Нервная система выполняет ряд важных функций:
- обеспечивает связь организма с окружающим миром;
- управляет работой всех органов;
- координирует функционирование всех систем органов, обеспечивая их согласованную работу.
Нервная ткань
Нервная ткань отличается от других тканей нашего организма тем, что обладает особыми свойствами — возбудимостью и проводимостью. Эти свойства нервной ткани обусловлены особенностями её строения.
В состав нервной ткани входят клетки двух видов. Основные функции выполняют нейроны, а клетки-спутники (клетки нейроглии) служат опорой и обеспечивают обмен веществ.

Рис. (1). Нервная ткань
Функции нейронов: генерирование и передача нервных импульсов; обработка и хранение поступающей информации.
Нервный импульс — это волна возбуждения (биоэлектрическая волна), распространяющаяся по нервным клеткам.
Нейрон — основная клетка нервной ткани. Он имеет тело и отростки двух типов. В теле нейрона располагается ядро и органоиды, а по отросткам передаются нервные импульсы.
Дендриты — это отростки, по которым нервные импульсы передаются к телу нейрона. Эти отростки сильно ветвятся. У нейрона может быть несколько дендритов.
Аксон — это отросток, по которому импульсы передаются от тела клетки. Аксон обычно ветвится только на конце. У каждого нейрона всего один аксон.

Рис. (2). Строение нейрона
Аксоны часто окружены оболочкой из жироподобного вещества миелина. Это вещество имеет белый цвет. Скопления миелинизированных аксонов образуют белое вещество головного и спинного мозга. Тела нервных клеток и дендриты не покрыты миелином. Они серого цвета, а их группы составляют серое вещество центральной нервной системы.
Передача нервных импульсов с одной клетки на другую происходит в синапсах.
Синапс — это место контакта между двумя нейронами или между нейроном и клеткой рабочего органа.
Главными элементами синапса являются мембраны двух клеток (пресинаптическая и постсинаптическая мембраны) и пространство между ними (синаптическая щель).

Рис. (3). Строение синапса
В аксоне пресинаптического нейрона вырабатывается медиатор — особое вещество, с помощью которого происходит передача нервного импульса.
Под действием нервного импульса медиатор выделяется в синаптическую щель. Рецепторы постсинаптической мембраны реагируют на его появление и генерируют возникновение нервного импульса в следующем нейроне. Так в синапсе происходит химическая передача возбуждения с одной клетки на другую.
Нейроны различаются по своему строению и выполняемым функциям.

Рис. (4). Виды нейронов
По выполняемым функциям выделяют три типа нейронов.
Чувствительные (сенсорные) нейроны проводят информацию от органов в мозг. Тела таких нейронов находятся в нервных узлах вне центральной нервной системы.
Другая группа нейронов передаёт информацию от головного и спинного мозга к органам. Это двигательные (моторные) нейроны. Их тела находятся в сером веществе центральной нервной системы, а аксоны находятся за пределами ЦНС.
Третий вид нейронов осуществляет связь между чувствительными и двигательными нейронами. Это вставочные нейроны, они находятся в головном и спинном мозге.
Скопление нейронов в головном или спинном мозге называют ядром.

Рис. (5). Типы нейронов и синапсы
Связь между органами и центральной нервной системой осуществляется через нервы.
Нерв — это орган, в состав которого входят пучки нервных волокон, покрытые соединительнотканной оболочкой.

