкую наружную и складчатую
внутреннюю мембраны и являются местом
синтезааденозинтрифосфорной кислоты
(АТФ)— основного источника энергии
для клеточных процессов — в цикле
окисления глюкозы (у позвоночных).
Большинство нервных клеток лишено
способности запасать гликоген (полимер
глюкозы), что усиливает их зависимость
в отношении энергии от содержания в
крови кислорода и глюкозы.
Фибриллярные
структуры: микротрубочки(диаметр
20-30 нм),нейрофиламенты(10 нм) имикрофиламенты (5 нм). Микротрубочки
и нейрофиламенты участвуют во
внутриклеточном транспорте различных
веществ между телом клетки и отходящими
отростками. Микрофиламенты изобилуют
в растущих нервных отростках и,
по-видимому, управляют движениями
мембраны и текучестью подлежащей
цитоплазмы.
Синапс —функциональное соединение нейронов,
посредством которого происходит передача
электрических сигналов между клетками.Щелевой контактобеспечивает
электрический механизм связи между
нейронами(электрический синапс).
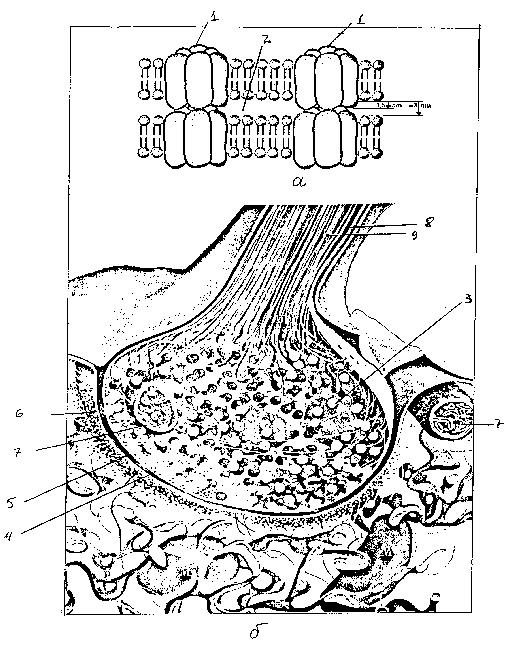
Рис. 2. Строение
синаптических контактов:
а
— щелевого контакта, б — химического
синапса (с изменениями по ):
1 — коннексон,
состоящий из 6 субъединиц, 2 — внеклеточное
пространство,
3 — синаптическая
везикула, 4 — пресинаптическая мембрана,
5 — синаптическая
щель, 6 —
постсинаптическая мембрана,7 — митохондрия,
8 — микротрубочка,
9
— нейрофиламенты
Химический синапсотличается ориентацией мембран в
направлении от нейрона к нейрону, что
проявляется в неодинаковой степени
уплотненности двух смежных мембран и
наличием группы небольших везикулвблизи синаптической щели. Такая
структура обеспечивает передачу сигнала
путем экзоцитоза медиатораиз
везикул.
Синапсы также
классифицируются в зависимости от того,
чем они образованы: аксо-соматические,
аксо-дендритные, аксо-аксонные и
дендро-дендритные.
Дендриты
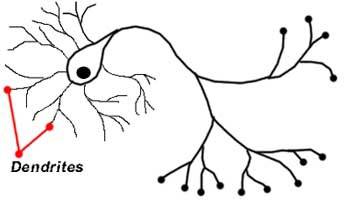
Дендриты – древовидные расширения в начале нейронов, которые служат для увеличения площади поверхности клетки. У многих нейронов их большое количество (тем не менее, встречаются и такие, у которых есть только один дендрит). Эти крошечные выступы получают информацию от других нейронов и передают её в виде импульсов к телу нейрона (соме). Место контакта нервных клеток, через которое передаются импульсы – химическим или электрическим путём, – называется синапсом.
Характеристики дендритов:
- Большинство нейронов имеют много дендритов
- Тем не менее, некоторые нейроны могут иметь только один дендрит
- Короткие и сильно разветвленные
- Участвует в передаче информации в тело клетки
Сома
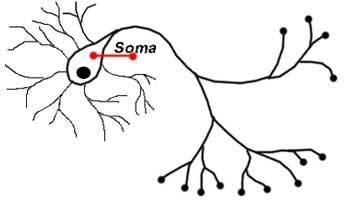
Сомой, или телом нейрона, называется место, где сигналы от дендритов аккумулируются и передаются дальше. Сома и ядро не играют активной роли в передаче нервных сигналов. Эти два образования служат скорее для поддержания жизнедеятельности нервной клетки и сохранения её работоспособности. Этой же цели служат митохондрии, которые обеспечивают клетки энергией, и аппарат Гольджи, который выводит продукты жизнедеятельности клеток за пределы клеточной мембраны.
Аксонный холмик
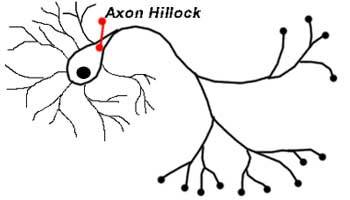
Аксонный холмик – участок сомы, от которого отходит аксон, – контролирует передачу нейроном импульсов. Именно тогда, когда общий уровень сигналов превышает пороговое значение холмика, он посылает импульс (известный, как потенциал действия) далее по аксону, к другой нервной клетке.
Аксон
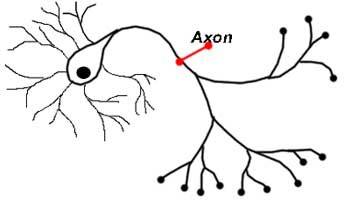
Аксон – это удлиненный отросток нейрона, который отвечает за передачу сигнала от одной клетки к другой. Чем больше аксон, тем быстрее он передаёт информацию. Некоторые аксоны покрыты специальным веществом (миелином), который выступает в качестве изолятора. Аксоны, покрытые миелиновой оболочкой, способны передавать информацию намного быстрее.
Характеристики Аксона:
- У большинства нейронов имеется только один аксон
- Участвует в передаче информации от тела клетки
- Может или не может иметь миелиновую оболочку
Терминальные ветви
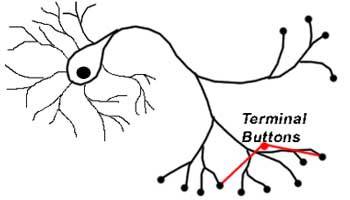
На конце Аксона расположены терминальные ветви – образования, которые отвечают за передачу сигналов к другим нейронам. В конце терминальных ветвей как раз и находятся синапсы. В них для передачи сигнала к другим нервным клеткам служат особые биологически активные химические вещества – нейромедиаторы.
Теги: мозг, нейрон, нервная система, строение
Есть что сказать? Оставть комментарий!:
Вывод
Физиология человека поражает своей слаженностью. Мозг стал величайшим творением эволюции. Если представлять организм в форме слаженной системы, то нейроны – это провода, по которым проходит сигнал от головного мозга и обратно. Их число огромно, они создают уникальную сеть в нашем организме. Ежесекундно по ней проходят тысячи сигналов. Это потрясающая система, которая позволяет не только функционировать организму, но и контактировать с окружающим миром.
Без невронов тело просто не сможет существовать, потому следует постоянно заботиться о состоянии своей нервной системы
Важно правильно питаться, избегать переутомления, стрессов, вовремя лечить заболевания
Введение в нейробиологию
7. Нейроны
Основными структурными особенностями нейронов являются перикария, дендриты и аксоны Нейроны содержат те же самые внутриклеточные компоненты, что и другие клетки Молекулярные маркеры могут использоваться для идентификации нейронов
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 85—86 миллиардов нейронов[1][2]. Нейроны могут соединяться один с другим, формируя биологические нейронные сети. Нейроны разделяют на рецепторные, эффекторные и вставочные.
Сложность и многообразие функций нервной системы определяются взаимодействием между нейронами. Это взаимодействие представляет собой набор различных сигналов, передаваемых между нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов. Ионы генерируют электрический заряд (потенциал действия), который движется по телу нейрона.
Важное значение для науки имело изобретение метода Гольджи в 1873 году, позволявшего окрашивать отдельные нейроны[3][4]. Термин «нейрон» (нем. Neuron) для обозначения нервных клеток введён Г. В. Вальдейером в 1891 году[5][6].

Строение нейронов
Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20—30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии.(Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в среднем в 10—50 раз больше, чем нейронов).
В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Аксоны и дендриты

Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу. Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторнойклеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г.
Not to be confused with Neutron.
| Neuron | |
|---|---|

Anatomy of a multipolar neuron |
|
| Identifiers | |
| MeSH | D009474 |
| NeuroLex ID | sao1417703748 |
| TA98 | A14.0.00.002 |
| TH | H2.00.06.1.00002 |
| FMA | 54527 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A neuron, neurone, or nerve cell is an electrically excitable cell that fires electric signals called action potentials. Neurons communicate with other cells via synapses — specialized connections that commonly use minute amounts of chemical neurotransmitters to pass the electric signal from the presynaptic neuron to the target cell through the synaptic gap. The neuron is the main component of nervous tissue in all animals except sponges and placozoa. Non-animals like plants and fungi do not have nerve cells.
Neurons are typically classified into three types based on their function. Sensory neurons respond to stimuli such as touch, sound, or light that affect the cells of the sensory organs, and they send signals to the spinal cord or brain. Motor neurons receive signals from the brain and spinal cord to control everything from muscle contractions to glandular output. Interneurons connect neurons to other neurons within the same region of the brain or spinal cord. When multiple neurons are functionally connected together, they form what is called a neural circuit.
A typical neuron consists of a cell body (soma), dendrites, and a single axon. The soma is a compact structure, and the axon and dendrites are filaments extruding from the soma. Dendrites typically branch profusely and extend a few hundred micrometers from the soma. The axon leaves the soma at a swelling called the axon hillock and travels for as far as 1 meter in humans or more in other species. It branches but usually maintains a constant diameter. At the farthest tip of the axon’s branches are axon terminals, where the neuron can transmit a signal across the synapse to another cell. Neurons may lack dendrites or have no axon. The term neurite is used to describe either a dendrite or an axon, particularly when the cell is undifferentiated.
Most neurons receive signals via the dendrites and soma and send out signals down the axon. At the majority of synapses, signals cross from the axon of one neuron to a dendrite of another. However, synapses can connect an axon to another axon or a dendrite to another dendrite.
The signaling process is partly electrical and partly chemical. Neurons are electrically excitable, due to maintenance of voltage gradients across their membranes. If the voltage changes by a large enough amount over a short interval, the neuron generates an all-or-nothing electrochemical pulse called an action potential. This potential travels rapidly along the axon and activates synaptic connections as it reaches them. Synaptic signals may be excitatory or inhibitory, increasing or reducing the net voltage that reaches the soma.
In most cases, neurons are generated by neural stem cells during brain development and childhood. Neurogenesis largely ceases during adulthood in most areas of the brain.
Nervous system[edit]
Schematic of an anatomically accurate single pyramidal neuron, the primaryious excitatory neuron of cerebral cortex, with a synaptic connection from an incoming axon onto a dendritic spine
Neurons are the primary components of the nervous system, along with the glial cells that give them structural and metabolic support.[1] The nervous system is made up of the central nervous system, which includes the brain and spinal cord, and the peripheral nervous system, which includes the autonomic and somatic nervous systems.[2] In vertebrates, the majority of neurons belong to the central nervous system, but some reside in peripheral ganglia, and many sensory neurons are situated in sensory organs such as the retina and cochlea.
Axons may bundle into fascicles that make up the nerves in the peripheral nervous system (like strands of wire make up cables). Bundles of axons in the central nervous system are called tracts.
Anatomy and histology[edit]