Рис. (6). Нерв
Нервы выполняют проводниковую функцию. Они связывают головной и спинной мозг с кожей, органами чувств и с внутренними органами.
Нервы бывают чувствительные, двигательные и смешанные.
Чувствительные нервы проводят нервные импульсы от рецепторов в мозг. В их состав входят дендриты чувствительных нейронов.
Двигательные нервы состоят из аксонов двигательных нейронов. Их функция — проведение импульсов от мозга к рабочим органам.
Смешанные нервы образованы чувствительными и двигательными волокнами и способные проводить импульсы как к ЦНС, так и от ЦНС.
Нервные сплетения представлены сетчатыми скоплениями нервных волокон разных нервов, связывающих ЦНС с внутренними органами, скелетными мышцами и кожей.
Наиболее известное солнечное сплетение находится в брюшной полости.
Источники:
Рис. 1. Нервная ткань https://image.shutterstock.com/image-photo/mammalian-nervous-tissue-under-microscope-600w-74170234.jpg
Рис. 2. Строение нейрона https://image.shutterstock.com/image-vector/education-chart-biology-nerve-cell-600w-661087429.jpg
Рис. 3. Строение синапса https://image.shutterstock.com/image-illustration/gap-between-two-nerve-cells-600w-1284912691.jpg
Рис. 4. Виды нейронов https://image.shutterstock.com/image-illustration/different-kinds-neurons-scheme-structure-600w-138356969.jpg
Рис. 5. Типы нейронов и синапсы © ЯКласс
Рис. 6. Нерв https://image.shutterstock.com/image-illustration/nerve-structure-anatomy-600w-1041115012.jpg
 Карта потока аксонного рефлекса. Стимуляция аксона может вызвать электрический ток ко всем эффекторным тканям, которые иннервирует нейрон, а также обратно к соме нейрона; это отличается от нормального нейрона, который запускается только по аксону.
Карта потока аксонного рефлекса. Стимуляция аксона может вызвать электрический ток ко всем эффекторным тканям, которые иннервирует нейрон, а также обратно к соме нейрона; это отличается от нормального нейрона, который запускается только по аксону.
аксонный рефлекс (или ответная реакция) — это ответ, стимулируемый периферическими нервами тела, который движется от тела нервной клетки и ветви для стимуляции органов-мишеней. Рефлексы — это отдельные реакции, которые реагируют на раздражитель, составляющий строительные блоки общей передачи сигналов в нервной системе организма. Нейроны — это возбудимые клетки, которые обрабатывают и передают эти рефлекторные сигналы через свои аксоны, дендриты и тела клеток. Аксоны непосредственно облегчают межклеточную коммуникацию, проецирующуюся из тела нейрональной клетки в другие нейроны, локальную мышечную ткань, железы и артериолы. При аксонном рефлексе передача сигналов начинается в середине аксона в месте стимуляции и передает сигналы непосредственно в эффекторный орган, минуя как центр интеграции, так и химический синапс, присутствующий в рефлексе спинного мозга. Импульс ограничен одним раздвоенным аксоном или нейроном, аксон которого разветвляется на два отдела и не вызывает общего ответа на окружающие ткани.
Рефлекторная дуга аксона отличается от рефлекторной дуги спинного мозга . В рефлекторном пути спинного мозга афферентный нейрон передает информацию спинному мозгу интернейронам. Эти интернейроны действуют коллективно, обрабатывают и воспринимают входящие стимулы и стимулируют эффекторные нейроны, действующие как центр интеграции. Эффекторные нейроны, покидающие центр интеграции, передают ответ исходной ткани, в которой возник рефлекс, что приводит к ответу. Рефлекс аксона приводит к локализованному ответу только на локально иннервируемые клетки того единственного нейрона, от которого возник сигнал. Путь рефлекса аксонов не включает в себя центр интеграции или синапс, который передает связь между нейронами в рефлекс спинного мозга. Таким образом, стимул направляется к эффекторному органу, не попадая в тело нейрональной клетки, и, следовательно, указывает на то, что рефлекс аксона не является истинным рефлексом, когда афферентные импульсы проходят через центральную нервную систему до того, как стимулируют эфферентные нейроны.
Аксонный рефлекс был обнаружен и был описан как «новый тип периферического рефлекса», который обходит центр интеграции и синапс в центральной нервной системе. Открытие аксонального рефлекса показало, что аксонный рефлекс активирует местные артериолы, вызывая расширение сосудов и сокращение мышц. Это сокращение мышц наблюдалось у людей, страдающих астмой, когда высвободившиеся нейропептиды вызывали сокращение гладких мышц дыхательных путей. Точно так же высвобождение холинергических агентов в судомоторных нервных окончаниях вызывает рефлекс аксона, который стимулирует потовые железы, заставляя тело потеть в ответ на тепло. Рефлекс аксонов возможен благодаря передаче сигналов от кожных рецепторов на коже.
Содержание
- 1 Исследования и открытия
- 2 Физиология
- 2.1 Расширение сосудов
- 2.2 Астма
- 2.3 Реакция на пот
- 3 Механизмы
- 4 См. Также
- 5 Ссылки
Исследования и открытия
Аксонный рефлекс был открыт двумя русскими учеными Ковалевским и Соковниным в 1873 году. Они описали аксонный рефлекс как новый тип периферического (или местного) рефлекса, при котором электрический сигнал начинается в середине аксон и передача немедленно пропускают как центр интеграции, так и химический синапс, что обычно наблюдается в рефлексе спинного мозга.
В 1890 году британский физиолог Джон Ньюпорт Лэнгли исследовал движение шерсти у кошек, подвергшихся воздействию низких температур. Лэнгли заметил, что даже после стимуляции кошачья шерсть на прилегающих участках продолжала расти. Лэнгли пришел к выводу, что первичная нейрональная стимуляция не прекращается после первого синапса, а скорее участвует в разветвлении соединений с множеством нейронов, вызывая рост кошачьей шерсти в окружающих областях. Лэнгли определил этот путь как «рефлекс аксонов».
 Нормальная рефлекторная дуга спинного мозга, тогда как рефлекс аксона будет обходить интернейрон. Это различие важно, потому что ранним исследователям приходилось различать рефлекс спинного мозга и рефлекс аксона, чтобы понять реакцию организма на стимулы.
Нормальная рефлекторная дуга спинного мозга, тогда как рефлекс аксона будет обходить интернейрон. Это различие важно, потому что ранним исследователям приходилось различать рефлекс спинного мозга и рефлекс аксона, чтобы понять реакцию организма на стимулы.
В начале 20 века британский кардиолог сэр Томас Льюис исследовал механическое истирание кожи. Кожа продемонстрировала трехфазный ответ. Сначала появляется красное пятно, которое распространяется наружу из-за высвобождения гистамина из тучных клеток. Во-вторых, более яркий красный цвет распространяется вокруг исходного пятна из-за расширения артериол. Последней фазой было выделение жидкости, заполненной волдырями, над исходным местом. Льюис полагал, что реакция кожи была вызвана расширением соседних кровеносных сосудов, которые были вызваны нервной системой через рефлекс аксонов. Этот трехфазный ответ был назван тройным ответом Льюиса. Расширение артериол в пораженной области происходит из-за расширения сосудов. Хотя Льюис наблюдал вазодилатацию, которая могла быть объяснена рефлексом аксона, пока не было прямых доказательств, объясняющих разветвление нервов от центра аксона, а не тела клетки, или какие химические агенты были ответственны за гусиную шишку, красную линию и расширение симптомы со стороны кровеносных сосудов.
В 1960-х годах ученые А. Янско-Габор и Я. Сольчани продемонстрировали, что при нанесении на кожу раздражающих химикатов и электростимуляторов стимулируются кожные ноцирецепторы. Эти датчики боли посылают сигналы в соседние ткани, что приводит к экстравазации, также известной как утечка из кровеносных сосудов. Этот ответ аналогичен исследованию Льюиса с расширением сосудов, поскольку оба они полагаются на неповрежденные сенсорные нервы, которые воздействуют на соседние ткани.
В конце 20 века возникли более сложные методы прямого наблюдения аксонного рефлекса из-за большего точные инструменты визуализации и более продвинутые методы. Одним из примеров являются исследования с использованием лазерного допплера, в которых используется лазерная допплеровская визуализация для наблюдения за кровотоком в коже и определения функции сосудов. Подобные методы экспериментального сбора позволяют получить экспериментальные данные, которые предлагают механизм, объясняющий, как взаимодействие нейронных факторов и генетических способностей делает некоторых людей более устойчивыми к холоду. Эти методы исследования помогли улучшить лечение и профилактику повреждений кожи и обморожений, вызванных простудой.
Физиология
Когда проксимальный импульс стимулирует рецепторы растяжения и тепла на одной ветви раздвоенного аксона, создаваемый сигнал перемещается назад к точке бифуркации аксона. Затем импульс отражается вниз по другой ветви аксона к исполнительному органу, вызывая рефлекс аксона. Аксональные рефлексы стимулируют многочисленные эффекторные органы, включая эндокринную, сосудистую и кровеносную системы, в зависимости от места стимуляции. Одним из примеров является зуд, тип ноцицепции, при котором рефлекс часто вызывает желание почесать. Соединение капсаицин может использоваться для истощения химических веществ в нервных окончаниях рефлекса аксонов и уменьшения симптомов зуда и боли.
Физиологически рефлекс аксона помогает поддерживать гомеостаз, или регулирование внутренней среды организма в ответ на изменение внешней среды, обеспечивая стабильность и относительно постоянство внутренней среды. Рефлекс аксона реагирует на внешние изменения температуры, химической концентрации и состава воздуха. Примеры механизмов, опосредованных рефлексом аксонов, включают зуд, воспаление, боль, астму и кожное кровообращение.
Расширение сосудов
Организм реагирует на различные типы травм, включая инфекцию, физическую травму или токсическое повреждение ткани. через воспаление. Когда болевые ощущения усиливаются, аксонный рефлекс стимулирует (и отвечает за) высвобождение многих необходимых химических веществ, которые способствуют локальному воспалению тканей в травмированной области. Аксонный рефлекс регулирует вазодилатацию или дополнительный кровоток к тканям-мишеням. Аксонный рефлекс позволяет мышцам сокращаться в кратчайшие сроки, регулируя прохождение сигнала в нервно-мышечном соединении.
 Сужение сосудов и расширение сосудов, эффект, который может быть вызван стимуляцией рефлекса аксонов в определенных тканях, продемонстрирован по сравнению с нормальным кровеносным сосудом.
Сужение сосудов и расширение сосудов, эффект, который может быть вызван стимуляцией рефлекса аксонов в определенных тканях, продемонстрирован по сравнению с нормальным кровеносным сосудом.
В кожном кровообращении рефлекс аксонов контролирует температуру и кровообращение в тканях посредством расширения сосудов. Маленькие нервные волокна, называемые терморецепторами, чувствительны к температуре и могут действовать как сенсоры, которые инициируют опосредованную аксонным рефлексом вазодилатацию. Нервно-мышечные заболевания можно предсказать на ранней стадии по наличию аномальных рефлексов мышечных волокон и соответствующих подергиваний. Это происходит из-за того, что аксоны могут генерировать свои собственные потенциалы действия, когда они сверхвозбуждены исходным стимулом; это известно как потенциал фасцикуляции в мышечном волокне. Фасцикуляции являются характерными чертами бокового амиотрофического склероза (БАС) и могут быть свидетельством аномального рефлекса аксона при дальнейших исследованиях.
.
Астма
В астме аксон рефлекс вызывает высвобождение различных нейропептидов, включая вещество P, нейрокинин A и кальцитонин. Все три нейропептида вызывают сокращение гладкой мускулатуры дыхательных путей, что также происходит по схожему механизму при аллергии.
Этот же механизм реакции также ответственен за потерю тепла телом в конечностях, что продемонстрировано с помощью теста Охотника. Одним из клинических тестов, которые могут быть выполнены для пациента, является QSART или количественный судомоторный аксонный рефлекс, который стимулирует вегетативную нервную систему человека путем стимуляции потовых желез за счет стимулирования аксонных рефлексов. Кожу стимулируют электричеством, вызывая указанные рефлексы аксонов, что позволяет оценить тип и тяжесть вегетативных нервных расстройств и периферических невропатий, таких как астма или рассеянный склероз.
Реакция потоотделения
Люди и приматы используют судомоторную реакцию, чтобы вызвать терморегуляцию, или контроль температуры своего тела, в основном через симпатическую нервную систему. с незначительным влиянием со стороны парасимпатической нервной системы. Чувствительные к теплу рецепторы присутствуют в коже, внутренних органах и спинном мозге, где они получают информацию из внешней среды и отправляют ее в центр терморегуляции в гипоталамусе.
Реакция потоотделения стимулирует мускариновые рецепторы M3 на потовых железах и рефлекс судомоторного аксона. При судомоторном рефлексе холинергические агенты связываются с никотиновыми рецепторами на окончаниях судомоторного нерва, вызывая импульс, который движется к соме, или противоположный нормальному импульсу. В соме постганглионарного симпатического судомоторного нейрона импульс разветвляется и проходит ортодромно или от сомы. Наконец, когда этот импульс достигает других потовых желез, он вызывает непрямую рефлекторную реакцию потоотделения на аксоны. Рефлексы судомоторных аксонов могут периферически усиливаться при передаче величины потенциала действия с помощью ацетилхолина. Ацетилхолин также активирует судомоторные волокна и первичные афферентные ноцицепторы, вызывая рефлексы аксонов в обоих. Однако при повреждении нервов (невропатия ) наблюдается некоторое усиление потоотделения, опосредованного рефлексом аксонов.
Механизмы
Кожные рецепторы — это сенсорные рецепторы в коже, которые обнаруживают изменения температуры (терморецепторы ) и боли (ноцицепторы ). Эти кожные рецепторы инициируют импульс через возбуждение главного сенсорного аксона в спинной мозг. Рефлекс аксона — это распространение этого импульса от главного аксона к соседним кровеносным сосудам в стимулированном участке кожи. Эти импульсы в пораженной области высвобождают химические вещества, которые вызывают расширение кровеносных сосудов и утечку, вызывая потливость кожи. Высвобождается ацетилхолин, что приводит к увеличению внеклеточного кальция, что вызывает внеклеточную гиперполяризацию с последующим расширением артериолы. Покраснение приводит к обострению рефлекса аксона.
Этот механизм вазодилатации подтверждается исследованиями, а эффективность вазомоторного ответа можно объяснить значением тау (постоянной времени кровообращения). на эту область воздействует датчик). В общем, значение Tau не сильно меняется при температурах 39 ° C и выше, тогда как при температурах ниже 39 ° C значение Tau значительно варьируется. Сигнал, вызывающий вазодилатацию, исходит от повышения температуры кожи, приближающейся к порогу около 40 ° C. Фаза охлаждения тау будет зависеть от механики тела и способности человека излучать тепло от тела.
См. Также
- Axon
- Терморегуляция
- Гипотермия
- Обморожение
- Реакция охоты
Ссылки
ФИЗИКА ЖИЗНЕННЫХ ПРОЦЕССОВ
АКСОН И НЕРВНЫЙ ИМПУЛЬС
Генерация и распространение нервного импульса в организмах животных, возбуждение нервных и мышечных клеток — важнейшие мембранные явления. Мембранная теория возбуждения была сформулирована Бернштейном еще в 1902 г. [1]. Согласно его теории, возбуждение определяется электрохимическими процессами, локализованными в мембранах нервных и мышечных клеток. Эти процессы сводятся к перемещению малых ионов. Позднее Лазарев опубликовал монографию об ионной природе нервного возбуждения [2]. Исследования, проведенные Ходжки- ном, Катцем, Хаксли, Тасаки и другими учеными [3—6], раскрыли принципиальные механизмы генерации нервного импульса и его распространения (см. также [7—9]).
Нервное возбуждение распространяется по нервным волокнам— аксонам. Принято разделять нервную систему высших организмов на центральную и периферическую. Периферическая система содержит аксоны, служащие для передачи сигналов, а также ганглии вегетативной нервной системы. Аксоны являются коммуникациями для афферентных сообщений от органов чувств, направляемых в центральную систему, и для эфферентных сигналов, направляющихся от центральной системы к мышцам. Аксоны представляют собой отростки центрально расположенных клеток. Нервная система беспозвоночных имеет иное строение, но она также содержит коммуникационные аксоны.
Исследование генерации нервного импульса и его распространения в аксоне — старая и традиционная проблема биофизики/ Уже Гельмгольц измерял скорость распространения нервного возбуждения/В настоящее время решены важные физические задачи, относящиеся к функции аксона. Напротив, современное состояние науки позволяет лишь формально моделировать работу центральной нервной системы и мы еще далеки от понимания физической природы высших ее функций — памяти и мышления.
На рис. 4.1, заимствованном из [5], представлена схема строения нервной клетки, именуемой иначе нейроном. Нейрон полу
чает сообщения от многих других нейронов через их тонкие ответвления, образующие контакты — синапсы — с телом клетки и его короткими отростками — дендритами. Аксон имеет макроскопические протяженность и сечение. Длины аксонов в теле крупных животных достигают нескольких метров.
Контакты с другими клетками образуются не только в синапсах.^ Боль — шая часть поверхности нейрона покрыта прилегающими к нему глиальными, или иіванновскими, клетками^ роль которых пока не выяснена. С другой стороны^из мембран шванновских клеток образуется миелиновая оболочка миелинизирован — ных аксонов, показанная ^схематически на рис. 4.1. ^Оболочка прерывается через каждые 1—2 мм длины аксона перехватами Ранвье, имеющими протяженность /Т около 1 мкм. В области перехватов мембрана аксона непосредственно контактирует с окружающей средойЛ^
Немиелинизированные во — ЛОКНа могут быть объеди — Рис. 4.1. Схема строения нервной клетки нены одной шванновской (двигательный нейрон).
К-ТТР. ТКПЙ чтп ППКЯ1ЯНП R гр — і тн~тел0 нейрона,1Ца — аксои1Смо — миелиновая клеткой, ЧТО UOKdJdHU В се ~ оболочка, перехват Ранвье^д-дендрит,
ЧЄНИИ на рис. 4.2. Погру — X*— ядро,^с —синапсыЛодн—окончание двига о г — г тельного нерва,мв — мышечное волокно.
Женныи в шванНЬвскую 1