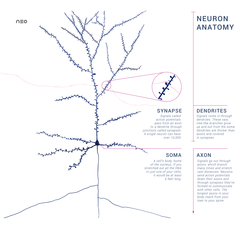
Diagram of the components of a neuron
Neurons are highly specialized for the processing and transmission of cellular signals. Given their diversity of functions performed in different parts of the nervous system, there is a wide variety in their shape, size, and electrochemical properties. For instance, the soma of a neuron can vary from 4 to 100 micrometers in diameter.[3]
- The soma is the body of the neuron. As it contains the nucleus, most protein synthesis occurs here. The nucleus can range from 3 to 18 micrometers in diameter.[4]
- The dendrites of a neuron are cellular extensions with many branches. This overall shape and structure are referred to metaphorically as a dendritic tree. This is where the majority of input to the neuron occurs via the dendritic spine.
- The axon is a finer, cable-like projection that can extend tens, hundreds, or even tens of thousands of times the diameter of the soma in length. The axon primarily carries nerve signals away from the soma and carries some types of information back to it. Many neurons have only one axon, but this axon may—and usually will—undergo extensive branching, enabling communication with many target cells. The part of the axon where it emerges from the soma is called the axon hillock. Besides being an anatomical structure, the axon hillock also has the greatest density of voltage-dependent sodium channels. This makes it the most easily excited part of the neuron and the spike initiation zone for the axon. In electrophysiological terms, it has the most negative threshold potential.
- While the axon and axon hillock are generally involved in information outflow, this region can also receive input from other neurons.
- The axon terminal is found at the end of the axon farthest from the soma and contains synapses. Synaptic boutons are specialized structures where neurotransmitter chemicals are released to communicate with target neurons. In addition to synaptic boutons at the axon terminal, a neuron may have en passant boutons, which are located along the length of the axon.
The accepted view of the neuron attributes dedicated functions to its various anatomical components; however, dendrites and axons often act in ways contrary to their so-called main function.[5]
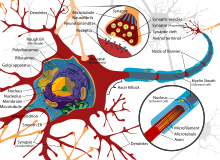
Diagram of a typical myelinated vertebrate motor neuron
Axons and dendrites in the central nervous system are typically only about one micrometer thick, while some in the peripheral nervous system are much thicker. The soma is usually about 10–25 micrometers in diameter and often is not much larger than the cell nucleus it contains. The longest axon of a human motor neuron can be over a meter long, reaching from the base of the spine to the toes.
Sensory neurons can have axons that run from the toes to the posterior column of the spinal cord, over 1.5 meters in adults. Giraffes have single axons several meters in length running along the entire length of their necks. Much of what is known about axonal function comes from studying the squid giant axon, an ideal experimental preparation because of its relatively immense size (0.5–1 millimeter thick, several centimeters long).
Fully differentiated neurons are permanently postmitotic[6] however, stem cells present in the adult brain may regenerate functional neurons throughout the life of an organism (see neurogenesis). Astrocytes are star-shaped glial cells. They have been observed to turn into neurons by virtue of their stem cell-like characteristic of pluripotency.
Membrane[edit]
Like all animal cells, the cell body of every neuron is enclosed by a plasma membrane, a bilayer of lipid molecules with many types of protein structures embedded in it.[7] A lipid bilayer is a powerful electrical insulator, but in neurons, many of the protein structures embedded in the membrane are electrically active. These include ion channels that permit electrically charged ions to flow across the membrane and ion pumps that chemically transport ions from one side of the membrane to the other. Most ion channels are permeable only to specific types of ions. Some ion channels are voltage gated, meaning that they can be switched between open and closed states by altering the voltage difference across the membrane. Others are chemically gated, meaning that they can be switched between open and closed states by interactions with chemicals that diffuse through the extracellular fluid. The ion materials include sodium, potassium, chloride, and calcium. The interactions between ion channels and ion pumps produce a voltage difference across the membrane, typically a bit less than 1/10 of a volt at baseline. This voltage has two functions: first, it provides a power source for an assortment of voltage-dependent protein machinery that is embedded in the membrane; second, it provides a basis for electrical signal transmission between different parts of the membrane.
Histology and internal structure[edit]

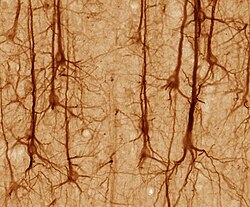
Golgi-stained neurons in human hippocampal tissue

Actin filaments in a mouse cortical neuron in culture
Numerous microscopic clumps called Nissl bodies (or Nissl substance) are seen when nerve cell bodies are stained with a basophilic («base-loving») dye. These structures consist of rough endoplasmic reticulum and associated ribosomal RNA. Named after German psychiatrist and neuropathologist Franz Nissl (1860–1919), they are involved in protein synthesis and their prominence can be explained by the fact that nerve cells are very metabolically active. Basophilic dyes such as aniline or (weakly) haematoxylin[8] highlight negatively charged components, and so bind to the phosphate backbone of the ribosomal RNA.
The cell body of a neuron is supported by a complex mesh of structural proteins called neurofilaments, which together with neurotubules (neuronal microtubules) are assembled into larger neurofibrils.[9] Some neurons also contain pigment granules, such as neuromelanin (a brownish-black pigment that is byproduct of synthesis of catecholamines), and lipofuscin (a yellowish-brown pigment), both of which accumulate with age.[10][11][12] Other structural proteins that are important for neuronal function are actin and the tubulin of microtubules. Class III β-tubulin is found almost exclusively in neurons. Actin is predominately found at the tips of axons and dendrites during neuronal development. There the actin dynamics can be modulated via an interplay with microtubule.[13]
There are different internal structural characteristics between axons and dendrites. Typical axons almost never contain ribosomes, except some in the initial segment. Dendrites contain granular endoplasmic reticulum or ribosomes, in diminishing amounts as the distance from the cell body increases.
Classification[edit]

Neurons vary in shape and size and can be classified by their morphology and function.[15] The anatomist Camillo Golgi grouped neurons into two types; type I with long axons used to move signals over long distances and type II with short axons, which can often be confused with dendrites. Type I cells can be further classified by the location of the soma. The basic morphology of type I neurons, represented by spinal motor neurons, consists of a cell body called the soma and a long thin axon covered by a myelin sheath. The dendritic tree wraps around the cell body and receives signals from other neurons. The end of the axon has branching axon terminals that release neurotransmitters into a gap called the synaptic cleft between the terminals and the dendrites of the next neuron.[citation needed]
Structural classification[edit]
Polarity[edit]
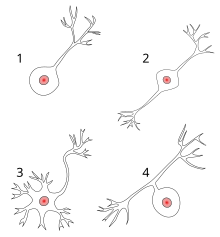
Most neurons can be anatomically characterized as:[citation needed]
- Unipolar: single process
- Bipolar: 1 axon and 1 dendrite
- Multipolar: 1 axon and 2 or more dendrites
- Golgi I: neurons with long-projecting axonal processes; examples are pyramidal cells, Purkinje cells, and anterior horn cells
- Golgi II: neurons whose axonal process projects locally; the best example is the granule cell
- Anaxonic: where the axon cannot be distinguished from the dendrite(s)
- Pseudounipolar: 1 process which then serves as both an axon and a dendrite
Other[edit]
Some unique neuronal types can be identified according to their location in the nervous system and distinct shape. Some examples are:[citation needed]
- Basket cells, interneurons that form a dense plexus of terminals around the soma of target cells, found in the cortex and cerebellum
- Betz cells, large motor neurons
- Lugaro cells, interneurons of the cerebellum
- Medium spiny neurons, most neurons in the corpus striatum
- Purkinje cells, huge neurons in the cerebellum, a type of Golgi I multipolar neuron
- Pyramidal cells, neurons with triangular soma, a type of Golgi I
- Renshaw cells, neurons with both ends linked to alpha motor neurons
- Unipolar brush cells, interneurons with unique dendrite ending in a brush-like tuft
- Granule cells, a type of Golgi II neuron
- Anterior horn cells, motoneurons located in the spinal cord
- Spindle cells, interneurons that connect widely separated areas of the brain
Functional classification[edit]
Direction[edit]
- Afferent neurons convey information from tissues and organs into the central nervous system and are also called sensory neurons.
- Efferent neurons (motor neurons) transmit signals from the central nervous system to the effector cells.
- Interneurons connect neurons within specific regions of the central nervous system.
Afferent and efferent also refer generally to neurons that, respectively, bring information to or send information from the brain.
Action on other neurons[edit]
A neuron affects other neurons by releasing a neurotransmitter that binds to chemical receptors. The effect upon the postsynaptic neuron is determined by the type of receptor that is activated, not by the presynaptic neuron or by the neurotransmitter. A neurotransmitter can be thought of as a key, and a receptor as a lock: the same neurotransmitter can activate multiple types of receptors. Receptors can be classified broadly as excitatory (causing an increase in firing rate), inhibitory (causing a decrease in firing rate), or modulatory (causing long-lasting effects not directly related to firing rate).[citation needed]
The two most common (90%+) neurotransmitters in the brain, glutamate and GABA, have largely consistent actions. Glutamate acts on several types of receptors, and has effects that are excitatory at ionotropic receptors and a modulatory effect at metabotropic receptors. Similarly, GABA acts on several types of receptors, but all of them have inhibitory effects (in adult animals, at least). Because of this consistency, it is common for neuroscientists to refer to cells that release glutamate as «excitatory neurons», and cells that release GABA as «inhibitory neurons». Some other types of neurons have consistent effects, for example, «excitatory» motor neurons in the spinal cord that release acetylcholine, and «inhibitory» spinal neurons that release glycine.[citation needed]
The distinction between excitatory and inhibitory neurotransmitters is not absolute. Rather, it depends on the class of chemical receptors present on the postsynaptic neuron. In principle, a single neuron, releasing a single neurotransmitter, can have excitatory effects on some targets, inhibitory effects on others, and modulatory effects on others still. For example, photoreceptor cells in the retina constantly release the neurotransmitter glutamate in the absence of light. So-called OFF bipolar cells are, like most neurons, excited by the released glutamate. However, neighboring target neurons called ON bipolar cells are instead inhibited by glutamate, because they lack typical ionotropic glutamate receptors and instead express a class of inhibitory metabotropic glutamate receptors.[16] When light is present, the photoreceptors cease releasing glutamate, which relieves the ON bipolar cells from inhibition, activating them; this simultaneously removes the excitation from the OFF bipolar cells, silencing them.[citation needed]
It is possible to identify the type of inhibitory effect a presynaptic neuron will have on a postsynaptic neuron, based on the proteins the presynaptic neuron expresses. Parvalbumin-expressing neurons typically dampen the output signal of the postsynaptic neuron in the visual cortex, whereas somatostatin-expressing neurons typically block dendritic inputs to the postsynaptic neuron.[17]
Discharge patterns[edit]
Neurons have intrinsic electroresponsive properties like intrinsic transmembrane voltage oscillatory patterns.[18] So neurons can be classified according to their electrophysiological characteristics:
- Tonic or regular spiking. Some neurons are typically constantly (tonically) active, typically firing at a constant frequency. Example: interneurons in neurostriatum.
- Phasic or bursting. Neurons that fire in bursts are called phasic.
- Fast spiking. Some neurons are notable for their high firing rates, for example some types of cortical inhibitory interneurons, cells in globus pallidus, retinal ganglion cells.[19][20]
Neurotransmitter[edit]
![]()
Synaptic vesicles containing neurotransmitters
Neurotransmitters are chemical messengers passed from one neuron to another neuron or to a muscle cell or gland cell.
- Cholinergic neurons – acetylcholine. Acetylcholine is released from presynaptic neurons into the synaptic cleft. It acts as a ligand for both ligand-gated ion channels and metabotropic (GPCRs) muscarinic receptors. Nicotinic receptors are pentameric ligand-gated ion channels composed of alpha and beta subunits that bind nicotine. Ligand binding opens the channel causing influx of Na+ depolarization and increases the probability of presynaptic neurotransmitter release. Acetylcholine is synthesized from choline and acetyl coenzyme A.
- Adrenergic neurons – noradrenaline. Noradrenaline (norepinephrine) is released from most postganglionic neurons in the sympathetic nervous system onto two sets of GPCRs: alpha adrenoceptors and beta adrenoceptors. Noradrenaline is one of the three common catecholamine neurotransmitter, and the most prevalent of them in the peripheral nervous system; as with other catecholamines, it is synthesised from tyrosine.
- GABAergic neurons – gamma aminobutyric acid. GABA is one of two neuroinhibitors in the central nervous system (CNS), along with glycine. GABA has a homologous function to ACh, gating anion channels that allow Cl− ions to enter the post synaptic neuron. Cl− causes hyperpolarization within the neuron, decreasing the probability of an action potential firing as the voltage becomes more negative (for an action potential to fire, a positive voltage threshold must be reached). GABA is synthesized from glutamate neurotransmitters by the enzyme glutamate decarboxylase.
- Glutamatergic neurons – glutamate. Glutamate is one of two primary excitatory amino acid neurotransmitters, along with aspartate. Glutamate receptors are one of four categories, three of which are ligand-gated ion channels and one of which is a G-protein coupled receptor (often referred to as GPCR).
-
- AMPA and Kainate receptors function as cation channels permeable to Na+ cation channels mediating fast excitatory synaptic transmission.
- NMDA receptors are another cation channel that is more permeable to Ca2+. The function of NMDA receptors depend on glycine receptor binding as a co-agonist within the channel pore. NMDA receptors do not function without both ligands present.
- Metabotropic receptors, GPCRs modulate synaptic transmission and postsynaptic excitability.
- Glutamate can cause excitotoxicity when blood flow to the brain is interrupted, resulting in brain damage. When blood flow is suppressed, glutamate is released from presynaptic neurons, causing greater NMDA and AMPA receptor activation than normal outside of stress conditions, leading to elevated Ca2+ and Na+ entering the post synaptic neuron and cell damage. Glutamate is synthesized from the amino acid glutamine by the enzyme glutamate synthase.
- Dopaminergic neurons—dopamine. Dopamine is a neurotransmitter that acts on D1 type (D1 and D5) Gs-coupled receptors, which increase cAMP and PKA, and D2 type (D2, D3, and D4) receptors, which activate Gi-coupled receptors that decrease cAMP and PKA. Dopamine is connected to mood and behavior and modulates both pre- and post-synaptic neurotransmission. Loss of dopamine neurons in the substantia nigra has been linked to Parkinson’s disease. Dopamine is synthesized from the amino acid tyrosine. Tyrosine is catalyzed into levodopa (or L-DOPA) by tyrosine hydroxlase, and levodopa is then converted into dopamine by the aromatic amino acid decarboxylase.
- Serotonergic neurons—serotonin. Serotonin (5-Hydroxytryptamine, 5-HT) can act as excitatory or inhibitory. Of its four 5-HT receptor classes, 3 are GPCR and 1 is a ligand-gated cation channel. Serotonin is synthesized from tryptophan by tryptophan hydroxylase, and then further by decarboxylase. A lack of 5-HT at postsynaptic neurons has been linked to depression. Drugs that block the presynaptic serotonin transporter are used for treatment, such as Prozac and Zoloft.
- Purinergic neurons—ATP. ATP is a neurotransmitter acting at both ligand-gated ion channels (P2X receptors) and GPCRs (P2Y) receptors. ATP is, however, best known as a cotransmitter. Such purinergic signalling can also be mediated by other purines like adenosine, which particularly acts at P2Y receptors.
- Histaminergic neurons—histamine. Histamine is a monoamine neurotransmitter and neuromodulator. Histamine-producing neurons are found in the tuberomammillary nucleus of the hypothalamus.[21] Histamine is involved in arousal and regulating sleep/wake behaviors.
Multimodel classification[edit]
Since 2012 there has been a push from the cellular and computational neuroscience community to come up with a universal classification of neurons that will apply to all neurons in the brain as well as across species. This is done by considering the three essential qualities of all neurons: electrophysiology, morphology, and the individual transcriptome of the cells. Besides being universal this classification has the advantage of being able to classify astrocytes as well. A method called Patch-Seq in which all three qualities can be measured at once is used extensively by the Allen Institute for Brain Science.[22]
Connectivity[edit]