Клетку аксон создает глубокую инвагинацию мембраны шванновской клетки, причем образуется мезаксон — две шванновские мембраны, расположенные рядом^В правом нижнем углу схематически показана двойная мембрана, разделяющая цитоплазму шванновской клетки и цитоплазму аксона, аксоплазму [9].
Герен [10] установила, что миелиновая оболочка аксона образована многократным спиральным закручиванием мембраны шванновской клетки вокруг аксоиаГМиелин представляет собой, — т-а«им~ойразом,/многослойную (доЧїбО слоев) мембрану, строение которой наблюдается с помощью электронного микроскопа. Миелии защищает мембрану аксона от окружающей среды.
6 М. В. Волькеиштейн
Структура миелина была детально изучена методом рентгенографии с разрешением 10 А [11]. Определены профили электронной плотности миелина седалищного нерва кролика и лягушки и миелина зрительного нерва кролика. На рнс. 4.3 показаны эти кривые для нервных волокон кролика и схема структуры миелина, соответствующая данным кривым. Миелин построен асимметрично, холестерин сосредоточен преимущественно на стороне

Рис. 4.2. Схема строения немиелинизированнного нервного волокна позвоночного.
А — аксон, м — мезаксон— шванновская клетка.
Миелина, соприкасающейся с мембраной аксона. Схема согласуется с липидным составом миелина млекопитающих, установленным ранее [12]. Белковые слои граничных областей миелина содержат йоду. Внутренняя область построена из гибких углевОдорОдных «хвостов» липидов.
/Уже давно установлено, что нервы и мышцы способны генерировать электродвижущие силы, биопотенциалы. Электрическая активность клеток проявляется в форме коротких разрядов, каждый из которых длится около 1 мс. Современная экспериментальная техника позволяет усиливать эти сигналы и регистрировать их с помощью осциллографа или другим путем. Установлено, что активность нерва всегда сопровождается электрическими явлениями — и при внешнем возбуждении и при посылке сигнала мозгом в соответствующем аксоне наблюдаются электрические импульсы. На рис. 4.4 показаны импульсы в зрительном нервном волокні краба Limulus, вызванные вспышкой света длительностью 1 с [13]. Частота и характер последовательности импульсов, имеющие несомненное информационное значе
ние, зависят от интенсивности и спектрального состава света [14, 15] (см. гл. 7) .1 Величина и длительность отдельного импульса не зависят отТгрироды и силы раздражения.»
О 20 40А
-J 1 1 1 1

0 0 .::•:
![]()
»w0#0
^ V J 1 O^^-v^.p..
Г У. Го 0

Внугрен^/7о/тярже=Стероид щ^Гибх/гя ^Стероид ^//плярная Внешний
Нийбеяоцт. группы^ +ягесткая цепь +жесткая белок
+еоЗа ‘— » цепь — цепь — —~ +вода углеводороды
Рис. 4.3. Схема миелиновой мембранной структуры.
Вверх* — профили электронной плотности для глазного (пунктир) н седалищного (сплошная
Лнння) нервов кролика.
Электрический сигнал, отвечающий отдельному импульсу, распространяющемуся вдоль аксона, называется потенциалом действия или спайком. Это — основная единица информации, передаваемой по нервному волокну.

Скорость распространения импульса по аксону равна по порядку величины 1—100 м/с, она меньше для немиелинизиро — ванных и больше для миелинизированных аксонов.


0,01
0,001
Рис. 4.4. Импульсы в зрительном нервном волокне Limulus, вызванные вспышкой света длительностью I с.
Цифры справа указывают относительную интенсивность вспышки. Период освещения указан разрывом верхней белой линии, отметка времени на нижней белой линии 0,2 с.
В табл. 4.1 [3] приведены некоторые данные. Проведение нервного возбуждения есть проведение электрического импульса. Несмотря на то, что аксоплазма представляет
Таблица 4.1
Скорость проведения возбуждения в нервном н мышечном волокнах
|
Волокно |
Температура, °С |
Диаметр волокна мкм |
Скорость проведения, м/с |
|
Миелинизированные нервные волок |
38 |
2-20 |
10-100 |
|
На кошки |
|||
|
Немиелинизированные нервные во |
38 |
0,3-1,3 |
0,7—2,3 |
|
Локна кошки |
|||
|
Миелинизированные нервные волок |
20 |
3-16 |
6-32 |
|
На лягушки |
35 |
||
|
Миелинизированные нервные во |
20 |
20 |
|
|
Локна креветки |
|||
|
Немиелинизированные нервные во |
20 |
30 |
5 |
|
Локна краба |
|||
|
Гигантский аксон кальмара (немие- |
20 |
500 |
Около 25 |
|
Линизированное волокно) |
|||
|
Мышечное волокно лягушки |
20 |
60 |
1,6 |
‘) Для миелинизированных волокон указаны значения наружного диа^ метра миелиновой оболочки.
Собой раствор электролитов, никоим образом нельзя считать, что ионный ток в ней определяет распространение импульса. («Удельное сопротивление аксоплазмы равно по порядку величины 10—100 Ом-см. Следовательно, сопротивление на единицу длины волокна диаметром 1 мкм составляет 109—Ю10Ом-см, т. е. превышает сопротивление медного провода того же диаметра в 108 раз. В таком проводнике очень велики потери и утечки. Между тем аксон передает первичный импульс на расстояния свыше метра б^а-затухания. и без иска^кения.^.
В нейрофизиологии и в биофизике нервнЪ’го»»возбуждения сыграло большую роль развитие методов работы на изолированных аксонах, техники введения микроэлектродов в аксон и метода перфузии, т. е. выдавливания аксоплазмы из волокна и ее замены искусственными растворамиДОсобенно удобна работа на гигантских аксонах кальмара, имеющих диаметр до 0,5 мм (см. табл. 4.1 ^Элементарный опыт состоит во введении в аксон двух микроэлектродов: первый служит для электрической стимуляции возбуждения, второй-^- для измерения генерируемого потенциала (рис. 4.5) [4]./Опыт показывает, что «»величина и временной ход токов действия совершенно не зависят от величины стимулирующего тока и что потенциал действия не возникает, если электрический стимул не достигает некоторого порогового потенциала. Тем самым, нервное волокно
Гл. 4. нервный импульс
Подчиняется закону «все или ничего». Иными словами, потенциал действия постоянной величины либо возникает, либо не возникает вовсе.
Для возбуждения аксона необходимо некоторое минимальное количество электричества.,’ Соответственно по мере уменьшения
Раздражение Осцилмерар
ЕЖЖ^,..»,, ,
Аксон
Рис. 4.5. Схема исследования нервного импульса при помощи двух микроэлектродов.
Длительности At подаваемого импульса необходимо увеличивать силу тока / — эти две величины обратно пропорциональны. Для длительных прямоугольных импульсов существует минимальная
Сила тока, достаточная для возбуждения; более слабый ток неэффективен при любой длительности.
Fta рис. 3/S показана кривая зависимости силы порогового тока от длительности импульса. Кривая имеет гиперболическую форму. Было предложено описывать ее эмпирической формулой
/п = а/At — f b,
Где Іп— пороговый ток, At— длительность импульса, b — так называемая реобаза — минимальное значение возбуждающего тока при At-> оо. При очень коротких импульсах, т. е. при At—>-0 /п = а/At, т. е. произведение IaAt есть константа, характеризующая пороговое значение количества электричества при коротких импульсах. Из приведенной формулы следует, что при /п = 2b получим А^хр = а/Ь. В действительности, однако, оказалось, что при At = ajb /п~1,4Ь и простая эмпирическая формула несправедлива во всем интервале значений A^.j
166

Рисі 4.6. Зависимость силы порогового тока от длительности импульса.
Если сообщить волокну два последовательных стимула, разделенных некоторым интервалом времени, то возникающая ситуация зависит от этого интервала. Немедленно после генерации импульса данное место волокна находится в абсолютном
Рефрактерном состоянии, т. е. не может быть возбуждено вновь. Затем следует относительное рефрактерное состояние. В таком состоянии заметно увеличено значение порогового потенциала и может возникнуть лишь медленно распространяющийся импульс. Продолжительность всего рефрактерного состояния варьирует от одной до нескольких миллисекунд.
В состоянии покоя аксоплазма имеет отрицательный заряд по отношению к внешней среде, характеризуемый потенциалом покоя около 80 мВ. Пороговое возбуждение при этом имеет потенциал, примерно равный —50 мВ, а потенциал действия при надпороговом возбуждении равен потенциалу противоположного знака, составляющему около 40 мВ. Это изображено схематически на рис. 4.7 [4].
Прохождение импульса определяется изменением состояния мембраны аксона при ее стимуляции. Как уже говорилось, в состоянии покоя клеточной мембраны в результате активного транспорта ионов концентрация ионов К+ в цитоплазме (аксо — плазме) значительно превосходит их концентрацию во внешней среде, а для ионов Na+ ситуация противоположна^ В табл. 4.2 [3] приведен состав аксоплазми аксонов Loligo и состав плазмы крови, характерный для внешней среды, и близкий к нему состав морской воды.
Таблица 4.2 Концентрация нонов н других веществ в аксонах Loligo
Концентрация, ммоль на 1 кг НаО
|
Вещество |
Морская |
||
|
Аксоплазма |
Кровь |
Вода |
|
|
К+ |
400 |
20 |
10 |
|
Na+ |
50 |
440 |
460 |
|
Сг |
40—150 |
560 |
540 |
|
Са++ |
0,4 |
10 |
10 |
|
Mg++ |
10 |
54 |
53 |
|
Изэтионовая кислота |
250 |
— |
— |
|
Аспарагнновая кислота |
75 |
— |
— |
|
Глутаминовая кислота |
12 |
— |
— |
|
Янтарная н фумаровая кислоты |
17 |
— |
— |
|
Ортофосфат |
2,5-9 |
— |
— |
|
АТФ |
0,7—1,7 |
— |
— |
|
Аргннннфосфат |
1,8-5,7 865 г/кг |
— |
— |
|
Вода |
870 г/кг |
966 г/кг |
При наложении на мембрану стимулирующего потенциала вначале увеличивается ее проницаемость для ионов Na+, в состоянии покоя значительно меньшая проницаемости для ионов К+. Ионы Na+ входят в аксон, в результате чего внутренняя поверхность мембраны изменяет знак своего заряда с отрицательного на положительный. Иными словами, происходит деполяризация мембраныЛЕсли воспользоваться для выражения разности потенциалов на двух сторонах мембраны аксона формулой (3.105)
+ + (4.1)
^ РКСК + PNacNa + РС1сС ‘
То состояние покоя мембраны характеризуется следующим отношением ионных проницаемостей:
Рк ■ ^Na •’ Реї = 1 : 0,04:0,45.
В состоянии возбуждения, отвечающем генерации потенциала действия, имеем
Рк: Pnа: Ра = 1 :20: 0,45.
Подчеркнем, что истолкование возбуждения как результата изменения ионных проницаемостей является феноменологическим и не раскрывает молекулярный механизм процесса.
Наружная среда
![]()
Активный участок
+ + + — — — — + + + + +
Импульс Э-
Электрические токи, возникающие при деполяризации мембраны аксона.
При деполяризации мембраны возникают токи, замыкающиеся через наружную проводящую среду (рис. 4.8) [5]. Они возбуждают следующий участок аксона.
Таким образом, согласно мембранной теории при генерации импульса в мембране аксона открывается некий «канал», сквозь который ионы Na+ проникают внутрь аксона, вызывая деполяризацию мембраны. Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Ионы К+ выходят наружу, что приводит к восстановлению нормального отрицательного заряда на внутренней стороне мембраны. Происходит реполяризация мембраны. После прохождения рефрактерного периода мембрана приобретает способность передавать новый импульс (рис. 4.9) (4].
+40 О
-за
3
Оотенциол действия
Птещиал пот
+ к — — у + + + + + + + + + + + +
Їфі TtII ‘МрЖ. 111111
+ T —+ + + + + + + + + + + ®
+40
![]()
О
+ + + + + + + + І » + + + + + + + + + + + + + + ї—+ + + + + +
+40 О
-ВО