A signal propagating down an axon to the cell body and dendrites of the next cell

Neurons communicate with each other via synapses, where either the axon terminal of one cell contacts another neuron’s dendrite, soma or, less commonly, axon. Neurons such as Purkinje cells in the cerebellum can have over 1000 dendritic branches, making connections with tens of thousands of other cells; other neurons, such as the magnocellular neurons of the supraoptic nucleus, have only one or two dendrites, each of which receives thousands of synapses.
Synapses can be excitatory or inhibitory, either increasing or decreasing activity in the target neuron, respectively. Some neurons also communicate via electrical synapses, which are direct, electrically conductive junctions between cells.[23]
When an action potential reaches the axon terminal, it opens voltage-gated calcium channels, allowing calcium ions to enter the terminal. Calcium causes synaptic vesicles filled with neurotransmitter molecules to fuse with the membrane, releasing their contents into the synaptic cleft. The neurotransmitters diffuse across the synaptic cleft and activate receptors on the postsynaptic neuron. High cytosolic calcium in the axon terminal triggers mitochondrial calcium uptake, which, in turn, activates mitochondrial energy metabolism to produce ATP to support continuous neurotransmission.[24]
An autapse is a synapse in which a neuron’s axon connects to its own dendrites.
The human brain has some 8.6 x 1010 (eighty six billion) neurons.[25] Each neuron has on average 7,000 synaptic connections to other neurons. It has been estimated that the brain of a three-year-old child has about 1015 synapses (1 quadrillion). This number declines with age, stabilizing by adulthood. Estimates vary for an adult, ranging from 1014 to 5 x 1014 synapses (100 to 500 trillion).[26]
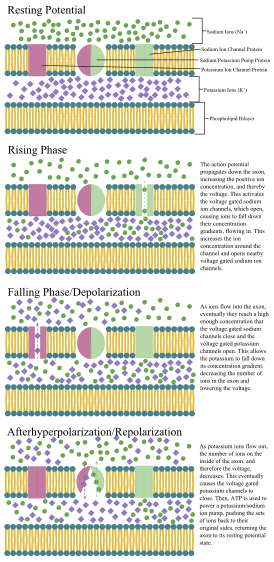
An annotated diagram of the stages of an action potential propagating down an axon including the role of ion concentration and pump and channel proteins
Nonelectrochemical signaling[edit]
Beyond electrical and chemical signaling, studies suggest neurons in healthy human brains can also communicate through:
- force generated by the enlargement of dendritic spines[27]
- the transfer of proteins – transneuronally transported proteins (TNTPs)[28][29]
They can also get modulated by input from the environment and hormones released from other parts of the organism,[30] which could be influenced more or less directly by neurons. This also applies to neurotrophins such as BDNF. The gut microbiome is also connected with the brain.[31]
Neurons also communicate with microglia, the brain’s main immune cells via specialised contact sites, called «somatic junctions». These connections enable microglia to constantly monitor and regulate neuronal functions, and exert neuroprotection, when needed.[32]
Mechanisms for propagating action potentials[edit]
In 1937 John Zachary Young suggested that the squid giant axon could be used to study neuronal electrical properties.[33] It is larger than but similar to human neurons, making it easier to study. By inserting electrodes into the squid giant axons, accurate measurements were made of the membrane potential.
The cell membrane of the axon and soma contain voltage-gated ion channels that allow the neuron to generate and propagate an electrical signal (an action potential). Some neurons also generate subthreshold membrane potential oscillations. These signals are generated and propagated by charge-carrying ions including sodium (Na+), potassium (K+), chloride (Cl−), and calcium (Ca2+).
Several stimuli can activate a neuron leading to electrical activity, including pressure, stretch, chemical transmitters, and changes of the electric potential across the cell membrane.[34] Stimuli cause specific ion-channels within the cell membrane to open, leading to a flow of ions through the cell membrane, changing the membrane potential. Neurons must maintain the specific electrical properties that define their neuron type.[35]
Thin neurons and axons require less metabolic expense to produce and carry action potentials, but thicker axons convey impulses more rapidly. To minimize metabolic expense while maintaining rapid conduction, many neurons have insulating sheaths of myelin around their axons. The sheaths are formed by glial cells: oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system. The sheath enables action potentials to travel faster than in unmyelinated axons of the same diameter, whilst using less energy. The myelin sheath in peripheral nerves normally runs along the axon in sections about 1 mm long, punctuated by unsheathed nodes of Ranvier, which contain a high density of voltage-gated ion channels. Multiple sclerosis is a neurological disorder that results from demyelination of axons in the central nervous system.
Some neurons do not generate action potentials, but instead generate a graded electrical signal, which in turn causes graded neurotransmitter release. Such non-spiking neurons tend to be sensory neurons or interneurons, because they cannot carry signals long distances.
Neural coding[edit]
Neural coding is concerned with how sensory and other information is represented in the brain by neurons. The main goal of studying neural coding is to characterize the relationship between the stimulus and the individual or ensemble neuronal responses, and the relationships among the electrical activities of the neurons within the ensemble.[36] It is thought that neurons can encode both digital and analog information.[37]
All-or-none principle[edit]
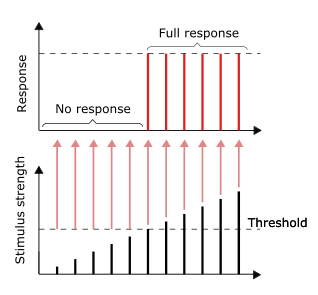
As long as the stimulus reaches the threshold, the full response would be given. Larger stimulus does not result in a larger response, vice versa.[38]: 31
The conduction of nerve impulses is an example of an all-or-none response. In other words, if a neuron responds at all, then it must respond completely. Greater intensity of stimulation, like brighter image/louder sound, does not produce a stronger signal, but can increase firing frequency.[38]: 31 Receptors respond in different ways to stimuli. Slowly adapting or tonic receptors respond to steady stimulus and produce a steady rate of firing. Tonic receptors most often respond to increased intensity of stimulus by increasing their firing frequency, usually as a power function of stimulus plotted against impulses per second. This can be likened to an intrinsic property of light where greater intensity of a specific frequency (color) requires more photons, as the photons can’t become «stronger» for a specific frequency.
Other receptor types include quickly adapting or phasic receptors, where firing decreases or stops with steady stimulus; examples include skin which, when touched causes neurons to fire, but if the object maintains even pressure, the neurons stop firing. The neurons of the skin and muscles that are responsive to pressure and vibration have filtering accessory structures that aid their function.
The pacinian corpuscle is one such structure. It has concentric layers like an onion, which form around the axon terminal. When pressure is applied and the corpuscle is deformed, mechanical stimulus is transferred to the axon, which fires. If the pressure is steady, stimulus ends; thus, typically these neurons respond with a transient depolarization during the initial deformation and again when the pressure is removed, which causes the corpuscle to change shape again. Other types of adaptation are important in extending the function of a number of other neurons.[39]
Etymology and spelling[edit]
The German anatomist Heinrich Wilhelm Waldeyer introduced the term neuron in 1891,[40] based on the ancient Greek νεῦρον neuron ‘sinew, cord, nerve’.[41]
The word was adopted in French with the spelling neurone. That spelling was also used by many writers in English,[42] but has now become rare in American usage and uncommon in British usage.[43][41]
History[edit]

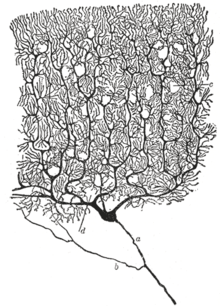
The neuron’s place as the primary functional unit of the nervous system was first recognized in the late 19th century through the work of the Spanish anatomist Santiago Ramón y Cajal.[44]
To make the structure of individual neurons visible, Ramón y Cajal improved a silver staining process that had been developed by Camillo Golgi.[44] The improved process involves a technique called «double impregnation» and is still in use.
In 1888 Ramón y Cajal published a paper about the bird cerebellum. In this paper, he stated that he could not find evidence for anastomosis between axons and dendrites and called each nervous element «an absolutely autonomous canton.»[44][40] This became known as the neuron doctrine, one of the central tenets of modern neuroscience.[44]
In 1891, the German anatomist Heinrich Wilhelm Waldeyer wrote a highly influential review of the neuron doctrine in which he introduced the term neuron to describe the anatomical and physiological unit of the nervous system.[45][46]
The silver impregnation stains are a useful method for neuroanatomical investigations because, for reasons unknown, it stains only a small percentage of cells in a tissue, exposing the complete micro structure of individual neurons without much overlap from other cells.[47]
Neuron doctrine[edit]

The neuron doctrine is the now fundamental idea that neurons are the basic structural and functional units of the nervous system. The theory was put forward by Santiago Ramón y Cajal in the late 19th century. It held that neurons are discrete cells (not connected in a meshwork), acting as metabolically distinct units.
Later discoveries yielded refinements to the doctrine. For example, glial cells, which are non-neuronal, play an essential role in information processing.[48] Also, electrical synapses are more common than previously thought,[49] comprising direct, cytoplasmic connections between neurons. In fact, neurons can form even tighter couplings: the squid giant axon arises from the fusion of multiple axons.[50]
Ramón y Cajal also postulated the Law of Dynamic Polarization, which states that a neuron receives signals at its dendrites and cell body and transmits them, as action potentials, along the axon in one direction: away from the cell body.[51] The Law of Dynamic Polarization has important exceptions; dendrites can serve as synaptic output sites of neurons[52] and axons can receive synaptic inputs.[53]
Compartmental modelling of neurons[edit]
Although neurons are often described of as «fundamental units» of the brain, they perform internal computations. Neurons integrate input within dendrites, and this complexity is lost in models that assume neurons to be a fundamental unit. Dendritic branches can be modeled as spatial compartments, whose activity is related due to passive membrane properties, but may also be different depending on input from synapses. Compartmental modelling of dendrites is especially helpful for understanding the behavior of neurons that are too small to record with electrodes, as is the case for Drosophila melanogaster.[54]
Neurons in the brain[edit]
The number of neurons in the brain varies dramatically from species to species.[55] In a human, there are an estimated 10–20 billion neurons in the cerebral cortex and 55–70 billion neurons in the cerebellum.[56] By contrast, the nematode worm Caenorhabditis elegans has just 302 neurons, making it an ideal model organism as scientists have been able to map all of its neurons. The fruit fly Drosophila melanogaster, a common subject in biological experiments, has around 100,000 neurons and exhibits many complex behaviors. Many properties of neurons, from the type of neurotransmitters used to ion channel composition, are maintained across species, allowing scientists to study processes occurring in more complex organisms in much simpler experimental systems.
Neurological disorders[edit]
Charcot–Marie–Tooth disease (CMT) is a heterogeneous inherited disorder of nerves (neuropathy) that is characterized by loss of muscle tissue and touch sensation, predominantly in the feet and legs extending to the hands and arms in advanced stages. Presently incurable, this disease is one of the most common inherited neurological disorders, affecting 36 in 100,000 people.[57]
Alzheimer’s disease (AD), also known simply as Alzheimer’s, is a neurodegenerative disease characterized by progressive cognitive deterioration, together with declining activities of daily living and neuropsychiatric symptoms or behavioral changes.[58] The most striking early symptom is loss of short-term memory (amnesia), which usually manifests as minor forgetfulness that becomes steadily more pronounced with illness progression, with relative preservation of older memories. As the disorder progresses, cognitive (intellectual) impairment extends to the domains of language (aphasia), skilled movements (apraxia), and recognition (agnosia), and functions such as decision-making and planning become impaired.[59][60]
Parkinson’s disease (PD), also known as Parkinsons, is a degenerative disorder of the central nervous system that often impairs motor skills and speech.[61] Parkinson’s disease belongs to a group of conditions called movement disorders.[62] It is characterized by muscle rigidity, tremor, a slowing of physical movement (bradykinesia), and in extreme cases, a loss of physical movement (akinesia). The primary symptoms are the results of decreased stimulation of the motor cortex by the basal ganglia, normally caused by the insufficient formation and action of dopamine, which is produced in the dopaminergic neurons of the brain. Secondary symptoms may include high level cognitive dysfunction and subtle language problems. PD is both chronic and progressive.
Myasthenia gravis is a neuromuscular disease leading to fluctuating muscle weakness and fatigability during simple activities. Weakness is typically caused by circulating antibodies that block acetylcholine receptors at the post-synaptic neuromuscular junction, inhibiting the stimulative effect of the neurotransmitter acetylcholine. Myasthenia is treated with immunosuppressants, cholinesterase inhibitors and, in selected cases, thymectomy.
Demyelination[edit]