Рис. 4.9. Распространение нервного импульса сопровождается изменениями нонной проницаемости мембранного аксона.
Наружный раствор
/ / і ±
А/гсоплаэма
Рнс. 4.10. Эквивалентная схема элемента возбудимой мембраны аксона.
Как видно из рис. 4.8, суммарный продольный ток через сечение аксона и окружающую среду равен нулю — в любом месте внутренние токи равны по силе и противоположны по направлению. Но плотность продольного тока и продольная разность потенциалов между двумя точками внутри аксона иные, чем снаружи.
Мембрана аксона имеет сопротивление 1000 Ом-см2, ее емкость равна 1 мкФ/см2, что соответствует бимолекулярному липидному слою толщиной 50 А с диэлектрической проницаемостью е=5 и удельным сопротивлением 2-Ю9 Ом-см. Во время генерации импульса проводимость мембраны увеличивается примерно в 1000 раз. Можно моделировать электрические свойства мембраны эквивалентной схемой, показанной на рис. 4.10 [3]. Рисунок изображает лишь один элемент мембраны, и следует представить себе длинную линейную последовательность таких элементов, моделирующих непрерывный кабель. Сопротивление R характеризует аксоплазму, наружный раствор имеется в большом избытке и изображается проводником без сопротивления. Натриевая и калиевая «батареи» и S’k,
Определяют генерацию импульса, добавочная «батарея» Sy ответственна за движение других ионов, не изменяющееся при возбуждении.
Таким образом, распространение импульса представляет собой самоподдерживающийся процесс, подобный горению бикфордова шнура. Очевидно, что процесс этот сводится к превращению химической энергии в электрическую.
Как уже сказано, распространение импульса в миелинизированном волокне происходит быстрее, чем в немиелинизированном. Доказано, что в миелинизированном аксоне проведение импульса является сальтаторным, т. е. происходит путем перескока импульса от одного перехвата Ранвье к другому. Калиевые и натриевые каналы открываются и закрываются только в перехватах Ранвье — миелинизированные участки хорошо изолированы. Миелин имеет малую емкость, чем и объясняется большая скорость проведения импульса. Схема сальтаторного проведения импульса показана на рис. 4.11. ^
![]()
РирГ 4.Т7Т^ Схема сальтаторного проведения импульса. пР—перехват Ранвье, л —миелин, а — аксо-
Прямые доказательства сальтаторной їеории, впервые предложенной Лилли [16], были получены в работах Кубо, Оно и Юге (см. [17]). В одиночном нервно-мышечном препарате, погруженном в раствор Рингера, пороговая величина стиму
лирующего тока зависит от расстояния между стимулирующим микрозлектродом и перехватом Ранвье. Она имеет наименьшее значение, если микрозлектрод помещен непосредственно в перехвате. Количественная теория этого эффекта была развита Та — саки (см. [6]).
Обозначим потенциалы трех последовательных перехватов — 1, 0 и 1 буквами ifi_i, if)0 и ij)j. Если миелин представляет собой идеальный изолятор, то ток, протекающий внутри аксона от 1 к 0, пропорционален if>i— if>o, от 0 к —1 — пропорционален ■ф-i — if>o. Через перехват 0 идет ток І, пропорциональный (грі — ij)0) — f (j)_i — %,). Имеем
‘ = І{(+-,-*о) + (+,-+о)}. (4.2)
Где R — сопротивление между двумя соседними перехватами (оно равно примерно 20 МОм).
Расстояния между микроэлектродом и перехватами 0, 1 и —1 равны Хо, Х, х-. Разность потенциалов, создаваемая микроэлектродом, дающим ток I, равна
Где р — удельное сопротивление среды (для раствора Рингера оно равно 100 Ом-см). Из (4.2) и (4.3) получаем
TOC o «1-3» h z i = Ч (4-4)
4яR х0 х х-1 ) 4 ‘
И если микроэлектрод очень близок к перехвату 0, т. е. Хи Х-1 » *0, то
= (4-5)
В опытах варьировалось расстояние х0 и при каждом значении Хо определялась величина /, отвечающая критическому значению і — іс. Опытные данные действительно согласуются с линейной зависимостью I от х0
І= — Ісх0. (4.6)
Установлено, что факторы, блокирующие распространение импульса (кокаин, уретан, ультрафиолетовое облучение), действуют именно на перехваты Ранвье, но не на участки между ними.
Прямые и строгие доказательства сальтаторной теории получены в работах Тасаки и его сотрудников [6, 18, 19]. Основной опыт ставился на одиночном волокне, лежащем в трех каплях раствора Рингера, разделенных двумя воздушными промежутками, расстояние между которыми равнялось приблизительно 1 мм (рис. 4.12) [3]. В первом опыте (рис. 4.12, а) в средней капле находилась только миелинизированная часть аксона, во втором (рис. 4.12,6)—в средней капле находился перехват Ранвье. Измерялся радиальный ток. Через сопротивление R проходит ток, равный il2— г23, где i2— ток, текущий в наружной цепи от капли 1 к капле 2, і23— от капли 2 к капле 3. Эта разность равна току, входящему в волокно из капли 2.