Demyelination is the act of demyelinating, or the loss of the myelin sheath insulating the nerves. When myelin degrades, conduction of signals along the nerve can be impaired or lost, and the nerve eventually withers. This leads to certain neurodegenerative disorders like multiple sclerosis and chronic inflammatory demyelinating polyneuropathy.
Axonal degeneration[edit]
Although most injury responses include a calcium influx signaling to promote resealing of severed parts, axonal injuries initially lead to acute axonal degeneration, which is the rapid separation of the proximal and distal ends, occurring within 30 minutes of injury.[63] Degeneration follows with swelling of the axolemma, and eventually leads to bead-like formation. Granular disintegration of the axonal cytoskeleton and inner organelles occurs after axolemma degradation. Early changes include accumulation of mitochondria in the paranodal regions at the site of injury. Endoplasmic reticulum degrades and mitochondria swell up and eventually disintegrate. The disintegration is dependent on ubiquitin and calpain proteases (caused by the influx of calcium ion), suggesting that axonal degeneration is an active process that produces complete fragmentation. The process takes about roughly 24 hours in the PNS and longer in the CNS. The signaling pathways leading to axolemma degeneration are unknown.
Neurogenesis[edit]
Neurons are born through the process of neurogenesis, in which neural stem cells divide to produce differentiated neurons. Once fully differentiated neurons are formed, they are no longer capable of undergoing mitosis. Neurogenesis primarily occurs in the embryo of most organisms.
Adult neurogenesis can occur and studies of the age of human neurons suggest that this process occurs only for a minority of cells, and that the vast majority of neurons in the neocortex forms before birth and persists without replacement. The extent to which adult neurogenesis exists in humans, and its contribution to cognition are controversial, with conflicting reports published in 2018.[64]
The body contains a variety of stem cell types that have the capacity to differentiate into neurons. Researchers found a way to transform human skin cells into nerve cells using transdifferentiation, in which «cells are forced to adopt new identities».[65]
During neurogenesis in the mammalian brain, progenitor and stem cells progress from proliferative divisions to differentiative divisions. This progression leads to the neurons and glia that populate cortical layers. Epigenetic modifications play a key role in regulating gene expression in differentiating neural stem cells, and are critical for cell fate determination in the developing and adult mammalian brain. Epigenetic modifications include DNA cytosine methylation to form 5-methylcytosine and 5-methylcytosine demethylation.[66] These modifications are critical for cell fate determination in the developing and adult mammalian brain. DNA cytosine methylation is catalyzed by DNA methyltransferases (DNMTs). Methylcytosine demethylation is catalyzed in several stages by TET enzymes that carry out oxidative reactions (e.g. 5-methylcytosine to 5-hydroxymethylcytosine) and enzymes of the DNA base excision repair (BER) pathway.[66]
At different stages of mammalian nervous system development two DNA repair processes are employed in the repair of DNA double-strand breaks. These pathways are homologous recombinational repair used in proliferating neural precursor cells, and non-homologous end joining used mainly at later developmental stages[67]
Intercellular communication between developing neurons and microglia is also indispensable for proper neurogenesis and brain development.[68]
Nerve regeneration[edit]
Peripheral axons can regrow if they are severed,[69] but one neuron cannot be functionally replaced by one of another type (Llinás’ law).[18]
See also[edit]
- Artificial neuron
- Bidirectional cell
- Biological neuron model
- Compartmental neuron models
- Connectome
- Dogiel cell
- List of animals by number of neurons
- List of neuroscience databases
- Neuronal galvanotropism
- Neuroplasticity
- Growth cone
- Sholl analysis
References[edit]
- ^ >Moore, Keith; Dalley, Arthur (2005). Clinically Oriented Anatomy (5th ed.). LWW. pp. 47. ISBN 0-7817-3639-0.
A bundle of nerve fibers (axons) connecting neighboring or distant nuclei of the CNS is a tract.
- ^ «What are the parts of the nervous system?». Retrieved 2022-07-08.
- ^ Davies, Melissa (2002-04-09). «The Neuron: size comparison». Neuroscience: A journey through the brain. Retrieved 2009-06-20.
- ^ Chudler EH. «Brain Facts and Figures». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ «16.7: Nervous System». Biology LibreTexts. 2021-01-14. Retrieved 2022-02-28.
- ^ Herrup K, Yang Y (May 2007). «Cell cycle regulation in the postmitotic neuron: oxymoron or new biology?». Nature Reviews. Neuroscience. 8 (5): 368–78. doi:10.1038/nrn2124. PMID 17453017. S2CID 12908713.
- ^ Giménez, C. (February 1998). «[Composition and structure of the neuronal membrane: molecular basis of its physiology and pathology]». Revista de Neurologia. 26 (150): 232–239. ISSN 0210-0010. PMID 9563093.
- ^ State Hospitals Bulletin. State Commission in Lunacy. 1897. p. 378.
- ^ «Medical Definition of Neurotubules». www.merriam-webster.com.
- ^ Zecca L, Gallorini M, Schünemann V, Trautwein AX, Gerlach M, Riederer P, Vezzoni P, Tampellini D (March 2001). «Iron, neuromelanin and ferritin content in the substantia nigra of normal subjects at different ages: consequences for iron storage and neurodegenerative processes». Journal of Neurochemistry. 76 (6): 1766–73. doi:10.1046/j.1471-4159.2001.00186.x. PMID 11259494. S2CID 31301135.
- ^ Herrero MT, Hirsch EC, Kastner A, Luquin MR, Javoy-Agid F, Gonzalo LM, Obeso JA, Agid Y (1993). «Neuromelanin accumulation with age in catecholaminergic neurons from Macaca fascicularis brainstem». Developmental Neuroscience. 15 (1): 37–48. doi:10.1159/000111315. PMID 7505739.
- ^ Brunk UT, Terman A (September 2002). «Lipofuscin: mechanisms of age-related accumulation and influence on cell function». Free Radical Biology & Medicine. 33 (5): 611–9. doi:10.1016/s0891-5849(02)00959-0. PMID 12208347.
- ^ Zhao B, Meka DP, Scharrenberg R, König T, Schwanke B, Kobler O, Windhorst S, Kreutz MR, Mikhaylova M, Calderon de Anda F (August 2017). «Microtubules Modulate F-actin Dynamics during Neuronal Polarization». Scientific Reports. 7 (1): 9583. Bibcode:2017NatSR…7.9583Z. doi:10.1038/s41598-017-09832-8. PMC 5575062. PMID 28851982.
- ^ Lee WC, Huang H, Feng G, Sanes JR, Brown EN, So PT, Nedivi E (February 2006). «Dynamic remodeling of dendritic arbors in GABAergic interneurons of adult visual cortex». PLOS Biology. 4 (2): e29. doi:10.1371/journal.pbio.0040029. PMC 1318477. PMID 16366735.
- ^ Al, Martini, Frederic Et (2005). Anatomy and Physiology’ 2007 Ed.2007 Edition. Rex Bookstore, Inc. p. 288. ISBN 978-971-23-4807-5.
- ^ Gerber U (January 2003). «Metabotropic glutamate receptors in vertebrate retina». Documenta Ophthalmologica. Advances in Ophthalmology. 106 (1): 83–7. doi:10.1023/A:1022477203420. PMID 12675489. S2CID 22296630.
- ^ Wilson NR, Runyan CA, Wang FL, Sur M (August 2012). «Division and subtraction by distinct cortical inhibitory networks in vivo». Nature. 488 (7411): 343–8. Bibcode:2012Natur.488..343W. doi:10.1038/nature11347. hdl:1721.1/92709. PMC 3653570. PMID 22878717.
- ^ a b Llinás RR (2014-01-01). «Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective». Frontiers in Cellular Neuroscience. 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ Kolodin YO, Veselovskaia NN, Veselovsky NS, Fedulova SA. Ion conductances related to shaping the repetitive firing in rat retinal ganglion cells. Acta Physiologica Congress. Archived from the original on 2012-10-07. Retrieved 2009-06-20.
- ^ «Ionic conductances underlying excitability in tonically firing retinal ganglion cells of adult rat». Ykolodin.50webs.com. 2008-04-27. Retrieved 2013-02-16.
- ^ Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y (January 2019). «Histamine: neural circuits and new medications». Sleep. 42 (1). doi:10.1093/sleep/zsy183. PMC 6335869. PMID 30239935.
- ^ «Patch-seq technique helps depict the variation of neural cells in the brain». News-medical.net. 3 December 2020. Retrieved 26 August 2021.
{{cite web}}: CS1 maint: url-status (link) - ^ Macpherson, Gordon (2002). Black’s Medical Dictionary (40 ed.). Lanham, MD: Scarecrow Press. pp. 431–434. ISBN 0810849844.
- ^ Ivannikov MV, Macleod GT (June 2013). «Mitochondrial free Ca²⁺ levels and their effects on energy metabolism in Drosophila motor nerve terminals». Biophysical Journal. 104 (11): 2353–61. Bibcode:2013BpJ…104.2353I. doi:10.1016/j.bpj.2013.03.064. PMC 3672877. PMID 23746507.
- ^ Herculano-Houzel S (November 2009). «The human brain in numbers: a linearly scaled-up primate brain». Frontiers in Human Neuroscience. 3: 31. doi:10.3389/neuro.09.031.2009. PMC 2776484. PMID 19915731.
- ^ Drachman DA (June 2005). «Do we have brain to spare?». Neurology. 64 (12): 2004–5. doi:10.1212/01.WNL.0000166914.38327.BB. PMID 15985565. S2CID 38482114.
- ^ Ucar, Hasan; Watanabe, Satoshi; Noguchi, Jun; Morimoto, Yuichi; Iino, Yusuke; Yagishita, Sho; Takahashi, Noriko; Kasai, Haruo (December 2021). «Mechanical actions of dendritic-spine enlargement on presynaptic exocytosis». Nature. 600 (7890): 686–689. Bibcode:2021Natur.600..686U. doi:10.1038/s41586-021-04125-7. ISSN 1476-4687. PMID 34819666. S2CID 244648506.
Lay summary:
«Forceful synapses reveal mechanical interactions in the brain». Nature. 24 November 2021. doi:10.1038/d41586-021-03516-0. Retrieved 21 February 2022. - ^ «Researchers discover new type of cellular communication in the brain». The Scripps Research Institute. Retrieved 12 February 2022.
- ^ Schiapparelli, Lucio M.; Sharma, Pranav; He, Hai-Yan; Li, Jianli; Shah, Sahil H.; McClatchy, Daniel B.; Ma, Yuanhui; Liu, Han-Hsuan; Goldberg, Jeffrey L.; Yates, John R.; Cline, Hollis T. (25 January 2022). «Proteomic screen reveals diverse protein transport between connected neurons in the visual system». Cell Reports. 38 (4): 110287. doi:10.1016/j.celrep.2021.110287. ISSN 2211-1247. PMC 8906846. PMID 35081342.
- ^ Levitan, Irwin B.; Kaczmarek, Leonard K. (2015). «Electrical Signaling in Neurons». The Neuron. Oxford University Press. pp. 41–62. doi:10.1093/med/9780199773893.003.0003. ISBN 978-0-19-977389-3.
- ^ O’Leary, Olivia F.; Ogbonnaya, Ebere S.; Felice, Daniela; Levone, Brunno R.; C. Conroy, Lorraine; Fitzgerald, Patrick; Bravo, Javier A.; Forsythe, Paul; Bienenstock, John; Dinan, Timothy G.; Cryan, John F. (1 February 2018). «The vagus nerve modulates BDNF expression and neurogenesis in the hippocampus». European Neuropsychopharmacology. 28 (2): 307–316. doi:10.1016/j.euroneuro.2017.12.004. ISSN 0924-977X. PMID 29426666. S2CID 46819013.
- ^ Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z (January 2020). «Microglia monitor and protect neuronal function through specialized somatic purinergic junctions». Science. 367 (6477): 528–537. Bibcode:2020Sci…367..528C. doi:10.1126/science.aax6752. PMID 31831638. S2CID 209343260.
- ^ Chudler EH. «Milestones in Neuroscience Research». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ Patlak J, Gibbons R (2000-11-01). «Electrical Activity of Nerves». Action Potentials in Nerve Cells. Archived from the original on August 27, 2009. Retrieved 2009-06-20.
- ^ Harris-Warrick, RM (October 2011). «Neuromodulation and flexibility in Central Pattern Generator networks». Current Opinion in Neurobiology. 21 (5): 685–92. doi:10.1016/j.conb.2011.05.011. PMC 3171584. PMID 21646013.
- ^ Brown EN, Kass RE, Mitra PP (May 2004). «Multiple neural spike train data analysis: state-of-the-art and future challenges». Nature Neuroscience. 7 (5): 456–61. doi:10.1038/nn1228. PMID 15114358. S2CID 562815.
- ^ Thorpe SJ (1990). «Spike arrival times: A highly efficient coding scheme for neural networks» (PDF). In Eckmiller R, Hartmann G, Hauske G (eds.). Parallel processing in neural systems and computers. North-Holland. pp. 91–94. ISBN 9780444883902. Archived from the original (PDF) on 2012-02-15.
- ^ a b Kalat, James W (2016). Biological psychology (12 ed.). Australia. ISBN 9781305105409. OCLC 898154491.
- ^ Eckert R, Randall D (1983). Animal physiology: mechanisms and adaptations. San Francisco: W.H. Freeman. p. 239. ISBN 978-0-7167-1423-1.
- ^ a b Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
Ramon y Cajal’s first paper on the Golgi stain was on the bird cerebellum, and it appeared in the Revista in 1888. He acknowledged that he found the nerve fibers to be very intricate, but stated that he could find no evidence for either axons or dendrites undergoing anastomosis and forming nets. He called each nervous element ‘an absolutely autonomous canton.’
- ^ a b Oxford English Dictionary, 3rd edition, 2003, s.v.
- ^ Mehta AR, Mehta PR, Anderson SP, MacKinnon BL, Compston A (January 2020). «Grey Matter Etymology and the neuron(e)». Brain. 143 (1): 374–379. doi:10.1093/brain/awz367. PMC 6935745. PMID 31844876.
- ^ «Google Books Ngram Viewer». books.google.com. Retrieved 19 December 2020.
- ^ a b c d López-Muñoz F, Boya J, Alamo C (October 2006). «Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal». Brain Research Bulletin. 70 (4–6): 391–405. doi:10.1016/j.brainresbull.2006.07.010. PMID 17027775. S2CID 11273256.
- ^ Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
… a man who would write a highly influential review of the evidence in favor of the neuron doctrine two years later. In his paper, Waldeyer (1891), … , wrote that nerve cells terminate freely with end arborizations and that the ‘neuron’ is the anatomical and physiological unit of the nervous system. The word ‘neuron’ was born this way.
- ^ «Whonamedit — dictionary of medical eponyms». www.whonamedit.com.
Today, Wilhelm von Waldeyer-Hartz is remembered as the founder of the neurone theory, coining the term «neurone» to describe the cellular function unit of the nervous system and enunciating and clarifying that concept in 1891.
- ^ Grant G (October 2007). «How the 1906 Nobel Prize in Physiology or Medicine was shared between Golgi and Cajal». Brain Research Reviews. 55 (2): 490–8. doi:10.1016/j.brainresrev.2006.11.004. PMID 17306375. S2CID 24331507.
- ^ Witcher MR, Kirov SA, Harris KM (January 2007). «Plasticity of perisynaptic astroglia during synaptogenesis in the mature rat hippocampus». Glia. 55 (1): 13–23. CiteSeerX 10.1.1.598.7002. doi:10.1002/glia.20415. PMID 17001633. S2CID 10664003.
- ^ Connors BW, Long MA (2004). «Electrical synapses in the mammalian brain». Annual Review of Neuroscience. 27 (1): 393–418. doi:10.1146/annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Guillery RW (June 2005). «Observations of synaptic structures: origins of the neuron doctrine and its current status». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1458): 1281–307. doi:10.1098/rstb.2003.1459. PMC 1569502. PMID 16147523.
- ^ Sabbatini RM (April–July 2003). «Neurons and Synapses: The History of Its Discovery». Brain & Mind Magazine: 17.
- ^ Djurisic M, Antic S, Chen WR, Zecevic D (July 2004). «Voltage imaging from dendrites of mitral cells: EPSP attenuation and spike trigger zones». The Journal of Neuroscience. 24 (30): 6703–14. doi:10.1523/JNEUROSCI.0307-04.2004. hdl:1912/2958. PMC 6729725. PMID 15282273.
- ^ Cochilla AJ, Alford S (March 1997). «Glutamate receptor-mediated synaptic excitation in axons of the lamprey». The Journal of Physiology. 499 (Pt 2): 443–57. doi:10.1113/jphysiol.1997.sp021940. PMC 1159318. PMID 9080373.
- ^ Gouwens NW, Wilson RI (2009). «Signal propagation in Drosophila central neurons». Journal of Neuroscience. 29 (19): 6239–6249. doi:10.1523/jneurosci.0764-09.2009. PMC 2709801. PMID 19439602.
- ^ Williams RW, Herrup K (1988). «The control of neuron number». Annual Review of Neuroscience. 11 (1): 423–53. doi:10.1146/annurev.ne.11.030188.002231. PMID 3284447.
- ^ von Bartheld CS, Bahney J, Herculano-Houzel S (December 2016). «The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting». The Journal of Comparative Neurology. 524 (18): 3865–3895. doi:10.1002/cne.24040. PMC 5063692. PMID 27187682.
- ^ Krajewski KM, Lewis RA, Fuerst DR, Turansky C, Hinderer SR, Garbern J, Kamholz J, Shy ME (July 2000). «Neurological dysfunction and axonal degeneration in Charcot-Marie-Tooth disease type 1A». Brain. 123 (7): 1516–27. doi:10.1093/brain/123.7.1516. PMID 10869062.
- ^ «About Alzheimer’s Disease: Symptoms». National Institute on Aging. Archived from the original on 15 January 2012. Retrieved 28 December 2011.
- ^ Burns A, Iliffe S (February 2009). «Alzheimer’s disease». BMJ. 338: b158. doi:10.1136/bmj.b158. PMID 19196745. S2CID 8570146.
- ^ Querfurth HW, LaFerla FM (January 2010). «Alzheimer’s disease». The New England Journal of Medicine. 362 (4): 329–44. doi:10.1056/NEJMra0909142. PMID 20107219. S2CID 205115756.
- ^ «Parkinson’s Disease Information Page». NINDS. 30 June 2016. Archived from the original on 4 January 2017. Retrieved 18 July 2016.
- ^ «Movement Disorders». The International Neuromodulation Society.
- ^ Kerschensteiner M, Schwab ME, Lichtman JW, Misgeld T (May 2005). «In vivo imaging of axonal degeneration and regeneration in the injured spinal cord». Nature Medicine. 11 (5): 572–7. doi:10.1038/nm1229. PMID 15821747. S2CID 25287010.
- ^ Kempermann G, Gage FH, Aigner L, Song H, Curtis MA, Thuret S, Kuhn HG, Jessberger S, Frankland PW, Cameron HA, Gould E, Hen R, Abrous DN, Toni N, Schinder AF, Zhao X, Lucassen PJ, Frisén J (July 2018). «Human Adult Neurogenesis: Evidence and Remaining Questions». Cell Stem Cell. 23 (1): 25–30. doi:10.1016/j.stem.2018.04.004. PMC 6035081. PMID 29681514.
- ^ Callaway, Ewen (26 May 2011). «How to make a human neuron». Nature. doi:10.1038/news.2011.328.
By transforming cells from human skin into working nerve cells, researchers may have come up with a model for nervous-system diseases and perhaps even regenerative therapies based on cell transplants. The achievement, reported online today in Nature, is the latest in a fast-moving field called transdifferentiation, in which cells are forced to adopt new identities. In the past year, researchers have converted connective tissue cells found in skin into heart cells, blood cells, and liver cells.
- ^ a b Wang Z, Tang B, He Y, Jin P (March 2016). «DNA methylation dynamics in neurogenesis». Epigenomics. 8 (3): 401–14. doi:10.2217/epi.15.119. PMC 4864063. PMID 26950681.
- ^ Orii KE, Lee Y, Kondo N, McKinnon PJ (June 2006). «Selective utilization of nonhomologous end-joining and homologous recombination DNA repair pathways during nervous system development». Proceedings of the National Academy of Sciences of the United States of America. 103 (26): 10017–22. Bibcode:2006PNAS..10310017O. doi:10.1073/pnas.0602436103. PMC 1502498. PMID 16777961.
- ^ Cserép, Csaba; Schwarcz, Anett D.; Pósfai, Balázs; László, Zsófia I.; Kellermayer, Anna; Környei, Zsuzsanna; Kisfali, Máté; Nyerges, Miklós; Lele, Zsolt; Katona, István (September 2022). «Microglial control of neuronal development via somatic purinergic junctions». Cell Reports. 40 (12): 111369. doi:10.1016/j.celrep.2022.111369. PMC 9513806. PMID 36130488. S2CID 252416407.
- ^ Yiu G, He Z (August 2006). «Glial inhibition of CNS axon regeneration». Nature Reviews. Neuroscience. 7 (8): 617–27. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
Further reading[edit]
- Bullock TH, Bennett MV, Johnston D, Josephson R, Marder E, Fields RD (November 2005). «Neuroscience. The neuron doctrine, redux». Science. 310 (5749): 791–3. doi:10.1126/science.1114394. PMID 16272104. S2CID 170670241.
- Kandel ER, Schwartz JH, Jessell TM (2000). Principles of Neural Science (4th ed.). New York: McGraw-Hill. ISBN 0-8385-7701-6.
- Peters A, Palay SL, Webster HS (1991). The Fine Structure of the Nervous System (3rd ed.). New York: Oxford University Press. ISBN 0-19-506571-9.
- Ramón y Cajal S (1933). Histology (10th ed.). Baltimore: Wood.
- Roberts A, Bush BM (1981). Neurones without Impulses. Cambridge: Cambridge University Press. ISBN 0-521-29935-7.
- Snell RS (2010). Clinical Neuroanatomy. Lippincott Williams & Wilkins. ISBN 978-0-7817-9427-5.
External links[edit]
- Neurobiology at Curlie
- IBRO (International Brain Research Organization). Fostering neuroscience research especially in less well-funded countries.
- NeuronBank an online neuromics tool for cataloging neuronal types and synaptic connectivity.
- High Resolution Neuroanatomical Images of Primate and Non-Primate Brains.
- The Department of Neuroscience at Wikiversity, which presently offers two courses: Fundamentals of Neuroscience and Comparative Neuroscience.
- NIF Search – Neuron via the Neuroscience Information Framework
- Cell Centered Database – Neuron
- Complete list of neuron types according to the Petilla convention, at NeuroLex.
- NeuroMorpho.Org an online database of digital reconstructions of neuronal morphology.
- Immunohistochemistry Image Gallery: Neuron
- Khan Academy: Anatomy of a neuron
- Neuron images
Что такое дендрит — функции и морфология
Дендриты (dendrite) — многочисленные тонкие трубчатые или округлые выпячивания клеточного тела (перикариона) нервной клетки. Сам термин говорит о чрезвычайной разветвленности этих участков нейронов (от греч. δένδρον (dendron) — дерево).
В поверхностной структуре нейроцитов могут насчитываться от нуля до множества дендритов. Аксон чаще всего единственный. Поверхность дендритов не имеет миелиновой оболочки в отличие от аксонных отростков.

Цитоплазма содержит те же клеточные компоненты, что и само тело нервной клетки:
- эндоплазматический гранулярный ретикулум;
- скопления рибосом — полисомы (белоксинтезирующие органеллы);
- митохондрии (энергетические “станции” клетки, которые, используя глюкозу и кислород, синтезируют необходимые высокоэнергетические молекулы);
- аппарат Гольджи (отвечает за доставку внутренних секретов к внешнему слою клетки);
- нейротубулы (микротрубочки) и нейрофиламенты — главные компоненты цитоплазмы, тонкие опорные структуры, которые обеспечивают сохранение определенной формы.
Строение дендритных окончаний напрямую связано с их физиологическими функциями — получением информации от аксонов, дендритов, перикариона соседних нервных клеток посредством многочисленных межнейронных контактов на основе избирательной чувствительности к определенным сигналам.
Строение нейрона: аксоны и дендриты
Основные статьи: Дендрит,Аксон
Схема строения нейрона
Аксон— обычно длинный отросток нейрона, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу.Дендриты— как правило, короткие и сильно разветвлённые отростки нейрона, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновойоболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Основная статья: Синапс
Си́напс(греч.σύναψις, отσυνάπτειν— обнимать, обхватывать, пожимать руку) — место контакта между двумянейронамиили между нейроном и получающей сигналэффекторнойклеткой. Служит для передачинервного импульсамежду двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
- Общая информация
- Аксоны нейрона
- Дендриты нейрона
- Тело
Важнейший элемент в нервной системе – нейронная клетка, или простой нейрон. Это специфическая единица нервной ткани, задействованная в передаче и первичной обработке информации, а так же, являющаяся главным структурным образованием в ЦНС. Как правило, клетки имеют универсальные принципы строения и включают в себя помимо тела, еще аксоны нейронов и дендриты.
Структура и типы
Внешняя поверхность дендритов покрыта тонкими выпячиваниями в виде мельчайших шипиков размером 2-3 мкм. Количество таких формирований на поверхности может варьировать от нуля до десятка тысяч. Формы самих микрошипиков многообразны, но самой часто встречаемой формой считается грибовидный шипик.
Количество шипиков на поверхности и их размеры могут быстро меняться. От этого зависит реакция нейрона на сигналы от других клеток.
На образование выпячиваний-шипиков, их форму и развитие влияют внутренние и внешние обстоятельства: возраст организма, активность синаптических связей, информационная загруженность нейронных цепей, образ жизни организма и многое другое.
Целостность и стабильность структуры шипиков могут подвергаться влиянию негативных факторов:
- патофизиологические факторы (например, нейродегенеративные процессы в нервной ткани, опосредованные тяжелой наследственностью);
- токсикологические агенты (при употреблении наркотиков, алкоголя, ядов различной природы).
Под воздействием этих негативных факторов во внутреннем строении микрошипиков происходят серьезные деструктивные превращения: разрушение цистерн шипикового аппарата, накопление мультивезикулярных тел (пропорционально степени разрушительных влияний).
После серии испытаний, проведенной с подопытными мышами, было доказано, что не столько сами дендриты, сколько дендритные шипики являются элементарными единицами хранения памяти и формирования синаптической пластичности.

Ветвление
Дендритные структуры образуются вследствие древовидного разветвления отростков нейронов. Этот процесс называется арборизацией. Количество точек (или узлов) ветвления обуславливает степень разветвленности и сложность окончаний дендрита. В цитоплазме узлов ветвления обычно сконцентрированы митохондрии, так как ветвление – энергозатратный физиологический процесс.
Структура дендритного дерева обуславливает физическую восприимчивую площадь, то есть количество входных импульсов, которые суммарно сможет принять и провести нейроцит.
Одно из главных предназначений дендритов состоит в наращивании контактной поверхности для синапсов (увеличении рецепторного поля).
Это позволяет клетке принимать и перенаправлять больший объем информации, которая поступает к телу нейрона. Степень разветвленности определяет то, как нейрон в итоге суммирует электрические сигналы, полученные от других клеток: чем больше и сложнее ветвление, тем более плотно нейроны прилегают друг к другу.
За счет разветвленного строения поверхность рецепторной мембраны нервной клетки увеличивается в 1000 и более раз.

Диаметр и длина
Дендритные окончания имеют разные размеры, но всегда характеризуются постепенным уменьшением диаметра претерминальных веточек. Длина обычно от нескольких мкм до 1 мм. Но, например, у некоторых чувствительных нейронов спинномозговых ганглиев дендриты очень длинные – до метра и более.
Строение дендрита
На основании изучения микроскопических препаратов нервных клеток установили, что большинство отростков имеют цилиндрическую форму. Их диаметр в среднем составляет 0,9 мкм. Длина дендритов варьирует в широких пределах. Например, звездчатые нейроны серого вещества коры головного мозга имеют короткие (не более 200 мкм) ветви дендритного дерева, тогда как отростки двигательного нейрона, входящего в передние рога спинного мозга, составляют порядка 2 мм.
Специальные образования — шипики, формирующиеся на ветвях нейроцитов, приводят к появлению большого числа синапсов – щелевидных мест контакта с аксоном, дендритом или сомой другого нейрона. Синапсы могут располагаться на теле дендрита и называются стволовыми или же непосредственно на его шипиках. Как мы уже знаем, дендриты – это разветвленные отростки нейроцитов, способные принимать возбуждение. Передача же биопотенциалов происходит в них с помощью молекул химических соединений — медиаторов, например, ГАМК или ацетилхолина. В мембране, покрывающей дендрит, обнаружены ионные каналы, избирательно пропускающие катионы кальция, натрия и калия, участвующие в прохождении нервных импульсов через нейрон.

Проведение нервного импульса
Рецепторная мембрана поверхности дендритов (как и тела нервной клетки) покрыта многочисленными синаптическими бляшками, которые передают возбуждение на восприимчивый участок поверхностной мембраны нейрона, где генерируется биоэлектрический потенциал.
Информация, закодированная в виде электрических импульсов, передается на электровозбудимую проводящую мембрану аксона. Таким образом формируются нейронные сети организма.

На что влияет разветвленность нервных отростков
Тело нейрона является универсальным передающим и одновременно принимающим биологическим объектом. Объем (прежде всего поступающей информации) прямо пропорционален количеству входящих нервных импульсов. Они определяются по степени ветвления дендритного дерева. Поэтому дендриты – это структуры нейроцита, играющие интегративную функцию.