1ме 1ма
Рис. 4.12. Мембранный ток:
А—через участок аксона, расположенный между перехватами Раивье, б —через перехват Ранвье. Отклонение осциллограммы вверх соответствует выходящему току.
В первом опыте радиальный ток направлен наружу, что объясняется пассивным разрядом миелинизированного участка через перехват. Правый перехват разряжается позже левого, соответственно на осциллограмме видны два пика. Во втором опыте первая фаза тока направлена наружу, что означает пассивную деполяризацию перехвата. Во второй фазе ток направлен внутрь — перехват Ранвье находится в активном состоянии. Эти опыты однозначно подтверждают теорию (дальнейшие подробности см. в [3, 6, 18, 19]).
Мембранная теория в целом непосредственно подтверждается опытами с перфузией аксонов. Оказалось возможным выдавить аксоплазму из гигантского аксона Loligo без повреждения мембраны и заменить аксоплазму искусственными растворами (см. [3, 6, 20—22]). Перфузированные аксоны способны проводить до 105—106 импульсов в течение нескольких часов.
Потенциал покоя исчезает при равенстве наружной и внутренней концентраций калия. При замене в аксоплазме КС1 на NaCl потенциал покоя падает до нуля. В то же время потенциал покоя малочувствителен к концентрациям К+, меньшим 20 мМ (см. также § 3.6). Перфузионные опыты показывают, что потенциал покоя действительно регулируется ионами калия.
При указанной замене К. С1 на NaCl и значительном увеличении концентрации К (до 600 мМ) создается положение, обратное нормальному, и внутренняя часть волокна заряжается положительно, а не отрицательно по отношению к наружной среде. И в самом деле, при заполнении волокна, погруженного в изотонический раствор КС1, таким же раствором NaCl внутренний раствор заряжен положительно по отношению к наружному и <р составляет 50—60 мВ.
Прямые опыты с мечеными атомами показывают, что проведение импульса связано с возрастанием скорости движения калия и натрия по градиентам концентрации. В гигантском аксоне каракатицы Sepia при каждом импульсе наблюдается входящий поток Na+, равный 10,3-Ю-12 моль/см2, и выходящий поток 6,6-Ю-12 моль/см2. Чистый вход Na+ 3,7-Ю-12 моль/см2 за импульс примерно равен выходу К+. Действительно, для изменения напряжения конденсатора емкостью 1 мкФ на 120 мВ нужен заряд 0,12-Ю-6 Кл, что эквивалентно 1,3-Ю-12 моль/см2 одновалентного катиона. Таким образом, измеренный вход Na+ более чем достаточен для возникновения потенциала действия.
За один импульс в немиелинизированном аксоне через 1 мкм2 поверхности проходит около 20 000 ионов Na+. В миелинизиро — ванном аксоне за один импульс через каждый перехват Ранвье входит 6-Ю6 ионов Na+. Площадь мембраны в перехвате примерно равна 20 мкм2, следовательно, через 1 мкм2 проходит 300 000 ионов Na+. Плотность ионного тока в перехвате примерно в 10 раз больше, чем в немиелинизированных гигантских аксонах.
Мембранная теория встречается, однако, с трудностями при попытках объяснить тепловые явления в нерве. Согласно мембранной теории генерация импульса происходит за счет свободной энергии, определяемой градиентом концентрации ионов Na+. Если перенос ионов адиабатичен, то система, совершая электрическую работу, должна охлаждаться. Однако оказалось, что во время генерации импульса сначала происходит нагревание нерва, за которым следует охлаждение [23]. Аналогичное явление наблюдается при разряде электрического органа угря на внешнее сопротивление [24]. Эти тепловые явления до сих пор не объяснены. Несомненно, что мембрана аксона представляет собой диссипативную систему, и можно думать, что к рассмотрению генерации нервного импульса применима неравновесная термодинамика. Соответствующая теория еще не построена, она должна описать и теплопродукцию нерва. Подлинное объяснение этих явлений требует, однако, раскрытия их молекулярных механизмов.
В заключение приведем электрические характеристики мие- линизированного нервного волокна лягушки (табл. 4.3) [3].
Таблица 4.3
Электрические характеристики аксона
|
Диаметр волокна |
14 мкм |
|
Толщина миелиновой оболочки |
2 мкм |
|
Расстояние между перехватами Ранвье |
2 мм |
|
Площадь мембраны в перехвате Ранвье |
22 мкм2 |
|
Сопротивление на единицу длины осевого ци |
140 МОм/см |
|
Линдра |
110 Ом • см |
|
Удельное сопротивление аксоплазми |
|
|
Емкость на единицу длины миелиновой оболочки |
10—16 пФ/см |
|
Емкость на едниицу площади миелиновой обо |
0,0025—0,005 мкФ/см2 |
|
Лочки |
|
|
Диэлектрическая проницаемость миелиновой обо |
5-10 |
|
Лочки |
|
|
Удельное сопротивление миелиновой оболочки |
500—800 МОм/см |
|
Емкость перехвата Ранвье |
0,6-1,5 пФ |
|
Емкость на единицу площади перехвата Ранвье |
3—7 мкФ/см2 |
|
Сопротивление перехвата Ранвье в покое |
40—80 МОм |
|
Сопротивление, умноженное на единицу площади |
10-20 Ом-см2 |
|
Мембраны в перехвате Ранвье |
|
|
Потенциал действия |
116 мВ |
|
Потенциал покоя |
71 мВ |
|
Максимальная плотность входящего тока |
20 мА/см2 |
|
Скорость проведения |
23 м/с |
В биологии особое значение имеют автокаталитические химические системы. Достаточно указать, что авторепродукция КДеток и организмов эквивалентна автокатализу. Вернемся сначала к феноменологическому термодинамическому рассмотрению. Как мы видели, для химических процессов критерий …
Неотъемлемой особенностью биологических объектов — клеток и организмов — является их историчность, т. е. возникновение и развитие изучаемой системы в конечном интервале времени. Развитие биологической системы всегда необратимо, и в …
Самые общие соображения показывают, что перенос электрона, сдвиг электронной плотности в конденсированной ср. еде должны сопровождаться изменениями положений атомов, атомных ядер среды. Все степени свободы молекулярной системы, т. е. системы, …
Аксон
-
Аксон (др.-греч. ἄξων «ось») — это нейрит (длинный цилиндрический отросток нервной клетки), по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.
Каждый нейрон состоит из одного аксона, тела (перикариона) и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные или мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону, а затем сгенерированный потенциал действия от начального сегмента аксона передаётся назад к дендритам. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС).
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
Источник: Википедия
Связанные понятия
Си́напс (греч. σύναψις, от συνάπτειν — соединение, связь) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём, посредством прохождения ионов из одной клетки в другую.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться один с другим, формируя биологические…
Дендрит (от греч. δένδρον (dendron) — дерево) — разветвлённый отросток нейрона, который получает информацию через химические (или электрические) синапсы от аксонов (или дендритов и сомы) других нейронов и передаёт её через электрический сигнал телу нейрона (перикариону), из которого вырастает. Термин «дендрит» ввёл в научный оборот швейцарский ученый В. Гис в 1889 году.
Миелин (в некоторых изданиях употребляется некорректная теперь форма миэлин) — вещество, образующее миелиновую оболочку нервных волокон.
Вставочный нейрон (синонимы: интернейрон, промежуточный нейрон; англ. interneuron, relay neuron, association neuron, bipolar neuron) — нейрон, связанный только с другими нейронами, в отличие от двигательных нейронов, иннервирующих мышечные волокна, и сенсорных нейронов, преобразующих стимулы из внешней среды в электрические сигналы.
Упоминания в литературе
Аксон — длинное нервное волокно, отходящее от тела некоторых нейронов и заканчивающееся терминалями (до нескольких сотен) на клетках-мишенях: нейронах, миоцитах, клетках железы и др. В зависимости от наличия или отсутствия аксона выделяют нейроны I и II типа. Область тела нейрона, которой начинается аксон, называют начальным сегментом аксон, или аксонным холмиком. Мембрана аксонного холмика генерирует потенциалы действия (нервные импульсы), распространяющиеся по аксону. Некоторые аксоны покрыты прерывистыми цепочками миелиновых оболочек (муфт) и называются миелинизированными (волокна спинномозговых нервов). Аксоны, свободные от миелина, называются немиелинизированными (волокна вегетативных нервов). Между муфтами имеются промежутки – перехваты Ранвье. В участках, лишенных миелиновой оболочки, мембрана аксона контактирует непосредственно с внеклеточной средой. В ЦНС миелиновые муфты образованы глиальными клетками, олигодендритами. Миелиновые муфты аксонов, выходящих за пределы ЦНС (периферических волокон), образованы шванновскими клетками. Одна шванновская клетка может образовывать муфты, покрывая до девяти нервных волокон. Главная функция аксонов – передача нервных импульсов от тела нейрона к терминалям, которые заканчиваются на клетках-мишенях утолщениями, синаптическими бляшками. Скорость движения информации по аксонам составляет от нескольких миллиметров в секунду в мелких немиелинизированных волокнах до 90 м/с в крупных миелинизированных. Миелиновая оболочка увеличивает скорость передачи потенциалов действия по аксону.
НЕЙРО́Н, нервная клетка; основная структурная и функциональная единица нервной системы. Нейроны обладают особыми свойствами – возбудимостью и проводимостью. Из нейронов построена нервная ткань. Как и другие клетки, нейрон имеет ядро, цитоплазму и клеточную мембрану. Ядро содержится в теле нейрона (его расширенная часть), от которого отходят отростки – короткие (дендриты) и один длинный (аксон). Длина аксона может достигать 1 м и более. Аксоны образуют тяжки, которые называются нервами (наиболее крупные – нервными стволами). Дендриты принимают сигналы из внешней среды или от другой нервной клетки и проводят возбуждение к аксону, который передаёт его в удалённые от тела нейрона области. Тела нейронов образуют серое вещество головного и спинного мозга, а дендриты и аксоны – белое мозговое вещество.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка – интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Нейрон. Основной функциональной и структурной единицей нервной системы является нейрон, строение которого показано на рис. 1.1. Нейрон состоит из клеточного тела, или сомы, коротких нервных отростков, называемых дендритами, и длинного нервного волокна – аксона. Область выхода аксона из клеточного тела нейрона называется аксонным холмиком. Хотя формально и дендриты и аксоны являются нервными волокнами, термин «нервное волокно» в основном употребляется при упоминании аксона. Дендриты передают нервный импульс к телу нейрона, тогда как аксон направляет импульс из тела нейрона на другие нервные клетки или на мышечные волокна.
Связанные понятия (продолжение)
Клетки Пуркинье (англ. Purkinje cells) — крупные нервные клетки коры мозжечка. Своё название клетки получили в честь их первооткрывателя, чешского врача и физиолога Яна Эвангелисты Пуркинье.
Пирамидальные нейроны, или пирамидные нейроны, — основные возбудительные нейроны мозга млекопитающих. Также обнаруживаются у рыб, птиц, рептилий. Напоминают по форме пирамиду, из которой вверх ведёт большой апикальный дендрит; имеют один аксон, идущий вниз, и множество базальных дендритов. Впервые были исследованы Рамон-и-Кахалем. Отмечены в таких структурах, как кора мозга, гиппокамп, миндалевидное тело (амигдала), но отсутствуют в обонятельной луковице, стриатуме, среднем мозге, ромбовидном мозге…
Подробнее: Пирамидальный нейрон
Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов. Термин ввёл в 1846 году Рудольф Вирхов.
Перикарион (др.-греч. περι- — приставка со значением «около, вокруг, кругом» + κάρυον — «орех») — сома (тело) нейрона, может иметь различную величину и форму. На цитолемме перикариона образуются многочисленные синаптические контакты с отростками других нейронов.
Нервная ткань — ткань эктодермального происхождения, представляет собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию.
Мотонейро́н (от лат. motor — приводящий в движение и нейрон; двигательный нейро́н) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса.
Шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Астроцит (лат. astrocytus; от греч. astron — звезда; и kýtos, здесь — клетка) — тип нейроглиальной клетки звездчатой формы с многочисленными отростками. Совокупность астроцитов называется астроглией.
Гранулярные клетки — несколько разновидностей мелких нейронов мозга. Название «гранулярная клетка» («зернистая клетка», «клетка-зерно») используется анатомами для нескольких разных типов нейронов, единственной общей особенностью которых является крайне малый размер тел этих клеток.
Подробнее: Гранулярная клетка
Гипотала́мус (лат. hypothalamus, от греч. ὑπό — «под» и θάλαμος — «комната, камера, отсек, таламус») — небольшая область в промежуточном мозге, включающая в себя большое число групп клеток (свыше 30 ядер), которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма. Гипоталамус связан нервными путями практически со всеми отделами центральной нервной системы, включая кору, гиппокамп, миндалину, мозжечок, ствол мозга и спинной мозг. Вместе с гипофизом гипоталамус образует гипоталамо-гипофизарную…
Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие.
Корзинчатые нейроны — тормозные ГАМК-эргические вставочные нейроны молекулярного слоя мозжечка. Длинные аксоны корзинчатых нейронов образуют корзиноподобные синапсы с телами клеток Пуркинье. Корзинчатые нейроны многополярны, их дендриты свободно ветвятся.
Подробнее: Корзинчатый нейрон
Тала́мус, иногда — зри́тельные бугры (лат. Thalamus; от др.-греч. θάλαμος — «камера, комната, отсек») — отдел головного мозга, представляющий собой большую массу серого вещества, расположенную в верхней части таламической области промежуточного мозга хордовых животных, в том числе и человека. Впервые описан древнеримским врачом и анатомом Галеном. Таламус — это парная структура, состоящая из двух половинок, симметричных относительно межполушарной плоскости. Таламус находится глубже структур большого…
Полоса́тое те́ло (лат. corpus striatum) — анатомическая структура конечного мозга, относящаяся к базальным ядрам полушарий головного мозга.
Гиппока́мп (от др.-греч. ἱππόκαμπος — морской конёк) — часть лимбической системы головного мозга (обонятельного мозга). Участвует в механизмах формирования эмоций, консолидации памяти (то есть перехода кратковременной памяти в долговременную). Генерирует тета-ритм при удержании внимания.
Латеральное коленчатое тело (наружное коленчатое тело, ЛКТ) — легко распознаваемая структура мозга, которая помещается на нижней латеральной стороне подушки таламуса в виде достаточно большого плоского бугорка. В ЛКТ приматов и человека морфологически определено шесть слоев: 1 и 2 — слои крупных клеток, 3-6 — слои мелких клеток. Слои 1, 4 и 6 получают афференты от контрлатерального (расположенного в противоположном по отношению к ЛКТ полушарии) глаза, а слои 2, 3 и 5 — от ипсилатерального (расположенного…
Зубчатая извилина (лат. gyrus dentatus) или зубчатая фасция гиппокампа (лат. fascia dentata hippocampi) — зазубренная извилина, расположенная в глубине борозды гиппокампа и переходящая в ленточную извилину. В некоторых классификациях она вместе с аммоновым рогом считается частью самого гиппокампа, однако большинство авторов относят её к гиппокамповой формации. В её структуре выделяют три слоя: полиморфный хилус, гранулярный слой и молекулярный слой, который непрерывно переходит в молекулярный слой…
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в виде кратковременного изменения мембранного потенциала на небольшом участке возбудимой клетки (нейрона или кардиомиоцита), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к внутренней поверхности мембраны, в то время, как в покое она заряжена положительно. Потенциал действия является физиологической основой нервного импульса.
Обонятельная луковица — часть обонятельного мозга, парное образование, состоящее из тел вторых нейронов биполярного типа обонятельного анализатора. Располагается во внутричерепной полости между лобной долей сверху и решётчатой пластинкой решётчатой кости снизу, через отверстие которой в неё поступают нервные волокна обонятельной области носа, а сзади продолжается в обонятельный тракт.
Реце́птор — объединение из терминалей (нервных окончаний) дендритов чувствительных нейронов, глии, специализированных образований межклеточного вещества и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражитель) в нервный импульс. В некоторых рецепторах (например, вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками эпителиального происхождения или…
Ганглий (др.-греч. γάγγλιον — узел), или нервный узел — скопление нервных клеток, состоящее из тел, дендритов и аксонов нервных клеток и глиальных клеток. Обычно ганглий имеет также оболочку из соединительной ткани. Имеются у многих беспозвоночных и всех позвоночных животных. Часто соединяются между собой, образуя различные структуры (нервные сплетения, нервные цепочки и т. п.).
Голубое пятно (голубоватое место, голубоватое пятно, синее пятно/место, лат. locus coeruleus) — ядро, расположенное в стволе мозга на уровне моста (участок голубоватого цвета в верхнелатеральной части ромбовидной ямки ствола головного мозга кнаружи от верхней ямки), часть ретикулярной формации. Система его проекций очень широка — аксоны восходят к верхним слоям коры больших полушарий, гиппокампу, миндалине, перегородке, полосатому телу, коре мозжечка. Нисходящие проекции идут в спинной мозг к симпатическим…
Афферентация (от лат. afferens — «приносящий») — постоянный поток нервных импульсов, поступающих в центральную нервную систему от органов чувств, воспринимающих информацию как от внешних раздражителей (экстерорецепция), так и от внутренних органов (интерорецепция). Находится в прямой зависимости от количества и силы воздействующих раздражителей, а также от состояния — активности или пассивности — индивида.
Белое вещество (лат. substantia alba) — компонент центральной нервной системы позвоночных животных и человека, состоящий главным образом из пучков аксонов, покрытых миелином. Противопоставляется серому веществу мозга, состоящему из клеточных тел нейронов. Цветовая дифференциация белого и серого вещества нервной ткани обусловлена белым цветом миелина.
Нервно-мышечный синапс (также нейромышечный, либо мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в этом синапсе является ацетилхолин.
Ретикулярная формация (лат. reticulum — сеточка, formatio — образование) — это образование, тянущееся вдоль всей оси ствола головного мозга. Своим названием оно обязано сетчатой структуре, образуемой его нервными клетками с очень сложными связями. Формация состоит из ретикулярных ядер и большой сети нейронов с разветвлёнными аксонами и дендритами, представляющих единый комплекс, который осуществляет активацию коры головного мозга и контролирует рефлекторную деятельность спинного мозга. Эта сеть нейронов…
Средний мозг, или мезэнцефалон (англ. Midbrain, лат. Mesencephalon; термин «мезэнцефалон» происходит от др.-греч. μέσος — «месос» — средний, и ἐγκέφᾰλος — «энкефалос» — буквально «находящийся внутри головы», то есть головной мозг) — это отдел головного мозга хордовых животных, развивающийся из среднего из трёх первичных мозговых пузырей эмбриона. Этот отдел мозга ответствен за осуществление многих важных физиологических функций, таких, как зрение, слух, контроль движений, регуляция циклов сна и бодрствования…
Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
Мозгово́й ствол, или ствол головного мозга, — традиционно выделяющийся отдел третьего мозга, представляющий собой протяжённое образование, продолжающее спинной мозг.
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг…
Конечный мозг (лат. telencephalon) — самый передний отдел головного мозга. Состоит из двух полушарий большого мозга (покрытых корой), мозолистого тела, полосатого тела и обонятельного мозга. Является наиболее крупным отделом головного мозга. Это также самая развитая структура, покрывающая собой все отделы головного мозга.
Дендритный шипик — мембранный вырост на поверхности дендрита, способный образовать синаптическое соединение. Шипики обычно имеют тонкую дендритную шейку, оканчивающуюся шарообразной дендритной головкой. Дендритные шипики обнаруживаются на дендритах большинства основных типов нейронов мозга. В создании шипиков участвует белок калирин.
Эпиталамус (буквально «надталамус») — это самая дорсальная (верхняя) задняя часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо эпиталамуса, входят также таламус, субталамус и метаталамус, но не входят гипоталамус и гипофиз, причисляемые к гипоталамической области. Эпиталамус возвышается над таламусом. В число его структур входят поводок эпиталамуса, также называемый поводком мозга, треугольник поводка, спайка поводков, подспаечный орган…
Передний мозг, или прозэнцефалон (лат. prosencephalon, англ. forebrain) — это отдел головного мозга хордовых животных, выделяемый на основании особенностей его эмбрионального развития у этих видов животных. Передний мозг (прозэнцефалон) является одним из трёх первичных мозговых пузырей, образующихся в первичной нервной трубке вскоре после нейруляции и образования нейромер, на так называемой трёхпузырьковой стадии развития ЦНС эмбриона. Двумя другими первичными мозговыми пузырями являются средний…
Миоци́ты, или мы́шечные клетки — особый тип клеток, составляющий основную часть мышечной ткани. Миоциты представляют собой длинные, вытянутые клетки, развивающиеся из клеток-предшественников — миобластов. Существует несколько типов миоцитов: миоциты сердечной мышцы (кардиомиоциты), скелетной и гладкой мускулатуры. Каждый из этих типов обладает особыми свойствами. Например, кардиомиоциты, помимо прочего, генерируют электрические импульсы, задающие сердечный ритм.
Головно́й мозг (лат. cerebrum, др.-греч. ἐγκέφαλος) — главный орган центральной нервной системы подавляющего большинства хордовых, её головной конец; у позвоночных находится внутри черепа. В анатомической номенклатуре позвоночных, в том числе человека, мозг в целом чаще всего обозначается как encephalon — латинизированная форма греческого слова; изначально латинское cerebrum стало синонимом большого мозга (telencephalon).
Спинно́й мозг (лат. medulla spinalis) — орган центральной нервной системы позвоночных, расположенный в позвоночном канале. Принято считать, что граница между спинным и головным мозгом проходит на уровне перекреста пирамидных волокон (хотя эта граница весьма условна). Внутри спинного мозга имеется полость, называемая центральным каналом (лат. canalis centralis). Спинной мозг защищён мягкой, паутинной и твёрдой мозговой оболочкой. Пространства между оболочками и спинномозговым каналом заполнены спинномозговой…
Бледный шар (лат. globus pallidus s. pallidum) — парная структура переднего мозга, относящаяся к базальным ядрам, часть чечевицеобразного ядра, вентромедиальная часть полосатого тела. Подразделяется на латеральную и медиальную части.
Промежуточный мозг, или диэнцефалон (лат. Diencephalon, англ. Diencephalon; термин «диэнцефалон» происходит от др.-греч. διά — «диа-», обозначающее «через», «между», и ἐγκέφαλος — «энкефалос», буквально «находящийся внутри головы», то есть головной мозг) — отдел головного мозга хордовых животных, который образуется в процессе эмбрионального развития из задней части зародышевого переднего мозга (прозэнцефалона). На пятипузырьковой стадии из задней части зародышевого переднего мозга (прозэнцефалона…
Олигодендроциты, или олигодендроглия — это вид нейроглии, открытый Пио дель Рио-Ортегой (1928 год). Олигодендроциты есть только в центральной нервной системе, которая у позвоночных включает в себя головной мозг и спинной мозг.
Синаптогенез — процесс формирования синапсов между нейронами в нервной системе. Синаптогенез происходит на протяжении всей жизни здорового человека, а взрыв формирования синапсов, т. н. избыточный синаптогенез (exuberant synaptogenesis), наблюдается на ранних стадиях развития головного мозга. Синаптогенез особенно важен в ходе критического периода развития особи (в биологии развития, такого периода, когда нервная система особенно чувствительна к экзогенным стимулам), когда имеет место быть интенсивное…
Хвостатое ядро (лат. nucleus caudatus) — парная структура головного мозга, относящаяся к стриатуму. Расположена спереди от таламуса, от которого (на горизонтальном срезе) его отделяет белая полоска вещества — внутренняя капсула. Передний отдел хвостатого ядра утолщён и образует головку, caput nuclei caudati, которая составляет латеральную стенку переднего рога бокового желудочка. Головка хвостатого ядра примыкает внизу к переднему продырявленному веществу, в этом месте головка соединяется с чечевицеобразным…
Продолговатый мозг (лат. myelencephalon, medulla oblongata), или луковица головного мозга (лат. bulbus cerebri), — задний отдел головного мозга, непосредственное продолжение спинного мозга. Происходит из ромбовидного мозга и входит в ствол головного мозга. Регулирует такие основные процессы жизнедеятельности, как дыхание и кровообращение, поэтому в случае повреждения продолговатого мозга мгновенно наступает смерть.
Химический синапс — особый тип межклеточного контакта между нейроном и клеткой-мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество.
Электри́ческий си́напс (англ. electrical synapse) — место высокоспециализированных контактов (щелевых контактов) между нейронами, где происходит прямое перетекание электрических токов от одного нейрона к другому. В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм, в то время как в химическом синапсе расстояние между двумя нейронами составляет от 20 до 40 нм. У многих животных в нервной системе имеются как химические, так и электрические синапсы. По сравнению с химическими…
Упоминания в литературе (продолжение)
Основная единица нервной системы — нервная клетка (нейрон). Нейроны покрыты глиальными клетками, доставляющими им питание и удерживающими их на месте. Каждая нервная клетка состоит из тела, дендрита и аксона. В периферической нервной системе аксоны собраны в пучки – нервы. Бо́льшая часть аксонов покрыта миелиновой оболочкой, которая увеличивает скорость передачи информации в нервной системе, а также препятствует передаче нервных импульсов близлежащим клеткам. Современные исследователи опровергли бытовавшее ранее мнение о том, что «нервные клетки не восстанавливаются». Доказано, что клетки в зонах мозга, связанных с памятью и процессом обучения, продолжают развиваться и делиться в течение жизни человека.
Нервные клетки имеют несколько отростков – чувствительных древовидно ветвящихся дендритов, которые проводят к телу нейронов возбуждение, возникающее на их чувствительных нервных окончаниях, расположенных в органах, и одних двигательный аксон, по которому нервный импульс передается от нейрона к рабочему органу или другому нейрону. Нейроны вступают друг с другом в контакт с помощью окончаний отростков, образуя рефлекторные цепи, по которым передаются (распространяются) нервные импульсы.
От нервных центров по основному отростку (аксону) идут сигналы к каждой клетке и каждому органу тела, заставляя их путем электрической стимуляции выполнять определенную функцию. Нервные центры состоят из сотен и даже тысяч нервных клеток. Соответственно, существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Основной ее элемент – нейрон: микроскопически малая нервная клетка с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Вспомогательные клетки нейроглии (астроциты) не только создают физическую опору для нейронов, но вместе с сосудами обеспечивают потребности нервной ткани в кислороде и необходимых для жизни веществах, включая аминокислоты, липиды, гликопротеиды. Тело нервной клетки имеет микроскопические размеры, но длина аксона может достигать одного метра! Отростки нейронов, как правило, укрыты миелиновой оболочкой, которая обеспечивает стабильность обмена веществ в длинных нервных проводниках и высокую скорость передачи возбуждения.
Нейрон представляет собой микроскопически малую нервную клетку с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Главная структурная и функциональная единица нервной системы – нейрон, представляющий собой клетку с отростками – длинным аксоном и короткими дендритами. Нейроны соединяются между собой синапсами, образуя нейронные цепи, приводимые в действие рефлекторно: в ответ на раздражение, поступаемое из внешней или внутренней среды, возбуждение из нервных окончаний передается по центростремительным волокнам в головной и спинной мозг, оттуда импульсы по центробежным волокнам поступают в различные органы, а по двигательным – к мышцам.
Аксон – вырост цитоплазмы, приспособленный для проведения информации, которая собирается ден-дритами и перерабатывается в нейроне. Аксон дендритной клетки имеет постоянный диаметр и покрыт миелиновой оболочкой, которая образована из глии, у аксона разветвленные окончания, в которых находятся митохондрии и секреторные образования.
Два слова о строении нейрона. У него есть центральная толстенькая часть – «тело», в котором находится клеточное ядро с генами. От тела отходят два вида отростков: «входные» (дендриты) и «выходные» (аксоны). Дендритов обычно много, а аксон, как правило, один, но на конце он может ветвиться.
При поражении центров коры головного мозга очень важна локализация, ведь повреждения происходят на микроскопическом уровне. Головной мозг состоит из нейронов и глиальных клеток, которые соединены между собой сложными проводимыми путями – аксонами и дендритами. По ним передаются нервные импульсы. Все мы еще со школьного курса биологии знаем, что в головном мозге есть серое и белое вещества. Серым веществом является скопление нейронов и коротких проводимых путей между ними. А вот белое вещество составляют длинные, сложные проводимые пути – нервные волокна, которые покрыты особой миелиновой оболочкой – она необходима для нормального прохождения импульсов и подобна изоляционной ленте на электрическом проводе.
В состоянии относительного физиологического покоя синапс находится в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается и ее содержимое в виде одного кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