Более того, отростки расширяют площадь контакта нервных клеток между собой. Дополнительное же образование синапсов в разы повышает эффективность работы всех отделов, как головного и спинного мозга, так и нервной системы в целом.
Дендритная кристаллизация
В случае увеличения температуры в сторону жидкой фазы, то есть при положительном градиенте температуры , образуются относительно плоские грани кристаллов. Пока кристаллы окружены жидкой фазой, они растут свободно и имеют почти правильную геометрическую форму, которая определяется анизотропией скорости роста. В результате взаимного столкновения граней соседних кристаллов на завершающей стадии кристаллизации их правильная форма искажается. Так образуются кристаллиты произвольной формы.
В металлах и сплавах вследствие интенсивного выделения скрытой теплоты кристаллизации на границе кристалла и сильно переохлажденного расплава возникает отрицательный градиент температуры, который меняет механизм роста кристаллов. Чем дальше от границы, тем больше является переохлаждение расплава. Поэтому любая выпуклость на поверхности зародыша попадает в зону большего переохлаждения и растет вглубь расплава быстрее, чем плоские участки поверхности, образуя вытянутую первичную ось кристалла.
Направление роста осей кристалла совпадает с направлением оси пирамиды, которую образуют плоскости густого упаковки атомов. Поэтому в кристаллах с кубической или тетрагональной решеткой оси кристаллитов перпендикулярны между собой, а с гексагональной — размещены под углом 60 °. Наконец, когда температуры осей кристалла и расплава между ними выравниваются, начинается медленнее кристаллизация в межосевому пространстве. Так из зародышей вырастают древовидные кристаллиты — дендриты (с греч. Δένδρον — дерево). Механизм такого роста называют дендритным.
Дендриты растут до момента столкновения с соседями на завершающей стадии кристаллизации, приобретая при этом нерегулярной формы и определенных размеров. Размер дендритных кристаллитов является важным фактором, определяющим свойства литых сплавов. Мелкодисперсная структура обеспечивает лучшие механические и технологические свойства по сравнению с грубодисперсной. В практике литейного производства для большинства отливок стремятся обеспечить мелкозернистую структуру, реализующих преимущественно модифицирования или из-за изменения температуры расплава или скорости охлаждения при кристаллизации.
( 1 оценка, среднее 5 из 5 )
Нейрон состоит из
тела диаметром от 3 до 130 мкм, содержащего
ядро (с большим количеством ядерных
пор) и органеллы (в том числе сильно
развитый шероховатый ЭПР с активными
рибосомами, аппарат Гольджи), а также
из отростков. Выделяют два вида отростков:
дендриты и аксоны.
Аксон — обычно
длинный отросток, приспособленный для
проведения возбуждения от тела нейрона.
Дендриты — как правило, короткие и
сильно разветвлённые отростки, служащие
главным местом образования влияющих
на нейрон возбуждающих и тормозных
синапсов (разные нейроны имеют различное
соотношение длины аксона и дендритов).
Нейрон может иметь несколько дендритов
и обычно только один аксон. Один нейрон
может иметь связи со многими (до 20-и
тысяч) другими нейронами.
Дендриты делятся
дихотомически, аксоны же дают коллатерали.
В узлах ветвления обычно сосредоточены
митохондрии.
Дендриты не имеют
миелиновой оболочки, аксоны же могут
её иметь. Местом генерации возбуждения
у большинства нейронов является аксонный
холмик — образование в месте отхождения
аксона от тела. У всех нейронов эта зона
называется триггерной.
Вопрос № 3. Строение нейрона.
Тело нервной клетки
состоит из протоплазмы (цитоплазмы и
ядра), снаружи ограничена мембраной из
двойного слоя липидов (билипидный слой).
Липиды состоят из гидрофильных головок
и гидрофобных хвостов, расположены
гидрофобными хвостами друг к другу,
образуя гидрофобный слой, который
пропускает только жирорастворимые
вещества (напр. кислород и углекислый
газ). На мембране находятся белки: на
поверхности (в форме глобул), на которых
можно наблюдать наросты полисахаридов
(гликокаликс), благодаря которым клетка
воспринимает внешнее раздражение, и
интегральные белки, пронизывающие
мембрану насквозь, в которых находятся
ионные каналы.
Нейрон состоит из
тела диаметром от 3 до 130 мкм, содержащего
ядро (с большим количеством ядерных
пор) и органеллы (в том числе сильно
развитый шероховатый ЭПР с активными
рибосомами, аппарат Гольджи), а также
из отростков. Выделяют два вида отростков:
дендриты и аксоны. Нейрон имеет развитый
цитоскелет, проникающий в его отростки.
Цитоскелет поддерживает форму клетки,
его нити служат «рельсами» для транспорта
органелл и упакованных в мембранные
пузырьки веществ (например, нейромедиаторов).
В теле нейрона выявляется развитый
синтетический аппарат, гранулярная ЭПС
нейрона окрашивается базофильно и
известна под названием «тигроид».
Тигроид проникает в начальные отделы
дендритов, но располагается на заметном
расстоянии от начала аксона, что служит
гистологическим признаком аксона.
Различается
антероградный (от тела) и ретроградный
(к телу) аксонный транспорт.
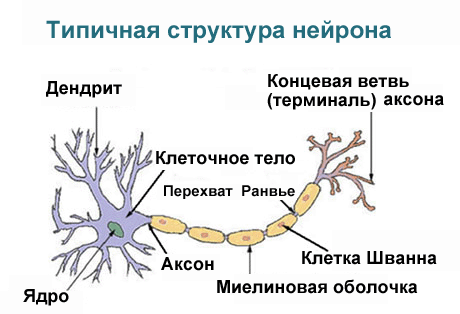
Вопрос № 4. Нейросекреторные клетки.
Ряд клеток обладающих
секреторной активностью, клетки
гипоталамуса. Они вырабатывают секрет
попадающий непосредственно в спинномозговую
жидкость. Имеют ряд особенностей-
липоиды. Гранулы секрета не растворяются
в воде и спирте. Ядра нейросекреторной
клетки не правильной формы, более
крупные, что говорит о высокой активности.
Секрет содержит белок, полисахариды и
липиды. В цитоплазме находятся гранулы
и капли секрета. Аксоны нейросекреторных
клеток характеризуются многочисленными
расширениями, которые возникают в связи
с временным накоплением нейросекрета.
Крупные и гигантские расширения
называются «телами Геринга». В пределах
мозга аксоны нейросекреторной клетки
лишены миелиновой оболочки.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Дендрит – это короткий, дихотомически (раздваивающийся) ветвящийся отросток, отходящий от тела нейрона. Согласно определению, отросток образует синапсы (электрические, химические) с другими элементами нервной ткани, через которые получает сигналы от периферических рецепторов и клеток организма в гистологической структуре органов и систем. Слово «дендрит» также имеет другое значение, если используется не в медицинской сфере, а в области материаловедения, в данном случае подразумевается кристаллическая структура материала независимо от его состава. Термин применим к камням, металлам, сплавам, искусственным соединениям, обладающим кристаллической, древовидной структурой.
Что такое дендрит — функции и морфология
Дендриты (dendrite) — многочисленные тонкие трубчатые или округлые выпячивания клеточного тела (перикариона) нервной клетки. Сам термин говорит о чрезвычайной разветвленности этих участков нейронов (от греч. δένδρον (dendron) — дерево).
В поверхностной структуре нейроцитов могут насчитываться от нуля до множества дендритов. Аксон чаще всего единственный. Поверхность дендритов не имеет миелиновой оболочки в отличие от аксонных отростков.
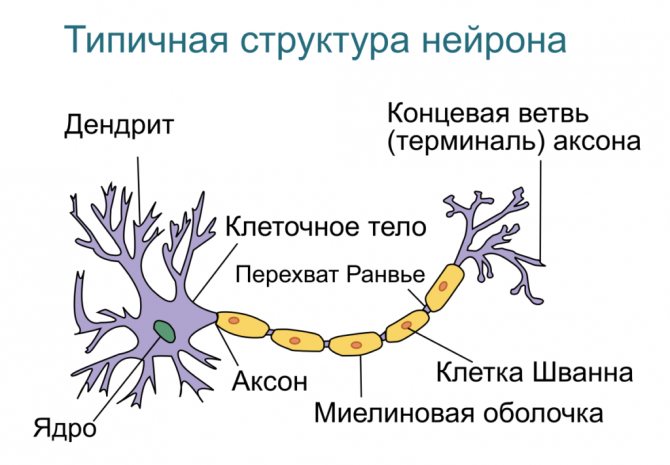
Цитоплазма содержит те же клеточные компоненты, что и само тело нервной клетки:
- эндоплазматический гранулярный ретикулум;
- скопления рибосом — полисомы (белоксинтезирующие органеллы);
- митохондрии (энергетические “станции” клетки, которые, используя глюкозу и кислород, синтезируют необходимые высокоэнергетические молекулы);
- аппарат Гольджи (отвечает за доставку внутренних секретов к внешнему слою клетки);
- нейротубулы (микротрубочки) и нейрофиламенты — главные компоненты цитоплазмы, тонкие опорные структуры, которые обеспечивают сохранение определенной формы.
Строение дендритных окончаний напрямую связано с их физиологическими функциями — получением информации от аксонов, дендритов, перикариона соседних нервных клеток посредством многочисленных межнейронных контактов на основе избирательной чувствительности к определенным сигналам.
Функции
Аксон и дендрит несмотря на разное морфологическое строение обладают схожими функциями – служат связующими элементами, благодаря которым поддерживается взаимодействие между всеми клетками организма, происходит интеграция всех физиологических процессов. Основная функция дендрита – восприятие сигналов от других нервных клеток и рецепторов внутренних органов. Дендритные ответвления также воспринимают внешние раздражители.
В результате образуются синаптические связи нескольких видов – аксонодендритические (контакт дендрит-аксон), дендро-дендритические (контакт дендрит-дендрит), аксошипиковые (контакт аксон-дендритный шипик). Полученные импульсы поступают к телу нейрона. Конечные сегменты дендритных отростков служат участком синаптического контакта, откуда к соме поступают тормозные и возбуждающие стимулы. Благодаря синаптическим контактам один нейрон связан с многочисленными (свыше 20 тысяч) нервными клетками.
Последние исследования показывают, что дендритные отростки способны самостоятельно генерировать сигналы. Ранее считалось, что импульсы генерирует только тело клетки, роль отростков сводится к передаче сигналов. Ученые выяснили, что активность дендритных отростков намного выше, чем сомы, когда изучали характер и силу сигналов, передающихся в пределах нервной ткани. Сигнал, проходящий по стволу отростка, может меняться.
Структура и типы
Внешняя поверхность дендритов покрыта тонкими выпячиваниями в виде мельчайших шипиков размером 2-3 мкм. Количество таких формирований на поверхности может варьировать от нуля до десятка тысяч. Формы самих микрошипиков многообразны, но самой часто встречаемой формой считается грибовидный шипик.
Количество шипиков на поверхности и их размеры могут быстро меняться. От этого зависит реакция нейрона на сигналы от других клеток.
На образование выпячиваний-шипиков, их форму и развитие влияют внутренние и внешние обстоятельства: возраст организма, активность синаптических связей, информационная загруженность нейронных цепей, образ жизни организма и многое другое.
Целостность и стабильность структуры шипиков могут подвергаться влиянию негативных факторов:
- патофизиологические факторы (например, нейродегенеративные процессы в нервной ткани, опосредованные тяжелой наследственностью);
- токсикологические агенты (при употреблении наркотиков, алкоголя, ядов различной природы).
Под воздействием этих негативных факторов во внутреннем строении микрошипиков происходят серьезные деструктивные превращения: разрушение цистерн шипикового аппарата, накопление мультивезикулярных тел (пропорционально степени разрушительных влияний).
После серии испытаний, проведенной с подопытными мышами, было доказано, что не столько сами дендриты, сколько дендритные шипики являются элементарными единицами хранения памяти и формирования синаптической пластичности.
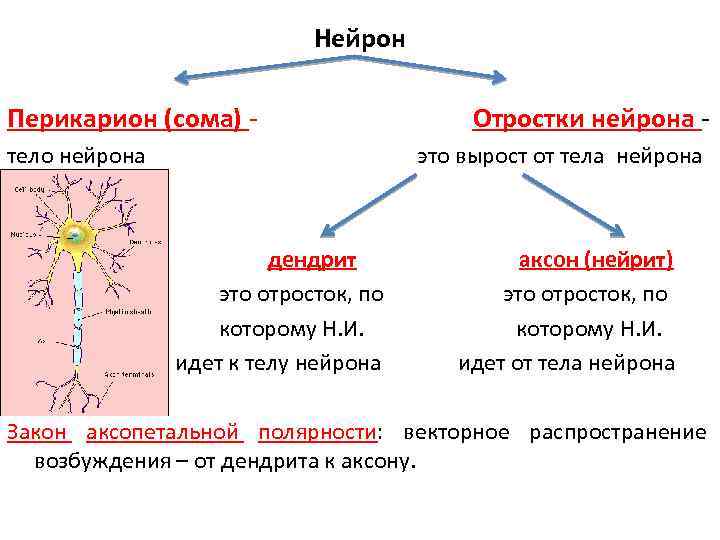
Ветвление
Дендритные структуры образуются вследствие древовидного разветвления отростков нейронов. Этот процесс называется арборизацией. Количество точек (или узлов) ветвления обуславливает степень разветвленности и сложность окончаний дендрита. В цитоплазме узлов ветвления обычно сконцентрированы митохондрии, так как ветвление – энергозатратный физиологический процесс.
Структура дендритного дерева обуславливает физическую восприимчивую площадь, то есть количество входных импульсов, которые суммарно сможет принять и провести нейроцит.
Одно из главных предназначений дендритов состоит в наращивании контактной поверхности для синапсов (увеличении рецепторного поля).
Это позволяет клетке принимать и перенаправлять больший объем информации, которая поступает к телу нейрона. Степень разветвленности определяет то, как нейрон в итоге суммирует электрические сигналы, полученные от других клеток: чем больше и сложнее ветвление, тем более плотно нейроны прилегают друг к другу.
За счет разветвленного строения поверхность рецепторной мембраны нервной клетки увеличивается в 1000 и более раз.
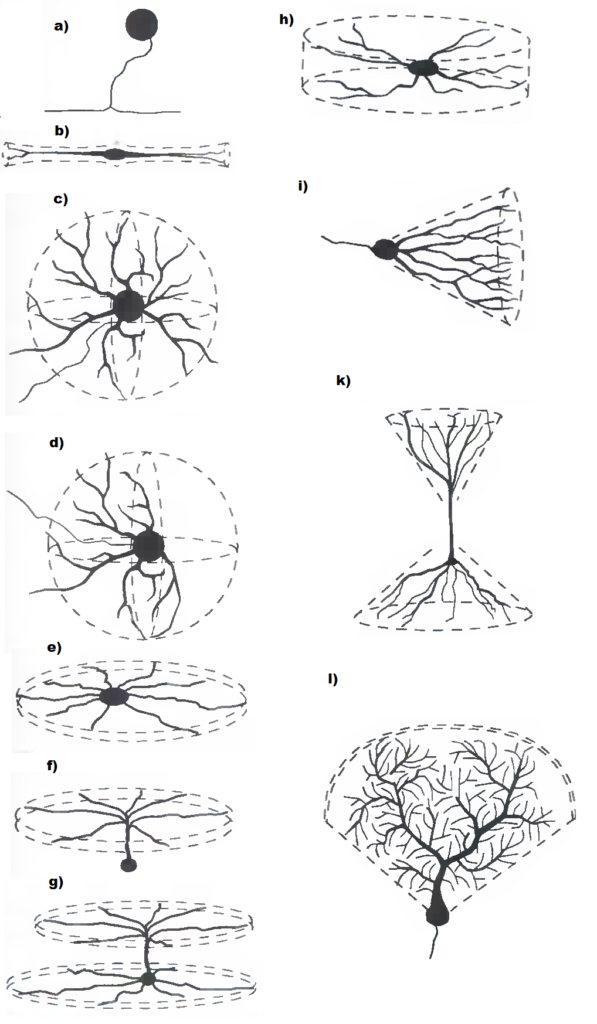
Диаметр и длина
Дендритные окончания имеют разные размеры, но всегда характеризуются постепенным уменьшением диаметра претерминальных веточек. Длина обычно от нескольких мкм до 1 мм. Но, например, у некоторых чувствительных нейронов спинномозговых ганглиев дендриты очень длинные – до метра и более.
Общие сведения
Нейрон – основная структурно-функциональная единица нервной ткани. Нервная клетка – образование с многочисленными отростками размером 4-130 мкм. От нейрона отходят несколько (реже один) дендритов и единственный аксон. Дендрит в биологии – это такой отросток, который передает возбуждение от периферических рецепторов к телу нейрона, что обуславливает его ведущую роль в восприятии внешних стимулов. Особенности дендритных ответвлений, которые наблюдаются в ходе микроскопических исследований:
- Система микротрубочек.
- Наличие шипиков, отходящих от общей оси.
- Присутствие узлов ветвления.
- Наличие эндоплазматического ретикулума (внутриклеточный органоид, представленный системой канальцев, полостей, пузырьков, окруженных мембраной).
Отростки, находящиеся рядом с сомой (телом), утолщенные, образуют большое число синаптических контактов. Мембрана отростка наподобие мембраны самого нейрона состоит из большого количества белковых молекул, которые играют роль химических рецепторов. Рецепторные образования наделены специфической чувствительностью к определенным химическим соединениям.