Таким образом, нейрофизиологические и психофизиологические механизмы tDCS и ТКМП достаточно многообразны, даже при приложении стандартизированных параметров тока и площади электродов (при tDCS). Нам представляется маловероятным сведение механизмов поляризаций к делоляризации или гиперполяризации мембраны нейронов. Если для пирамидных нейронов с их пространственной ориентацией в коре можно себе представить, что, например, при анодной поляризации входящий (гиперполяризующий) ток протекает преимущественно через дендрита, а выходящий (деполяризующий) ток – через начальный сегмент аксона (где генерируются спайки), вызывая его деполяризацию и увеличение возбудимости моторной коры, то это допущение плохо приложимо к другим областям коры, где ориентация нейронов не столь очевидна. Кроме того, возбуждающие или тормозные потенциалы длятся не более 10 мсек. Пассивная деполяризация мембраны еще короче. Установленные эффекты tDCS длятся десятки минут, а то и дольше. Следовательно, де– или гиперполяризация, если и играют существенную роль в возникновении эффектов, то она сводится к пусковой роли, запускающей каскад синаптических и метаболических реакций которые и лежат в основе более длительных эффектов поляризаций.
Надо сказать, что нейрон устроен очень интересно: у него, как у всякого компьютера, есть корпус – тело. И все же славится нейрон не этим, главное в нем – это его отростки. Весь нейрон чем-то напоминает дерево, которое имеет развитую крону и ствол. Крону у дерева образуют ветки, а у нейрона это короткие отростки, они называются дендритами. У дерева ствол, а у нейрона – длинный отросток, аксон. А теперь о месте соединения нервных клеток – это синапс. Если два компьютера соединить шнуром, то контакт компьютера и шнура будет как раз тем самым синапсом в нервной системе.
Активность генов в нейронах основана на свойствах молекул ДНК, т. е. активность вызвана причинно-следственными связями, которые определяют особенности контактов молекул ДНК с окружающими атомами и молекулами. В свою очередь, активность генов является причиной всех событий клеточного метаболизма и, значит, обмена веществ между нейронами и с окружающей средой. Все процессы в сообществе нервных клеток (клеточном сообществе – КС) – рождение клеток, их возбуждение и торможение, функциональная специализация и формирование в них устойчивых молекулярных структур, синтез в клетках нейромедиаторов и образование у них аксонов, дендритов и шипиков, образование и разрыв межклеточных связей, распространение и прекращение возбуждений в сообществе, гибель клеток – это детерминистские процессы. Так у нейронов в течение суток изменяются параметры метаболизма: количество и соотношение синтезируемых и разлагающихся веществ, в т. ч. количество синтезируемых нейромедиаторов и гормонов. У нейронов, как и у всех организмов, имеются суточные ритмы. В соответствии с этими ритмами в продолжение суток изменяются состав и количество клеток, возбуждающихся от воздействий окружающей среды и следов таких воздействий, результаты конкуренции клеток и узлов за прекращение своего возбуждения, используемые возбуждениями маршруты связей между клетками, конфигурация формирующихся способов прекращения возбуждений, распространяющихся из одних и тех же узлов.
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь. Коллатерали, идущие от задних корешков шейных сегментов, входят в состав клиновидного пучка, идут вверх к его ядру и к дополнительному клиновидному ядру. Его аксоны соединяются с мозжечком. Третья группа коллатеральных афферентных волокон заканчивается в задних рогах спинного мозга. Там расположены вторые нейроны, чьи аксоны образуют передний спино-мозжечковый путь.
Соответственно существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Двигательная единица – это отдельный мотонейрон и мышечные волокна, которые он иннервирует. Таким образом, нейрон определяет, являются ли волокна медленно или быстро сокращающимися. Мотонейрон в МС двигательной единице имеет небольшое клеточное тело и иннервирует группу из 10-180 мышечных волокон. У мотонейрона в БС двигательной единице большое клеточное тело и больше аксонов, и он иннервирует от 300 до 800 мышечных волокон. Отсюда следует, что каждый МС-мотонейрон в состоянии активировать значительно меньшее количество мышечных волокон в противоположность БС-мотонейрону. При этом необходимо отметить, что сила, производимая отдельными МС- и БС-волокнами по величине отличается незначительно. МС- и БС-волокна имеют разные функции во время физической активности. МС-волокнам присущ высокий уровень аэробной выносливости, они эффективны в производстве АТФ на основе окисления углеводов и жиров и более приспособлены к выполнению длительной работы невысокой интенсивности. Быстро сокращающиеся мышечные волокна приспособлены к анаэробной деятельности (без кислорода), и при их работе АТФ образуется благодаря анаэробным реакциям. Б С двигательные единицы производят большую силу, однако легко устают ввиду ограниченной выносливости и используются главным образом при выполнении кратковременной работы высокой интенсивности.
• длинный отросток, отходящий от тела клетки, который тянется на большое расстояние – до 1,5–1,7 м. Он составляет основной, или осевой, отросток нервной клетки. Его называют аксоном (в переводе с латыни axis — ось, основание, основной).
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь.
Формирование мышечной ткани начинается на 4-6-й неделе внутриутробного развития. В это время формируются первичные мышечные волокна. Несколько позже в мышцы прорастают аксоны мотонейронов спинного мозга. С этой стадии начинается синхронное формирование нервно-мышечного аппарата, причем определяющее значение имеет развитие нервных элементов, происходящее на 6-7-м месяцах внутриутробного развития. К моменту рождения примерно половина мышечных волокон уже прошла стадию первичной дифференцировки, и уже определены как «белые» или «красные». Дифференцировочные процессы усиливаются в возрасте от 1 до 2 лет, а затем на стадии полового созревания.
Нейросекреторные клетки, как и обычные нервные клетки, воспринимают сигналы, поступающие к ним от других отделов нервной системы, но далее передают полученную информацию уже гуморальным путем (не по аксонам, а по сосудам) – посредством нейрогормонов. Таким образом, совмещая свойства нервных и эндокринных клеток, нейросекреторные клетки объединяют нервные и эндокринные регуляторные механизмы в единую нейроэндокринную систему. Этим обеспечивается, в частности, способность организма адаптироваться к изменяющимся условиям внешней среды. Объединение нервных эндокринных механизмов регуляции осуществляется на уровне гипоталамуса и гипофиза.
Гидроцефалический (интерстициальный) отек. Еще одной формой отека головного мозга, сопровождающейся увеличением объема интерстициального пространства, является гидроцефалический отек, обусловленный блокадой путей, соединяющих интерстициальное пространство головного мозга с макроскопическими ликворосодержащими пространствами. Для клинициста эта форма отека имеет практическое значение. Так, у больных с острой гидроцефалией в начале происходит увеличение объема интерстициальной жидкости в перивентрикулярных отделах. Узкие в норме пространства между глиальными клетками и аксонами расширяются. Астроциты набухают, атрофируются и погибают. У больных с хронической гидроцефалией деструкция аксонов, разрушение миелина, фагоцитоз липидов микроглии являются характерными гистологическими признаками. Кроме стаза внеклеточной жидкости, причиной отека у этих больных может быть и обратный ток спинномозговой жидкости из желудочков мозга. Так же, как и при любой иной форме отека мозга, в зоне отек; снижается регионарный мозговой кровоток. По-видимому, часть функциональных расстройств в ЦНС, наблюдаемых у больных с гидроцефалией, обусловлена снижением регионарного кровотока в зоне отека.
Эти два гормона относятся к особой группе, поскольку, синтезируясь в гипоталамусе, транспортируются по аксонам (отросткам нейронов) в задний гипофиз и там выделяются в системный кровоток.
Между концами может остаться небольшой диастаз, но он не должен превышать 1 мм. Свободный промежуток между концами нерва заполнит гематома, а в дальнейшем образуется соединительно—тканная прослойка, через эту гематому и соединительную ткань будут прорастать тяжи швашювских клеток и вновь сформированные аксоны.
Аксон – это волокнистая ось, отходящая от тела нейрона, покрытая миелиновым слоем, обеспечивающая связь с другими нейронами и клетками рабочих органов. Представляет собой удлиненный осевой отросток, по которому передаются потенциалы действия (возбуждения), что делает его важнейшим структурным элементом ЦНС.
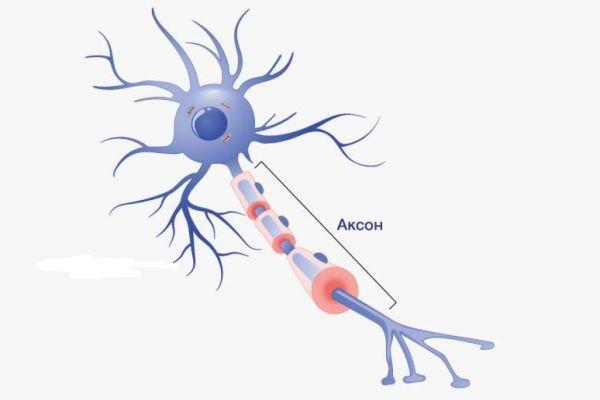
Определение
Мозговое вещество – высокоорганизованная структура, образованная нервными клетками, от которых отходят аксоны. Из нервных клеток состоит мозговая ткань. Аксон в переводе с греческого означает «ось» – это такой отросток, элемент мозгового вещества, который обеспечивает взаимодействие между клетками разного типа (нейроны, клетки иннервируемых органов), что ассоциируется с тонким, четким управлением работой органов и систем. Функции ткани ЦНС:
- Воспринимает раздражения, преобразуя их в импульсы.
- Поддерживает передачу импульсов от управляющих отделов мозга к исполнительным органам.
- Формирует ответную реакцию на раздражающее воздействие.
- Обеспечивает взаимодействие в работе систем и органов, поддерживает интеграцию структурных единиц организма.
- Обеспечивает взаимосвязь организма с внешней средой.
Согласно определению в биологии, аксон (англ. axon) – удлиненный отросток, по которому идут импульсы от тела нейрона к другим нервным клеткам и структурным элементам всех тканей организма. Мозговая ткань в период внутриутробного развития образуется из нервной пластины. Края пластинки прогибаются, что приводит к формированию валиков и желобка. В результате смыкания краев валиков возникает нервная трубка – основа ЦНС.
Дифференциация клеток, образующих трубку, приводит к появлению нейробластов и спонгиобластов. Первые служат основой для формирования нейронов, вторые – для образования нейроглии. Нейроны (анат.) – основные структурные элементы мозгового вещества. Они характеризуются отсутствием функции деления, что приводит к постепенному уменьшению их численности. Тело нейрона состоит из ядра и цитоплазмы. В зависимости от типа нейронов меняется геометрическая форма тела, которая бывает круглая, овальная, пирамидальная и другая.
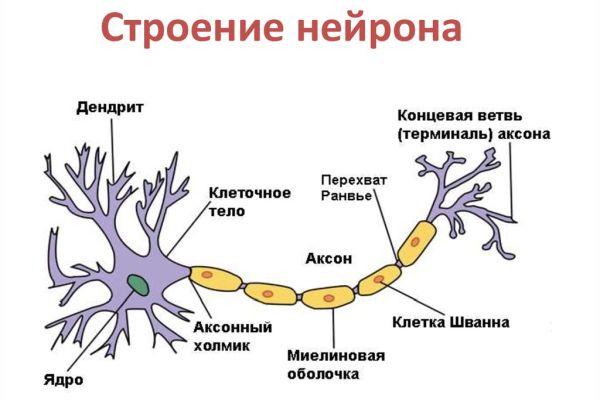
Цитоскелет, состоящий из микротрубочек и нейрофибриллов, обеспечивает опорную и трофическую функцию. Цитоскелет поддерживает форму нейрона, обеспечивает транспорт веществ и органелл. От тела ответвляются отростки – единичный аксон и множественные дендриты. Аксон нейрона почти не ветвится, иногда образует коллатеральные (обходные) сегменты. Концевые сегменты (окончания) разветвляются, называются терминали.
Терминали взаимосвязаны с окончаниями других нейронов и с клетками, образующими паренхиму (ткань) рабочих органов – мышц, желез. Количество дендритов варьируется от 1 до нескольких. Тонкие ответвления дендритов оканчиваются небольшими шипами, где сосредоточены терминали аксональных отростков многих тысяч других клеток. Дендриты воспринимают раздражения или потенциалы действия от других клеток и передают их по волокнам к телу своего нейрона.
Рост аксона зависит от особенностей строения и жизнедеятельности нейрона, который поддерживает функцию питания отростка. К примеру, если перерезать аксональный ствол, сегмент, связанный с телом, остается жизнеспособным и продолжает деятельность, участок, утративший связь с телом, отмирает. Аксоны образуют нервы, что предполагает сложную структурно-морфологическую организацию ЦНС.
Строение
Аксон – это длинный отросток нейрона, который обеспечивает взаимодействие между нервными клетками. Согласно анатомии, аксон ответвляется от холмика, находящегося на теле. Холмик аксона представляет собой структуру, где постсинаптический потенциал преобразуется в биоэлектрический сигнал. Чтобы в холмике происходила генерация биоэлектрических сигналов, необходима согласованная деятельность каналов –натриевых, кальциевых, нескольких типов калиевых.
Длина аксона у человека существенно варьируется в зависимости от вида нейрона, от которого отходит аксональная ось. Минимальная длина – около 1 миллиметра, максимальная – около 1,5 метров. Длина более 1 метра наблюдается в случаях, когда отросток отходит от спинного мозга в область конечностей. Диаметр аксональной оси также неодинаковый у разных типов клеток, равен около 1-20 микрон. Импульсы проходят быстрее по аксональным осям большего диаметра.
Размеры аксонального отростка нередко достигают 99% от общего объема нервной клетки, в структуру которой он входит. Аксон состоит из протоплазмы (аксоплазмы), где находятся тончайшие волокна, белковые нити – нейрофибриллы, из чего образован ствол аксонального ответвления. Согласно одной из теорий, нейрофибриллы – проводники питательных веществ. Аксональная протоплазма также содержит митохондрии и микротрубочки, которые представляют собой самые крупные элементы цитоскелета.
Диаметр микротрубочек составляет около 24 нанометров. Они обеспечивают внутриклеточный транспорт веществ, в том числе поддерживают трофику аксональных отростков. Тело (перикарион) – источник протеинов и нейромедиаторов, распространяющихся по аксональной оси посредством микротрубочек, которые у аксона имеют направленную полярную ориентацию (в отличие от микротрубочек дендритов).
Положительно заряженные концы микротрубочек направлены к сегменту терминали, отрицательно заряженные концы – к телу. Строение аксона предполагает наличие оболочки. Аксон покрыт глиальным (миелиновым) слоем по всей длине, чем защищен от разрушающих внешних воздействий. Миелиновый слой в аксональных отростках периферического отдела сформирован клетками Шванна.
Миелиновая оболочка, покрывающая нервную ось, обеспечивает ее механическую прочность, электрохимическую изоляцию, трофику (питание). Миелиновый слой ускоряет проведение биоэлектрических сигналов. Нервы – пучки объединенных аксональных отростков, которые покрыты оболочкой из соединительной ткани и снабжены кровеносными сосудами.
Функции
Основная задача нейронов – переработка данных. С их помощью осуществляется получение, обработка, передача информации отделам нервной и других систем организма.
Если дендриты проводят сигналы по направлению к телу нервной клетки (перикариону), то аксональный отросток передает импульсы от перикариона к другим клеткам.
Основная функция аксонов – проведение импульсов в пределах нейрональной сети и к исполнительным органам. Аксональные ответвления относятся к первичным проводниковым путям в нервной системе. Вспомогательная функция – транспорт веществ. При помощи аксонального транспорта осуществляется движение белков, синтезированных в теле, нейромедиаторов, органелл. Многие вещества способны двигаться в обоих направлениях.