Обозначенные химические вещества – нейромедиаторы торможения и возбуждения, активно участвуют в процессе, когда импульсы распространяются по нервной ткани и поступают к соме. Строение дендрита предполагает наличие шипиков, которые образуют синаптические контакты с терминалями – концевыми участками сотен тысяч нервных клеток. Огромное количество шипиков располагается на отростках нервных клеток, образующих корковый слой больших полушарий.
Дендритный шипик сформирован из тела и головки. Размеры и форма структурных компонентов шипика существенно варьируются. Благодаря шипикам значительно увеличивается площадь постсинаптической мембраны. Шипики в дендритной структуре – лабильные образования, которые под воздействием внешних стимулов изменяют конфигурацию, дегенерируют (разрушаются), регенерируют (появляются вновь).
Количество синапсов определяет качество передачи импульсов и скорость, с которой они распространяются. Несколько дендритных ответвлений образуют единую ветку. Совокупность всех дендритов является дендритным деревом – поверхностью, воспринимающей сторонние раздражители. Исследования показывают, дендритные деревья составляют 90% мозгового вещества.
Проведение нервного импульса
Рецепторная мембрана поверхности дендритов (как и тела нервной клетки) покрыта многочисленными синаптическими бляшками, которые передают возбуждение на восприимчивый участок поверхностной мембраны нейрона, где генерируется биоэлектрический потенциал.
Информация, закодированная в виде электрических импульсов, передается на электровозбудимую проводящую мембрану аксона. Таким образом формируются нейронные сети организма.
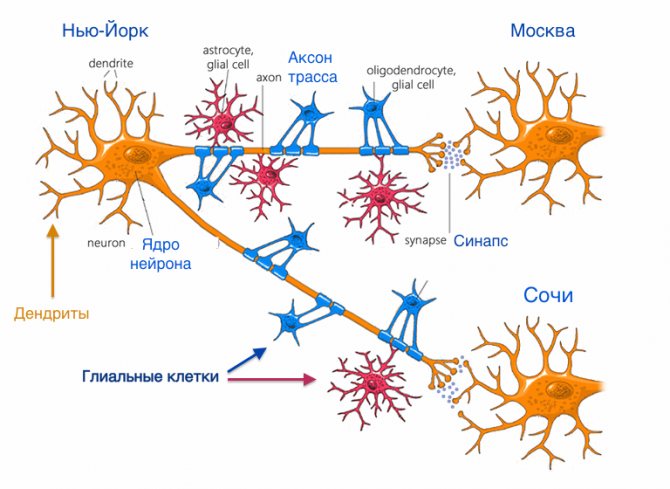
Роль в нейронных процессах
Человек рождается с генетически определенным числом отростков-дендритов на каждом нейроне. Постепенное увеличение и усложнение мозговых структур и построение нервной системы, которые происходят при постнатальном развитии, реализуется за счет разветвления, увеличения массы дендритов.
Согласно данным многочисленных исследований, в пике развития нервной системы дендриты занимают порядка 60-75 % от всей массы нервных клеток.
Согласно фундаментальным теориям, описывающим принципы работы нервной системы, дендриты всегда считались отделом нейрона, принимающим импульс и проводящим его на тело нервной клетки.
Однако современные исследования нейробиологов с использованием новейших технологий таких, как микроэлектроды, выявило большую электрическую активность дендритов по сравнению с телом клетки.
Данные исследования подтвердили тот факт, что дендритные окончания способны сами генерировать электрические импульсы – локальные потенциалы действия.
Строение и функции дендритов
.
Функции дендритов.
Хотелось бы отметить, что основные трудности, с которыми сталкивается исследователь при изучении функции дендритов, — это отсутствие сведений о свойствах мембраны дендрита (в отличие от мембраны тела нейрона) из-за невозможности введения микроэлектрода внутрь дендрита.
Оценивая в целом геометрию дендритов, распределение синапсов и особое строение цитоплазмы в местах дендритных ветвлений, можно говорить о специальных локусах нейрона со своей собственной функцией. Самое простое, что можно было бы приписать дендритным площадкам в местах ветвления — это трофическая функция.
Из всего вышесказанного следует, что цитоплазма дендритов содержит много ультраструктурных компонентов , способных обеспечивать их важные функции. В дендрите есть определенные локусы, где его работа имеет свои особенности.
Главное назначение многочисленных дендритных разветвлений нервной клетки — это обеспечить взаимосвязь с другими нейронами. В коре головного мозга млекопитающих большая доля аксодендрических связей приходится на контакты с особыми специализированными выростами дендритов – дендрическими шипиками. Дендритические шипики являются филогенетически самыми молодыми образованиями в нервной системе. В онтогенезе они созревают значительно позже других нервных структур и представляют собой наиболее пластичный аппарат нервной клетки.
Как правило, дендрический шипик имеет в коре мозга млекопитающих характерную форму. (рис. 2). От основного дендритного ствола отходит сравнительно узкая ножка, которая заканчивается расширением — головкой. Вероятно такая форма дендритического придатка (наличие головки) связана, с одной стороны с увеличением площади синаптического контакта с аксонным окончанием, с другой служит для размещения внутри шипика специализированных органел, в частности шипикового аппарата, который имеется только в дендритических шипиках коры мозга млекопитающих. В этой связи кажется уместной аналогия с формой синаптического аксонного окончания, когда
тонкое претерминальное волокно образует расширение. Это расширение (синаптическая бляшка) образует обширный контакт с иннервируемым субстратом и содержит внутри большой набор ультраструктурных компонентов (синаптические пузырьки, митохондрии, нейрофиламенты, гранулы гликогена).
Существует гипотеза (которую, в частности, разделяет и развивает нобелевский лаурят Ф. Крик) о том, что геометрия шипиков может меняться в зависимости от функционального состояния мозга. При этом узкая шейка шипика может расширяться, а сам шипик уплощается, в результате чего увеличивается эффективность аксо-шипикового контакта.
Если форма и размеры дендрических шипиков в коре мозга млекопитающих могут несколько варьировать, то наиболее постоянно в них наличие специфического шипикового аппарата. Он представляет собой комплекс взаимосвязанных канальцев (цистерн), расположенных, как правило, в головке шипика. Вероятно, это органелла связана с очень важными функциями, присущими филогенетически самым молодым мозговым образованиям, так как шипиковый аппарат встречается в основном в коре головного мозга, и только у высших животных .
Несмотря ни на что шипик является производным дендрита, в нем отсутствуют нейрофиламенты и дендритические трубочки, его цитоплазма содержит грубо или тонко гранулированный матрикс . Еще одной характерной чертой шипика в коре мозга является обязательное присутствие на них синаптических контактов с аксонными окончаниями. Цитоплазма шипика имеет специальные компоненты, которые отличают его от дендритных стволов. Можно отметить своеобразную триаду в цитоплазме шипика: субсинаптическая специализация активных зон — шипиковый аппарат — митохондрии. Учитывая многообразие сложных и важных функций, которые выполняют митохондрии, можно ожидать также сложных функциональных проявлений в «триадах» при синаптической передаче. Можно говорить о том, что цитоплазма дендритического шипика и шипиковый аппарат могут иметь непосредственное отношение к синаптической функции.
Дендритические шипики и концы дендритов также очень чувствительны к действию экстремальных факторов. При любом виде отравления (например, алкогольном, гипоксическом, тяжелыми металлами — свинцом, ртутью и т.д.) меняется количество выявленных шипиков на дендритах клеток коры больших полушарий. По всей вероятности, шипики при этом не исчезают, но у них нарушаются цитоплазматические компоненты, и они хуже импрегнируются солями тяжелых металлов. Так как шипики — один из структурных компонентов обеспечения межнейрональных контактов, то неполадки в них приводят к серьезным нарушениям функции мозга.
В некоторых случаях при кратковременном действии экстремального фактора может наступить на первый взгляд пара дорсальная ситуация, когда количество выявленных шипиков на дендритах клеток мозга не уменьшается, а увеличивается . Так, это наблюдается при экспериментальной ишемии мозга в начальный ее период. Параллельно с увеличением числа выявленных шипиков может улучшаться функциональное состояние мозга. В данном случае гипоксия является фактором, который способствует усилению метаболизма в нервной ткани, лучшей реализации резервов, не используемых в обычной обстановке, и быстрому сгоранию шлаков, накопленных в организме. Ультраструктурно это проявляется в более интенсивной проработке цитоплазмы шипиков, разрастании и увеличении цистерн шипикового аппарата. Вероятно, этот феномен положительного действия гипоксии наблюдается тогда, когда человек, испытывая большие физические нагрузки в условиях гипоксии, покоряет горные вершины. Эти трудности компенсируются затем более интенсивной продуктивной работой, как мозга, так и других органов.
Формирование дендритов.
Дендриты и их межнейрональные связи формируются в процессе онтогенетического развития мозга. Причем дендриты, в частности апикальных, у молодых особей какое-то время остаются свободными для образования новых контактов. Участки дендрита, расположенные ближе к телу клетки, возможно , связаны с более прочными и простыми — натуральными условными рефлексами, а концы оставлены для образования новых связей, ассоциаций.
В зрелом возрасте на дендритах уже нет свободных от межнейрональных контактов участков, зато при старении прежде страдают именно концы дендритов и по насыщенности контактами
у старых особей они напоминают дендриты детского возраста. Это происходит как из-за того, что ослабляются транспортные белоксинтезирующие процессы в клетке, так и из-за нарушений кровоснабжения мозга. Может быть, именно здесь кроется та морфологическая основа для такого широко известного в неврологии и в обыденной жизни факта, когда старики трудно осваивают что-то новое, часто забывают текущие события и очень хорошо помнят о прошлом. То же самое наблюдается и при отравлению.
Как уже отмечалось, увеличение и усложнение дендритного древа в филогенезе необходимы не только для восприятия большого числа приходящих импульсов, но и для предварительной обработки.
Дендриты нейронов центральной нервной системы обладают синаптической функцией на всем протяжении, причем концевые участки нисколько не уступают в этом срединным. Если же речь идет о дистальных (концевых) участках апикальных дендритов пирамидных нейронов коры больших полушарий, то их доля в осуществлении межнейрональных взаимодействий еще более значительна, чем проксимальных. Там к большему числу концевых синаптических бляшек на самом стволе и разветвлениях апикального дендрита присоединяются еще контакты на дендритических шипиках.
Изучая эту проблему при помощи электронной микроскопии, исследователи также убедились в том, что концевые участки дендритов плотно покрыты синаптическими бляшками и, таким образом, принимают непосредственное участие в межнейрональных взаимодействиях. (рис. 3) Электронная микроскопия также показала, что дендриты могут образовывать контакты между собой. Эти контакты могут быть или параллельными, которым большинство авторов приписывают электротонические свойства, или типичными ассиметричными синапсами с ясно выраженными органеллами, обеспечивающими химическую передачу. Такие дендро-дендритические контакты только еще начинают привлекать внимание исследователей. Итак, дендрит на всем своем протяжении выполняет синаптическую функцию. Каким же образом поверхность дендрита приспособлена для обеспечения контактов с аксонными окончаниями?
Поверхностная мембрана дендрита устроена так, чтобы максимальным образом использоваться для межнейрональных контактов. Дендрит весь изрыт углублениями, складками, карманами, имеет различного неровности рода микровыросты, шипы, грибовидные придатки и др. Все эти рельефы дендритных стволов соответствуют форме и размерам приходящих синаптических окончаний. Причем в различных отделах нервной системы и у разных животных рельеф дендритной поверхности имеет специфические особенности. Конечно же, самым замечательным выростом дендритической мембраны является дендритический шипик.
Дендриты очень чувствительны к действию различных экстремальных факторов. Нарушения в них приводят ко многим заболеваниям, например психическим расстройствам.
Заключение.
Если учесть те ультраструктурные особенности цитоплазмы узлов дендритических ветвлений, дендритических шипиков, о которых я рассказала выше, а также эволюционные тенденции к увеличению и усложнению дендритного древа, то становится понятным, какая поистине колоссальная и важная работа выполняется этими нервными структурами. часто в некоторой автономности от ядросодержащей области нейрона . Не всегда, вероятно, тело нервной клетки играет решающую роль в обработке нервных импульсов и генерации аксонных ответов. Чувствительные нейроны межпозвоночных узлов, где импульс генерируется в рецепторах у первого перехвата Ранвье, нейропиль ганглиев беспозвоночных, где аксоны отходят вдали от клеточных тел, вынесенных на периферию ганглия, — все это говорит о возможности обработки и ответной генерации импульсов во внеядерной зоне нейрона.
Такие разделы неврологии, как высшая нервная деятельность, психиатрия, нейрофармакалогия, проблемы памяти и научения, настоятельно требуют точных сведений о структуре межнейрональных связей, в которых дендритам принадлежит первостепенная роль. Сейчас очевидна необходимость многопланового целенаправленного подхода к изучению геометрии, ультраструктуры, цитохимии и молекулярной
организации дендритов и аксодендрических синапсов. Функции
дендритов настолько сложны и многогранны, что, все еще остаются окончательно не выясненными.
СПИСОК ЛИТЕРАТУРЫ:
- Н.С. Косицын: Нервная клетка — здоровая и больная; издательство «Знание»; 1987/89
- А.Г. Хрипкова: Возрастная психология; М; Просвещение; 1987
- Под редакцией Р.Сопера: Биология; издательство «Мир»; 1990
- Под редакцией А.В.Коробкова: Нормальная физиология; М; высшая школа; 1980