В периферических сегментах аксона в него могут проникать вирусы и токсичные вещества, которые перемещаясь к телу нервной клетки, повреждают ее. Аксональный транспорт зависит от количества энергии АТФ. Если энергетический уровень АТФ понижается больше, чем в 2 раза, происходит блокировка аксонального транспорта.
Функции аксона заключаются в передаче импульсов. При взаимодействии аксона с телом другого нейрона образуется аксосоматический контакт. Если аксон взаимодействует с дендритами других клеток возникает аксодендритический контакт. Взаимодействие с аксоном другой клетки приводит к образованию аксо-аксонального контакта, который редко происходит в нервной системе, поддерживает тормозные рефлекторные реакции.
Особенности регенерации нервной ткани
Нервные клетки почти полностью лишены способности к регенерации. Однако нервные клетки способны восстанавливать поврежденные или утраченные ответвления. Процесс регенерации аксона возможен, если тело сохраняет жизнеспособность, и на пути роста аксонального отростка отсутствуют препятствия. В ходе процесса регенерации отросток вновь прорастает к органу-мишени.
Восстановление нервной проводимости в мышцах с нарушенной иннервацией – один из критериев успешного лечения невропатий разного генеза. При невропатиях травматического генеза восстановление функций мышц происходит за счет регенерации ствола прерванного аксона и ремиелинизации отростка (восстановление миелиновой оболочки). Периферический отдел нервной системы обладает более высоким потенциалом регенерационных возможностей в сравнении с центральным отделом.
Восстановление иннервации в мышечной или кожной ткани происходит благодаря сохранившимся аксонам, которые начинают ускоренно разрастаться и ветвиться. Процесс ветвления аксонов в зоне перехватов Ранвье (периодические разрывы миелинового слоя) получил название «спрутинг». В результате происходит частичное или полное возобновление первичной иннервации.
В ходе экспериментов установлено, что близлежащие интактные (не вовлеченные в патологический процесс) аксоны выпускают нервные волокна, которые иннервируют участок мускулатуры или кожных покровов с нарушенной проводимостью нервных импульсов. Различают виды спрутинга – коллатеральный (обходной) и регенераторный (терминальный).
Регенераторный спрутинг начинается после устранения в нейронах ретроградных изменений, обусловленных аксонотомией (повреждением, рассечением нервной оси). Это связано с потребностью в продукции аппарата ядра нервной клетки, производящего протеины. Материал, необходимый для регенераторного спрутинга, продуцируется в теле и транспортируется по микротрубочкам по всей длине оси. Параллельно происходит процесс ремиелинизации осевого ствола.
Аксон – удлиненный отросток нервной клетки, обеспечивающий взаимодействие между структурными элементами мозговой ткани и связь ЦНС с исполнительными органами.
Просмотров: 2 336
Нервные импульсы распространяются при перемещении ионов через мембрану нервной клетки и передаются из одной нервной клетки в другую с помощью нейромедиаторов.
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
См. также: