Запрос «Губка» перенаправляется сюда; см. также другие значения.
| Губки | |||||||||
 |
|||||||||
| Научная классификация | |||||||||
|---|---|---|---|---|---|---|---|---|---|
|
|||||||||
| Международное научное название | |||||||||
|
Porifera Grant in Todd, 1836 |
|||||||||
| Синонимы | |||||||||
|
|||||||||
| Классы[1] | |||||||||
|
|||||||||
| Геохронология | |||||||||
|
Гу́бки (лат. Porifera — «носитель пор») — парафилетическая группа организмов; многоклеточные водные, преимущественно морские, модульные беспозвоночные животные, не достигшие уровня тканевой организации. Часто образуют колонии. В настоящее время описано около 8000 видов[2]. Губки известны с докембрия[3]. Зародышевые листки имеют обратное расположение — дважды впячивание полюсов бластулы внутрь; тело губки образовано тремя основными группами клеток: пинакодермой — покровным слоем, хоанодермой — выстилкой жгутиковых камер и мезохилом — межклеточным веществом со скелетными элементами. Мышечная и нервная системы отсутствуют. Отличительной чертой губок является наличие водоносной системы, с помощью которой осуществляется дыхание, питание, выделение и размножение. Губки — фильтраторы. Размножение бесполое и половое. У большинства — развитие с метаморфозом: образуются типичные личинки — паренхимула и амфибластула.
Содержание
- 1 Общая схема строения
- 1.1 Дермальный слой
- 1.2 Гастральный слой
- 1.3 Мезохил
- 2 Водоносная система
- 2.1 Аскон
- 2.2 Сикон
- 2.3 Лейкон
- 3 Фильтрация
- 4 Локомоция и перемещение клеток по организму
- 5 Питание
- 6 Внутренний транспорт, газообмен и выделение
- 7 Интеграция
- 8 Биологически активные метаболиты и ассоциации с другими организмами
- 9 Биоэрозия
- 10 Размножение
- 10.1 Бесполое размножение
- 10.2 Половое размножение и развитие
- 11 Палеонтология и филогения
- 12 Влияние на культуру
- 13 Примечания
- 14 Литература
- 15 Ссылки
Общая схема строения
В самом простом случае губка имеет бокаловидную форму и обладает гетерополярной осевой симметрией. К субстрату губка крепится подошвой; на верхнем полюсе расположено устье — оскулюм, через которое из тела губки выводится вода. У колониальных осевая симметрия нарушается и возникает множество оскулюмов. Стенки губки пронизаны многочисленными порами, ведущими в парагастральную полость. Тело состоит из двух слоёв клеток: наружного — дермального и внутреннего, выстилающего внутреннюю полость — гастрального. Между ними формируется мезохил — бесструктурное вещество с отдельными разбросанными в нём клетками.
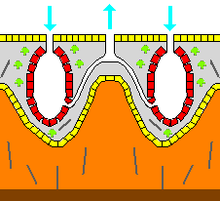
![]()
Гистология известковой губки
— пинакоциты
— хоаноциты
— археоциты и другие типы клеток в мезохиле
— мезохил
— спикулы
— карбонатные элементы
— субстрат
— направление тока воды
Дермальный слой
Дермальный слой губок — пинакодерма — имеет вид погружённого эпителия и образован тонким (1 мкм) слоем пинакоцитов. Пинакодерма выстилает наружную стенку тела всех губок, а также стенки крупных каналов известковых и обыкновенных губок. В зависимости от месторасположения пинакоциты делятся на несколько групп:
- экзопинакоциты — на наружной поверхности тела:
- базипинакоциты — на подошве, прикрепляют губку к субстрату;
- пороциты — перфорированные экзопинакоциты, способны к сокращению;
- эндопинакоциты — на выстилке каналов:
- просэндопинакоциты — приводящих;
- апэндопинакоциты — отводящих.
От настоящего эпителия пинакодерма большинства губок отличается отсутствием десмосом и плотных контактов.
Гастральный слой
Гастральный слой тела губок — хоанодерма — выстилает жгутиковые камеры или парагастральную полость и состоит из хоаноцитов — воротничковых клеток цилиндрической формы. Из центра обращённого в парагастральную полость конца хоаноцитов выходит длинный жгутик, окружённый плазматическим воротничком. Хоаноциты делятся на 2 типа:
- центральные клетки — регулируют ток воды внутри жгутиковых камер;
- апопильные клетки — расположены у входа в жгутиковые камеры, есть не у всех групп.

![]()
Клеточное строение губки асконоидного типа
| — хоаноцит — пинакоцит — лофоцит — ооцит — археоцит |
— склероцит — пороцит — мезохил — спикулы — ток воды |
Мезохил
Мезоглея — бесструктурное вещество, заполняющее пространство между жгутиковыми камерами. Имеет тонковолокнистую структуру и состоит из различных типов клеток:
- археоциты — недифференцированные резервные подвижные клетки, способные превращаться во все типы клеток, в том числе половые. Имеют крупное ядро с ядрышком, а также хорошо развитые эндоплазматическую сеть и аппарат Гольджи.
- трофоциты — содержат запас питательных веществ для размножения;
- тезоциты — клетки геммул, наполненные желтком;
- клетки опорного скелета:
- спонгиобласты — тонкие коллагеновые фибриллы;
- лофоциты — толстые коллагеновые фибриллы;
- склеробласты — клетки, образующие скелет губок;
- сократимые клетки:
- колленциты (миоциты) — клетки, способствующие сокращению мезохила;
- нейроидные клетки — возбудимые клетки;
- клетки со включениями — функция не ясна; вероятно, задействованы в запасании веществ, секреции гормонов и пищеварении.
В состав неклеточного компонента мезохила входят органические молекулы (гликопротеины), свободный коллаген, пищевые частицы и продукты секреционной деятельности клеток.
Водоносная система
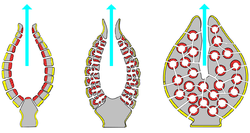
![]()
Основные типы строения водоносной системы губок: аскон, сикон и лейкон (слева направо)
![]()
Строение аскона
1 — оскулюм
2 — спонгиоцель
3 — хоанодерма
4 — остиум
5 — пинакодерма
![]()
Строение сикона
| 1 — оскулюм 2 — спонгиоцель 3 — приводящий канал 4 — отводящий канал 5 — хоанодерма |
6 — прозопиль 7 — остиум 8 — хоанодерма 9 — мезохил 10 — апопиль |
Аскон
У ряда Известковых губок произошли увеличение размера жгутиковых камер и редукция системы приводящих и отводящих каналов. Такие губки состоят из трубки или разветвлённой системы трубок, изнутри выстланной хоаноцитами; при этом поры (остии) открываются непосредственно в жгутиковые камеры. Водоносная система характерна только для двух родов — Leucosolenia и Clathrina.
Сикон
Формирование сикона вызвано разрастанием мезохила и впячиванием в него участков парагастральной полости, образующих радиальные трубки. Хоаноциты выстилают только жгутиковые трубки и исчезают с других участков атриальной полости. Стенки тела губки утолщаются, и между поверхностью тела и жгутиковыми трубками образуются приводящие каналы. Строение сиконального типа характерно лишь для немногих видов (например, для Scypha и Grantia).
Лейкон
Наиболее сложный тип строения. Это колониальные губки с многочисленными оскулюмами. В мощном слое мезоглей много скелетных элементов. Стенка тела пронизана сетью каналов, связывающих многочисленные жгутиковые камеры.
Фильтрация
В среднем каждые 5 секунд губка прокачивает количество воды, равное объёму своего тела. Поскольку вода несжимаема, её объём на входе должен быть в каждый момент равен объёму на выходе. С наибольшей скоростью вода течёт в области оскулюма, а с наименьшей — через жгутиковые камеры, так как эти два участка водоносной системы губки имеют соответственно наименьшую и наибольшую общую площадь сечения. Многие губки при необходимости могут снижать общую скорость движения воды или полностью останавливать поток, чтобы избежать попадания ила внутрь организма. Это достигается за счёт регуляции диаметра оскулюма с помощью сократимых миоцитов, закрывания остий (иногда посредством трубковидных пороцитов) или же регуляции биения жгутиков хоаноцитов. Последнее характерно, например, для стеклянных губок, у которых нет ни миоцитов, ни пороцитов.
Ток воды создаётся биением жгутиков хоаноцитов. Каждый жгутик совершает ундулирующие движения в одной плоскости. Расположенные на нижнем участке жгутика в области воротничка лопасти из мастигонем, возможно, помогают «выкачивать» из него воду. По меньшей мере у одного вида (Trochospongilla pennsylvanicus) плоскость биения жгутика хоаноцитов каждые несколько секунд немного смещается, так что в конечном счёте она поворачивается на 360°. Жгутики и воротнички хоаноцитов направлены в противоположную от остий сторону у асконоидных губок или от прозопилей — у сиконоидных и лейконоидных губок. Волна проходит по жгутику от его основания к вершине, создавая ток воды в сторону отводящих каналов и оскулюма.
У многих губок оскулюмы расположены на вершине специальных трубчатых выростов, сильно возвышающихся над основной массой тела, поверхность которого несёт остии. Скорость течения омывающей губку воды в зоне, где находятся оскулюмы, обычно выше, чем в придонном слое, у основания губки. Интенсивное движение воды над трубчатыми выростами создаёт область пониженного давления непосредственно в оскулюмах по сравнению с давлением в остиях. В результате возникает ток воды из области высокого давления в область низкого давления, то есть от остий к оскулюмам. Многие губки обитают в местах, где течения весьма значительны, поэтому дополнительно возникающий градиент давления в водоносной системе усиливает ток, создаваемый работой жгутиков. В свою очередь это позволяет губкам экономить энергию.
Локомоция и перемещение клеток по организму
Хотя губки в своём абсолютном большинстве — сидячие животные, ведущие прикреплённый образ жизни, некоторые виды способны к ограниченной локомоции. И пресноводные (Ephydatia), и морские (Chondrilla, Hymeniacidon, Tethya) губки могут перемещаться по субстрату со скоростью от 1 до 4 мм в день. Это достигается в результате амебоидного движения пинакоцитов и клеток других типов, которое осуществляется одновременно и более или менее согласованно. Губки могут также сокращаться (Clathrina coriacea) и изменять с помощью миоцитов диаметр оскулюма. Благодаря этим проявлениям сократимости останавливается или ограничивается поток воды через водоносную систему в ответ на неблагоприятные воздействия окружающей среды, например при внезапном увеличении количества взвеси ила в воде.
Отличительный признак губок — способность значительной части клеток, образующих тело, свободно перемещаться. Клетки мезохила (все они амёбоидные) передвигаются практически постоянно. Эндопинакоциты и хоаноциты тоже могут менять свою локализацию в процессе перестройки водоносной системы. Реорганизация последней проявляется в образовании новых и объединении уже существующих жгутиковых камер, слиянии и образовании новых ветвей каналов. В результате ток воды оптимизируется по мере роста губки или перемен в окружающей среде. Такие независимые и частые перемещения клеток возможны потому, что в тканях губок отсутствуют клеточные контакты.
Питание
Губки отфильтровывают пищевые частицы из воды, протекающей через их тело. Размеры частиц обычно варьируют от 50 до 1 мкм, но могут быть и ещё меньше. В эти пределы попадают одноклеточные планктонные организмы, такие, как динофлагелляты и бактерии, вирусы, мелкий органический детрит и, возможно, даже растворённые в воде органические вещества. В тропических морях, где губки очень многочисленны, мелкие фракции пищи примерно в 7 раз более доступны, чем крупные. Все клетки губок способны поглощать частицы путём фагоцитоза.
Фильтры, улавливающие пищевые частицы, — это приводящие каналы, постепенно уменьшающиеся в диаметре по мере погружения вглубь тела, и хоаноциты. Пищевые и иные частицы в зависимости от их диаметра отфильтровываются разными участками системы. Наиболее крупные частицы, диаметр которых превышает примерно 50 мкм, не проходят через остии, но их могут фагоцитировать клетки экзопинакодермы. Частицы в пределах от 5 до 50 осаждаются в приводящих каналах, их поглощают также путём фагоцитоза эндопинакоциты или археоциты. Последние проникают в просвет каналов через их выстилку между пинакоцитами или через пороциты. Мелкие частицы размерного класса бактерий поступают в жгутиковые камеры, где хоаноциты поглощают их путём фагоцитоза или пиноцитоза. Самые мелкие частицы могут улавливаться «ситом», образуемым микроворсинками воротничков хоаноцитов и окружающим их внеклеточным матриксом.
И хоаноциты, и археоциты поглощают и переваривают пищу в пищеварительных вакуолях, однако часто хоаноциты только захватывают частицы, а затем «передают» их археоцитам, в которых и осуществляется процесс пищеварения. Археоциты, по-видимому, также запасают питательные вещества (гликоген и липиды).
К семейству Cladorhizidae (Demospongiae) относятся хищные губки. Они ловят ракообразных и других мелких животных, которые прилипают к раскинутым губкой клейким, состоящим из клеток нитям. Когда добыча попалась, нить укорачивается и подтягивает жертву к поверхности тела. Губка медленно обволакивает пойманное животное и переваривает его, вероятно, с помощью археоцитов. У этих необычных губок нет ни хоаноцитов, ни водоносной системы.
Два основных типа частиц, подлежащих удалению из тела губки, — это непереваренные в процессе внутриклеточного пищеварения остатки пищи и неорганические частицы, попавшие в губку с током воды. Минеральные частицы необходимо удалять из приводящих каналов, иначе они закупорят их и нарушат их деятельность. Застрявшую в приводящем канале частицу фагоцитирует археоцит, который переносит её ближе к выходу из водоносной системы и путём экзоцитоза выводит в отводящий канал. У губок, использующих инородный материал для постройки скелета, археоцит может транспортировать захваченную частицу к месту сборки скелетных элементов.
Многие губки, как морские, так и пресноводные, содержат в тканях фотосинтезирующих эндосимбионтов и используют продукты фотосинтеза для питания. В археоцитах и других клетках пресноводных губок обычно поселяются зелёные водоросли (зоохлореллы). Морские губки (известковые и обыкновенные) могут выступать в роли хозяев динофлагеллят (зооксантелл) или, что бывает чаще, цианобактерий. В спонгиновых волокнах скелета губки Mycale laxissima из Белиза поселяются как зелёные, так и красные водоросли. В некоторых случаях симбиотические цианобактерии составляют до одной трети массы губки (например, у Verongia). Такие губки обитают на мелководных, хорошо освещённых участках дна, и их симбиотические бактерии часто локализуются в поверхностных участках тела. Губка использует избыточные продукты фотосинтеза в виде глицерола и фосфорилированных соединений. Некоторые губки с Большого Барьерного рифа за счёт симбиотических цианобактерий покрывают от 48 до 80 % своих энергетических потребностей. Помимо цианобактерий и других мутуалистических симбионтов в губках часто встречаются внутри- и внеклеточные бактерии, роль которых неясна.
Внутренний транспорт, газообмен и выделение
Расстояние между каналами водоносной системы, с помощью которой вентилируется тело губки, и любыми входящими в его состав клетками не превышает 1 мм. Соответственно поступление газов и продуктов обмена (в основном, аммиака) из тела в воду, протекающую по водоносной системе, обеспечивается простой диффузией. Возможно, и питательные вещества из многочисленных участков, где осуществляется внутриклеточное пищеварение, также распространяются по телу губки за счёт диффузии, хотя за их доставку к развивающимся гаметам и тканям ответственны способные к амёбоидному движению археоциты. По крайней мере у одного вида Aplysina (Verongia) есть специализированные внутренние волокна, служащие своего рода «рельсами», по которым движутся нагруженные питательными веществами археоциты.
Транспорт питательных веществ у стеклянных губок осуществляется в пределах синцития. После захвата пищевых частиц воротничковыми телами вакуоли с частицами транспортируются по всему синцитию губки вдоль пучков микротрубочек динеиновыми молекулярными моторами. Этот способ транспортировки идентичен транспорту везикул в псевдоподиях фораминифер и в аксонах нервных клеток высших животных.
Практически полное отсутствие межклеточных контактов в пинакодерме и хоанодерме заставляет предполагать, что эти слои образуют довольно ненадёжный регуляторный барьер между мезохилом и водой, омывающей губку снаружи. Вода извне свободно поступает в их тело. Соответственно интерстициальная по своему составу, то есть находящаяся между клетками губки, жидкость должна мало отличаться от воды, в которой губки живут. Это относится и к пресноводным видам. В клетках пресноводных губок, как правило, имеются сократительные вакуоли, но с их помощью осморегуляция осуществляется только в каждой отдельной клетке, а не во всём теле животного.
Интеграция
У губок нет ни нервных клеток, ни нервной ткани, хотя у некоторых видов в ограниченной мере проявляется способность к передаче импульсов. Как правило, эта проводимость представляет собой медленное, «эпителиальное» распространение электрического сигнала на расстояние нескольких миллиметров и приводит к местному сокращению миоцитов в ответ на локальный стимул. Малая скорость передачи сигнала связана с отсутствием специализированных межклеточных контактов (щелевых контактов), улучшающих проводимость эпителиев. Поэтому клеточные мембраны скорее играют роль изолирующих барьеров, нежели обеспечивают прохождение «волны» деполяризации. Стеклянные губки представляют собой исключение из этого общего правила. У Rhabdocalyptus dawsoni электрические импульсы (потенциалы действия) быстро распространяются по синцитиальным тяжам от точки стимуляции по всему телу губки. В результате биение жгутиков прекращается и ток воды через губку останавливается.
Биологически активные метаболиты и ассоциации с другими организмами
Очень часто продукты обмена губок препятствуют поселению других организмов на их поверхности или отпугивают хищников. Было показано, что 9 из 16 антарктических губок и 27 из 36 карибских губок токсичны для рыб. Однако далеко не всегда пагубные для рыб токсины отпугивают и других животных, да и среди рыб встречаются специализированные «губкоядные» виды, например в семействах Pomacanthidae (рыбы-ангелы) и Monacanthidae (единороговые), а также мавританский идол (Zanclus cornutus). Морские черепахи, особенно бисса (Eretmochelys), охотно питаются губками; кремнезёмные спикулы губок могут составлять до 95 % их фекалий. Представители одной из групп голожаберных моллюсков (Doridacea) демонстрируют узкую специализацию, питаясь лишь определёнными видами губок, почти как гусеницы ряда бабочек, которые тесно связаны с конкретными кормовыми растениями. Некоторые губки используют метаболиты в конкурентной борьбе с другими организмами за пространство. Например, карибская губка Chondrilla nucula выделяет вещества, которые убивают находящиеся поблизости кораллы, а губка после этого обрастает их прочные скелеты. Многие губки обладают сильным запахом, например «чесночная» губка Lissodendoryx isodictyalis. Встречаются среди губок и такие, прикосновение к которым вызывает сильное раздражение кожи (например, карибская «огненная» губка Tedania ignis). Выделяемые губками биохимические соединения активно изучаются для определения их медицинского и промышленного значения.
Многие губки становятся убежищами для эндосимбионтов — более мелких организмов, которые живут в их водоносной системе как в многоквартирном доме и используют токи воды и защиту, предоставляемую хозяином. Крупные лейкопоидные «дома» заселены мелкими ракообразными и офиурами. Один исследователь извлёк более 16000 раков-щелкунов (Alpheidae) из каналов водоносной системы одной-единственной крупной бочонковидной губки (Spheciospongia). Некоторые многощетинковые черви из семейства Spionidae поселяются в губках, питаются ими и благодаря этому приобретают цвет приютившей их губки. Острорылый краб (Oregonia gracilis) прикрепляет губок, водоросли и некоторых сидячих животных к своему панцирю, так что на нём образуется своего рода микросообщество. Разрастаясь на этом подвижном субстрате, организмы, формирующие сообщество, надёжно маскируют краба. Другие крабы (Dromiidae) отрезают и прикрепляют к себе на спину, как покрышку, верхнюю часть губки или используют для этой цели лишь её небольшой фрагмент, которым постепенно обрастает весь панцирь, обеспечивая крабу защиту.
Биоэрозия
Губки из семейства Clionidae (Demospongiae) играют важную роль в разрушении известковых раковин и коралловых рифов в морях. Cliona celata, например, сверлит раковины моллюсков, проделывая в их толще сеть туннелей. Периодически в образовавшиеся отверстия губка выдвигает небольшие участки тела, имеющие вид сосочков (папиллы). Одни папиллы несут остии, другие — оскулюмы. В конце концов губка полностью разрушает всю раковину.
«Сверление» осуществляют специализированные археоциты, которые носят название «травильных» клеток. Каждая такая клетка воздействует на раковину химическими веществами, после чего отделяет от неё маленький кусочек. Образовавшийся «осколок» транспортируется в водоносную систему и выводится наружу через оскулюм. Скрытая внутри раковины или скалы губка надёжно защищена от хищников, поэтому такой способ существования может повышать вероятность выживания как молодых, так и взрослых губок.
Размножение
Бесполое размножение
Бесполое размножение губок осуществляется путём фрагментации, почкования или образования зимующих пропагул (геммул). Фрагментация, как правило, происходит в результате повреждения губки течениями, прибоем или хищниками. Оторвавшиеся куски выживают благодаря способности губки к регенерации. Фрагмент прикрепляется к субстрату и, перестраиваясь, превращается в нормально функционирующую губку. Крайнее проявление фрагментации — диссоциация губки на отдельные клетки или группы клеток. Это может быть достигнуто путём протирания кусочка губки через мелкоячеистую ткань. Подобный эксперимент был впервые поставлен зоологом Г. В. Уилсоном в начале XX в. и с тех пор часто воспроизводился для изучения механизмов клеточного распознавания и моделирования морфогенеза.
Почкование у губок встречается относительно редко, но всё же имеет место у небольшого числа видов. Например, есть данные, что у Clathrina, имеющей асконоидный тип строения, свободные концы трубок вздуваются и образуют почки, которые затем отделяются, прикрепляются к субстрату и дают начало новой губке. У некоторых видов Tethya почки образуются на особых стебельках. Губки, относящиеся к родам Oscarella и Aplysilla, по некоторым данным, формируют папиллы, затем папиллы отделяются и дают начало новым организмам.
Многие пресноводные и некоторые морские губки образуют сотни, а то и тысячи напоминающих споры геммул. Как правило, это происходит осенью. Осенние геммулы пресноводных видов могут входить в состояние диапаузы, при котором практически полностью подавляется их метаболическая активность. Для активизации таких геммул необходимо, чтобы они в течение определённого периода подвергались воздействию очень низкой температуры. Затем геммулы прорастают и дают начало новым губкам. Обычно это происходит весной. Пока геммула находится в диапаузе, она устойчива к экстремальным воздействиям окружающей среды, таким, как очень низкая температура, повышенная солёность и высыхание.
Геммулы образуются в мезохиле погибающей губки вокруг группы богатых питательными веществами археоцитов. Спонгоциты формируют спонгиновую оболочку вокруг этой клеточной массы. Оболочка может быть инкрустирована спикулами, которые образуются в склероцитах. Она полностью покрывает клеточную массу; лишь на одном полюсе остаётся небольшое отверстие — микропиле. Сформированная геммула состоит из оболочки и находящихся внутри археоцитов, каждый из которых вскоре округляется и становится похож на эмбриональную клетку. Такие модифицированные археоциты называются тезоцитами.
Весной геммулы «прорастают»: периферические тезоциты дифференцируются в пинакодерму, которая, быстро разрастаясь, выступает наружу через микропиле, как надувающийся пузырь. Этот «пузырь» пинакодермы соприкасается с субстратом и прикрепляется к нему. Затем через микропиле внутрь пузыря перемещаются остальные тезоциты, которые составляли центральную часть геммулы. После дифференциации они образуют внутренние структуры молодой губки.
Наблюдения за развитием молодых губок из геммул позволили получить данные, ставящие под сомнение возможность использования по отношению к губкам понятия «индивидуальность». В процессе прорастания тезоциты из разных геммул одной и той же губки или из геммул, принадлежащих разным экземплярам, но обязательно одного вида, могут объединяться. В этом случае из них развивается одна новая губка, обладающая своей «индивидуальностью».
Половое размножение и развитие
Губки, за редким исключением, — гермафродиты. Когда приходит время, одна губка выбрасывает наружу сперматозоиды, которые переносятся токами воды к другой губке, внутри которой происходит оплодотворение яиц. У яйцекладущих видов (например, Cliona) зиготы выводятся во внешнюю среду, где и протекает их развитие. Многим губкам свойственно живорождение: зиготы остаются внутри материнского организма, который покидают уже сформированные личинки (иногда такие формы называют «личинкородящими»). Зародыши и личинки губок лецитотрофны.
Считается, что у губок нет половых органов (гонад). Половые клетки залегают в мезохиле либо небольшими группами (сперматозоиды), либо поодиночке (яйца). Сперматозоиды образуются из отдельных хоаноцитов или целых жгутиковых камер, которые погружаются в мезохил и окружаются тонкой клеточной стенкой, давая начало сперматоцисте. Яйца образуются из археоцитов или, как у некоторых известковых губок, — из дедифференцированных хоаноцитов. Обычно яйцо накапливает желток, фагоцитируя окружающие их питающие клетки, или клетки-«няньки». Яйцо и клетки-«няньки» иногда заключены в особый фолликул, имеющий клеточные стенки. (По определению, орган состоит из двух и более тканей. Если будет доказано, что клетки стенки цисты или фолликула и половые клетки имеют разное происхождение и формируются из разных тканей, то можно будет сперматоцисты и яйцевые фолликулы считать органами — гонадами.) Поскольку водоносная система одинаково снабжает все части тела, половые клетки довольно равномерно распределены по мезохилу, но при этом всегда находятся недалеко от канала или камеры, в пределах расстояния, на котором действует диффузия.
В период размножения стенка сперматоцисты разрушается, сперматозоиды попадают в отводящие каналы (или в атриум) и выбрасываются наружу через оскулюмы. Некоторые тропические виды внезапно извергают из оскулюмов молочные облачка сперматозоидов. Возможно, одновременное высвобождение большого количества сперматозоидов характерно для большинства губок.
Когда выброшенные сперматозоиды оказываются рядом с другой губкой, токи воды заносят их в её водоносную систему. Внутри губки сперматозоиды доставляются к поверхности хоанодермы или в жгутиковые камеры, где их фагоцитируют (но не переваривают) хоаноциты. Хоаноцит, захвативший сперматозоид, теряет жгутик и воротничок и трансформируется в амебоидную клетку, которая называется клеткой-носительницей. Она переносит головку сперматозоида (яйцо) к яйцу. Когда клетка-носительница достигает ближайшего расположенного в мезохиле яйца, она либо «передаёт» ему ядро спермия, либо яйцо фагоцитирует эту клетку вместе с содержащимся в ней ядром спермия. В любом случае оплодотворение происходит внутри «яичника» губки.
Как правило, сперматозоиды губок лишены акросомы — структуры, которая у большинства других животных отвечает за проникновение спермия через мембрану яйцеклетки при оплодотворении. Вероятно акросома не нужна в тех случаях, когда ядро спермия попадает в яйцо путём фагоцитоза. У губки Oscarella lobularis спермии всё же имеют акросому, что может свидетельствовать о существовании у этого вида обычного способа оплодотворения яйцеклетки.
Зигота претерпевает полное и равномерное дробление. Однако взаимное расположение образующихся бластомеров варьирует у разных видов губок. Разнообразны и развивающиеся из зародыша личинки. Они носят названия целобластула, амфибластула, паренхимула и трихимелла.
Личинка целобластула характерна для известковых губок, например видов рода Clathrina (Calcinea). Целобластула представляет собой полый шар, стенка которого состоит из одного слоя жгутиковых клеток. Пока личинка ведёт планктонный образ жизни, некоторые её клетки теряют жгутики, становятся амебоидными и мигрируют в бластоцель, так что он, в конце концов, полностью заполняется клетками. Полая целобластула превращается в лишённую полости стерробластулу.
Личинка амфибластула встречается у других известковых губок, например Grantia, Sycon и Leucosolenia (Calcaronea). Амфибластула представляет собой полый шар, состоящий из клеток двух типов: жгутиковых клеток на переднем полюсе личинки и безжгутиковых клеток с зернистой цитоплазмой на заднем полюсе. Когда личинка ещё находится в мезохиле материнского организма, все жгутики обращены в бластоцель, однако между «зернистыми» клетками на заднем полюсе личинки вскоре образуется отверстие, через которое она выворачивается наизнанку. После инверсии жгутики оказываются на наружной поверхности личинки, которая таким образом приобретает способность плавать с их помощью. На этой стадии личинка покидает материнский организм. Инверсия имеет место у тех губок, яйца которых образуются из хоаноцитов: после оплодотворения клетки делятся так, словно они должны образовать новые жгутиковые камеры, внутрь которых направлены жгутики. Обыкновенные губки Oscarella и Plakina тоже имеют в развитии личинку типа амфибластулы, но она образуется вторично, уже после стадии паренхимы. В своём развитии они проходят стадию плотной морулы, которая позднее в результате выселения клеток из центральной части в наружный клеточный слой (мультиполярная эмиграция) превращается в личинку особого типа — цинктобластулу, которая по своему строению принципиально отличается от типичной амфибластулы, свойственной известковым губкам.
Личинка паренхимула характерна для большинства обыкновенных губок. В этом случае зародыш развивается непосредственно в стерробластулу — клеточную массу без внутренней полости. Наружный слой в основном состоит из жгутиковых клеток, между которыми изредка попадаются клетки, лишённые жгутиков и содержащие в цитоплазме множество везикул. Внутри личинки находятся дифференцированные клетки многих типов: склероциты, колленциты, пинакоциты, даже жгутиковые камеры, а также археоциты. Таким образом, паренхимулу в какой-то степени можно считать «почти готовой» молодой губкой, специализированной для плавания.
Личинка трихимелла — типичная стадия развития стеклянных губок. Трихимелла представляет собой стерробластулу с пояском жгутиковых клеток по экватору. Внутри она заполнена клетками, содержащими желток, склероцитами (со спикулами), клетками других типов и даже жгутиковыми камерами.
Все личинки губок лецитотрофны и, следовательно, живут относительно недолго. Как правило, выход личинок стимулируется светом и обычно происходит на рассвете. По истечении определённого периода, продолжительность которого варьирует от нескольких часов до нескольких дней, личинки оседают и начинают ползать по дну в поисках походящего места для прикрепления. Обнаружив такое место, личинка прикрепляется, претерпевает метаморфоз и превращается в молодую губку. Её строение несколько отличается у разных губок и определяется типом личинки. Поскольку метаморфоз сопровождается перегруппировкой клеток с образованием более или менее чётко выраженных пластов, его часто сравнивают с гаструляцией у других многоклеточных. Однако и погружение клеток в полость личинки, что имеет место у многих губок при формировании стерробластул, также можно считать своего рода гаструляцией.
Попытки найти гаструляцию у губок сопровождают практически всю историю изучения этих организмов. В своё время даже выдвигалась идея об «извращении положения зародышевых листков» у губок. Считалось, что поверхностно расположенные жгутиковые клетки личинок представляют собой эктодерму, которая позднее погружается внутрь тела и даёт начало хоанодерме, а крупные безжгутиковые клетки — это энтодерма, которая остаётся снаружи и превращается в пинакодерму. Отсутствие у губок жёстко детерминированных тканевых дифференцировок позволяет некоторым исследователям говорить о том, что у этих животных не может быть и настоящей гаструляции, в процессе которой осуществляется становление двух первых зародышевых листков — экто- и энтодермы. Процессы обособления нескольких клеточных совокупностей, сопровождающие эмбриональное развитие губок, чисто внешне могут напоминать разные типы гаструляции настоящих многоклеточных животных, но не гомологичны им, точно так же, как не гомологичны ткани и органы Eumetazoa пинакодерме, хоанодерме и мезохилу губок.
Непосредственно перед метаморфозом целобластулы, которая к этому моменту уже превратилась в стерробластулу, её клетки дифференцируются и образуют сплошную массу тотипотентных клеток. После прикрепления клеточная масса распластывается по субстрату. Клетки, расположенные на её поверхности, дают начало пинакодерме, а залегающие более глубоко дифференцируются и дают начало всем остальным типам клеток, характерным для губок. В процессе перераспределения внутренних клеток щелевидные просветы между ними постепенно объединяются, образуя атриум.
Амфибластула оседает на субстрат и прикрепляется к нему передним, несущим жгутики концом. После прикрепления жгутиковые клетки теряют жгутики, мигрируют внутрь и образуют внутренние части губки. «Зернистые» клетки дают начало пинакодерме. Когда молодой организм начинает активно функционировать и питаться, он представляет собой миниатюрную асконоидную трубку. Эта стадия называется олинтус.
Метаморфоз личинок, относимых к парнехимулам, протекает немного по-разному у разных видов. Обычно после прикрепления личинки её внутренние клетки дифференцируются и перераспределяются. Из них образуется бо́льшая часть тела губки, если не вся она целиком.
У вида Mycale contarenii жгутиковые клетки превращаются в хоаноциты. У других видов (например, у некоторых пресноводных губок, а также у Microciona prolifera) жгутиковые клетки фагоцитируются археоцитами и, соответственно, не входят в число клеток, принимающих участие в формировании тела молодой губки. В любом случае, прежде чем приобрести лейконоидную организацию, молодая, только что претерпевшая метаморфоз губка часто характеризуется аксоноидным или сиконоидным строением, но имеет при этом толстые стенки. Ювенильные губки, находящиеся на этой стадии развития носят название рагон.
В умеренных широтах губки живут от одного года до нескольких лет. Представители многих тропических видов (а также, возможно, глубоководные губки) могут жить очень долго, до 200 лет и более. Некоторые губки начинают размножаться половым путём только по достижении ими возраста в несколько лет, в то время как другие приступают к размножению в возрасте 2 или 3 недель. Некоторые коралловые губки (Sclerospongia) растут очень медленно, давая прирост около 0,2 мм в год. Если эта скорость роста постоянна, то возраст коралловых губок диаметром 1 м может достигать 5000 лет.
Шестилучевая губка Scolymastra joubini живёт до 10 тысяч лет, это максимальная величина в животном мире.
Палеонтология и филогения
Первые свидетельства о существовании Hexactinellida относятся к протерозою. В позднем протерозое обнаружены и остатки обыкновенных губок, в том числе и «роговых». Известковых губок и Homoscleromorpha пока удаётся обнаруживать только начиная с палеозоя (кембрий).
Ископаемые «группы» Sphinctozoa и Stromatoporata в настоящее время не рассматриваются как самостоятельные таксоны. По мнению ряда исследователей, это не более чем своего рода морфологические типы, отражающие разные варианты организации тела губок. Это в полной мере относится и к Sclerospongae. Вымершие Archaeocyatha, которых и сейчас рассматривают как самостоятельный класс, по-видимому, ближе всего стояли к обыкновенным губкам. Представители таксона Archaeocyatha имели пористый известковый скелет в форме перевёрнутого полого конуса с радиальными септами и двойной стенкой. Представители Stromatoporata напоминали современных коралловых губок (Sclerospongiae). Они тоже обладали массивным базальным известковым скелетом с внутренними трубками, но в отличие от Sclerospongiae у них не было кремнезёмных спикул. Губки, входящие в группу Sphinctozoa (в основном от ордовика до триаса), имели пористый известковый скелет с кольчатыми перетяжками, напоминающий нитку бус. Изучение современной реликтовой сфинктозойной губки Vaceletia crypta, относящейся к этому же таксону, позволяет сделать вывод о том, что мягкие ткани вымерших Sphinctozoa находились внутри скелета. Черты сходства в строении тканей Vaceletia crypta и обыкновенных губок дают основание относить эту губку (а возможно, и некоторых других Sphinctozoa) к Demospongiae.
Представители Archaeocyatha, Sphinctozoa, Stromatoporata, а также древние Demospongiae в кембрийских и мезозойских морях играли большую роль в образовании рифов. Это заставляет некоторых спонгиологов полагать, что основными тенденциями в эволюции губок были редукция присущих этим древним формам массивных скелетов, из которых формировались рифы, и замена их скелетами из спикул. Низкая скорость роста ныне живущих губок с массивными скелетами по сравнению со скоростью роста кораллов могла быть причиной эволюционного заката рифообразующих губок. Этой гипотезе противоречит тот факт, что хоанофлагелляты, возможно, имеющие с губками общего предка, строят свой скелет из спикул, состоящих из двуокиси кремния.
Систематика губок весьма противоречива даже в тех случаях, когда речь идёт о таксонах самого высокого уровня. В последнее время коралловых губок (ранее самостоятельный таксон Sclerospongiae) включают в группу Demospongiae. Недавно было также предложено выделить два подтаксона — Symplasma (Hexactinellida) и Cellularia, поскольку это формально отражает существующую разницу между синцитиальной организацией Hexactinellida и клеточным строением Calcarea и Demospongiae.
Влияние на культуру

- Персонаж одноимённого культового мультсериала Губка Боб Квадратные Штаны.
Примечания
- ↑ Тип Губки в Мировом реестре морских видов (World Register of Marine Species) (англ.) (Проверено 31 октября 2011)
- ↑ Вестхайде В., Ригер Р. От простейших до моллюсков и артропод // Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — Т. 1. — iv+512+iv с. — 1000 экз. — ISBN 978-5-87317-491-1
- ↑ Шарова И. Х. Зоология беспозвоночных — М.: Гуманит. изд. центр ВЛАДОС, 2005 — С. 108 — ISBN 978-5-87317-491-1
Литература
- Ересковский А. В. Сравнительная эмбриология губок (Porifera). — СПб.: СПбГУ, 2005. — 304 с.
- Колтун В. М. Стеклянные, или шестилучевые, губки Северных и Дальневосточных морей СССР. (Класс Hyalospongiae). Определители по фауне СССР, издаваемые Зоологическим институтом Академии Наук СССР. Выпуск 94. — Ленинград: Наука, 1967. — 128 с. DjVu
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Протисты и низшие многоклеточные // Зоология беспозвоночных. Функциональные и эволюционные аспекты = Invertebrate Zoology: A Functional Evolutionary Approach / пер. с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Т. 1. — 496 с. — 3000 экз. — ISBN 978-5-7695-3493-5
Ссылки
- Популярная статья о губках.
- Тип губки — Porifera
|
Систематика животных |
|||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Губки | |||||||||||||||||||||||||||||||||||||||||
| Пластинчатые | |||||||||||||||||||||||||||||||||||||||||
| Эуметазои |
|
||||||||||||||||||||||||||||||||||||||||
| Устаревшие таксоны | Зоофиты · Мезозои · Черви · Первичнополостные черви · Головохоботные · Членистые |
Типичное строение тела губок
Несмотря на многообразие форм тела и замысловатости внешнего облика, все известные губки по внешней форме подразделяют на три основных типа, получивших следующие названия: аскон, сикон и лейкон.
Аскон — наиболее примитивная форма строения тела губки. Асконоидные имеют небольшое бокалообразное или мешковидное тело, основание которого закреплено на субстрате. В верхней части тела имеется устье — отверстие, иногда называемое оскулумом.

Тело таких губок образовано двумя слоями клеток — наружным и внутренним, между которыми расположено студневидное вещество — мезоглея, состоящая из разных по строению и назначению клеток. Наружный слой клеток («кожа») составлен плоскими клетками — пинакоцитами, которые образуют кроющий эпителий, отделяющий мезоглею от внешней среды. Среди пинакоцитов встречаются более крупные клетки — пороциты, которые имеют внутренний канал, открывающийся наружу и обеспечивающий связь внутренности губки с наружной средой.
Внутренний слой тела выстлан воротничковыми клетками — хоаноцитами, имеющими вытянутую форму и жгутик, основанием закрепленный в плазматическом воротничке, имеющей вид воронки, открытой в сторону внутренней полости губки.
Мезоглея содержит неподвижные звездчатые клетки (колленциты), которые выполняют соединительно-опорные функции, скелетные клетки (склеробласты), а также археоциты — недифферинцированные клетки, способные превращаться во все прочие клетки (скелетные, покровные, половые и т. д.). Звездчатые клетки, по мнению некоторых ученых, являются зачатками нервных элементов, способных передавать раздражения. Однако, это предположение пока не подтверждено результатами исследований. Губки очень слабо реагируют даже на самые сильные внешние раздражения, и передача раздражений от одного участка тела к другому практически незаметна. Это свидетельствует об отсутствии у губок нервной системы.
Связь между внешней средой и внутренней (атриальной) полостью губки осуществляется не только через верхнюю воронку-отверстие, но и через поры в покровных клетках тела.
Сикон является более совершенной конструкцией тела губок. При дальнейшем разрастании мезоглеи, участки атриальной полости покрываются впадинами в местах прохождения радиальных трубок. При этом хоаноциты сосредоточены только в этих впадинах и на внутренней поверхности жгутиковых трубок. Стенки тела губки становятся более толстыми, между внешним слоем и жгутиковыми трубками образуются приводящие каналы, соединяющие внутреннюю полость животного с внешней стороной тела.

Таким образом, у сиконоидных губок хоаноциты выстилают жгутиковые трубки, которые сообщаются с внешней средой, с одной стороны, посредством наружных пор или системы приводящих каналов, а с другой — через атриальную полость и устье. Однако, проводящие каналы в мезоглее сиконоидных имеют вид своеобразных трубок, не имеющих расширений — камер, присутствующих у губок более высокой ступени развития — лейконов.
Лейконоидный тип губок отличается от рассмотренных выше тем, что у них мезоглея разрастается еще больше, чем у сиконоидных, при этом хоаноциты сосредотачиваются в небольших жгутиковых камерах, появляющихся в жгутиковых трубках. Эти камеры не имеют непосредственной связи с атриальной полостью, а сообщаются с ней посредством системы отводящих каналов. Связь с внешней средой поддерживается через наружные поры и приводящие каналы.
Лейкон — самый сложный и прогрессивный тип строения организма губок, большинство этих животных во взрослом состоянии имеют лейконоидное строение тела.
***
Анатомическое строение губки
- Главная страница
- Места обитания губок
- Анатомия губки
- Биология и образ жизни
- Половое размножение губок
- Бесполое размножение губок
- Систематика губок
- Известковые губки
- Стеклянные губки
- Обыкновенные губки
- Четырехлучевые губки
- Кремнероговые губки
- Пресноводные губки
- Ядовитые и опасные губки
- Клионы — сверлящие губки
- «СпанчБоб — каратист»
- «Найди улитку Гэри»
- «Губка Боб — стрелок из лука»
- «Спанч Боб и Патрик — пираты»
Общая характеристика типа Губки
Определение Губок
Губки в биологии или Spongia представляют тип беспозвоночных животных, которые, по всей вероятности, происходят от колониальных воротничковых простейших, у которых есть жгутики.
Тип Губки формирует слепую ветвь в основании филогенетической системы древа многоклеточных. Принято считать, что возникновение губок произошло в докембрии примерно миллиард лет назад. Мезозой же считается периодом расцвета этой группы живых организмов.
Где обитают губки? Под губками понимают преимущественно морских организмов — только небольшая часть гибок является пресноводной. Для них характерен неподвижный образ жизни: они прикрепляются к субстрату и почти не реагируют на раздражение. Обычно губки ведут колониальный образ жизни, но встречаются также единичные особи. У губок твердое и жесткое тело, зачастую — яркая окраска (исключениями в этом плане являются серые и зеленоватые бадяги). Их окраска варьирует и зависит от количества пигмента. Неприятный запах и вкус свойственен для большинства губок.
Несмотря на то, что губки являются довольно примитивными, они довольно разнообразны в видовом плане. В тип Губки входит свыше 8 тысяч видов. К примеру, в Черном море встречается около 300 видов, а в Каспийском — только 1.
На территории России встречаются обычно байкальские виды и некоторые виды бадяг.
Особенности представителей типа Губки
Специфическая черта организации губок — примитивное строение всех систем органов. Тело преимущественно бесформенное, какая-либо симметрия отсутствует. Если говорить о форме, то обычно это бокаловидная и мешковидная, а высота — от нескольких мм. до 1,5 метров. Внутри тела располагается парагастральная полость, которая на вершине открывается устьевым отверстием. У губок отсутствуют полноценные органы и ткани, однако можно отметить заметную тенденцию к специализации клеток. Также губки не имеют единой категории тотипотентных или плюрипотентных клеток.
Среди них выделяют:
- пинаокциты или поверхностные клетки;
- хоаноциты или жгутиковые воротничковые клетки, устилающие внутреннюю полость.
Между двумя обозначенными слоями находится мезоглея. В ней есть амебоциты, склеробласты и прочие группы клеток. Поверхность тела клеток пронизывают многочисленные поры: они открываются в каналы, проходящие насквозь в стенку тела.
Губки можно классифицировать на основании степени развития канальной системы на:
- аксоны;
- лейконы;
- сиконы.
Губки сиконы и аксоны можно соотнести друг с другом как часть и целое. Принято полагать, что сиконовидные губки появились в результате незавершенного почкования — это случилось вследствие высокой интеграции колоний, в которых жгутиковые камеры соответствуют отдельным особям.
Почти у всех губок есть скелет, который состоит из кремниевых или известковых игл. У роговых губок скелет состоит из рогового вещества — спонгина. Губки — отличные фильтраторы, процеживающие воду. Такое мероприятие происходит за счет биения жгутиков хоаноцитов. Позже вода проходит через канальную систему и парагастральную полость. В воде имеется детрит, разнообразные простейшие, бактерии, диатомовые водоросли и другие микроскопические организмы. За захват пищи отвечают хоаноциты и клетки стенок каналов.
Губки преимущественно являются гермафродитами. Из яйца происходит развитие личинки.
Личинка губок представляет собой паренхимулу или амфибластулу.
После того как личинка выходит наружу, определенное время она плавает, после чего оседает на дно и преображается в молодую губку. Развитие губок происходит с метаморфозом. В частности, наблюдается оригинальный процесс извращения зародышевых листков. Происходит миграция клеток наружного слоя внутрь, а клеток внутреннего слоя — на поверхность. Губки отличаются почкованием и образованием геммул. Встречаются яйцеродящие и живородящие губки. Если губки яйцеродящие, то они обычно являются раздельнополыми. Живородящие губки могут быть и гермафродитами.
Встречаются случаи смены губкой пола в течение года.
На основании строения скелета Губки делятся на 4 класса:
- Обыкновенные губки или Demospongiae. У них имеется скелет, который состоит из кремниевых игл. Также для них свойственен лейкоидный тип каналов. В большинстве случаев этот класс губок является колониальным, но встречаются и единичные представители. Обычно это морские животные. Обыкновенные губки — наиболее многочисленный класс, в который входят два отряда: Кремнероговые и Четырехлучевые.
Скелет Кремниевых губок представляет собой комплекс из кремниевых одноосных игл и спонгиновых волокон. В отряд Кремнероговые входят Бадяги и некоторые виды Туалетных губок. Набольшую ценность представляет Греческая губка или Euspongia officinalis.
У Четырехлучевых губок шаровидное тело и кремниевый скелет: иглы на этом скелете располагаются радиально. Основной средой обитания являются воды на глубине 400-450 метров.
- Известковые губки или Calcispongiae. В их основе лежат одноосный, трехлучевой или четырехлучевой скелеты из углекислого кальция. Тело таких губок преимущественно бочковидное и трубковидное. Они отличаются оригинальной особенностью — в них присутствует три типа канальной системы. Представителями этого класса являются губки Сикон, Сикандра, Леукандра и Асцетта.
- Коралловые губки или Sclerosspongiae. Это колониальные организмы, история которых начинается с мезозоя. В основе скелета лежит базальная масса арагонита или кальцита и кремниевые одноосные иглы. Также у этих губок имеется довольно тонкая живая ткань. Канальная система лейконоидного типа.
- Стеклянные губки или Шестилучевые губки (Hyalospongia Hexactinellida). Это организмы, у которых скелет включает шесть кремниевых игл. Основной средой обитания являются океаны. Нередко эти губки используются как украшения: губка корзинка Венеры, Эуплектелла, Хиалонема.
Для губок характерна врожденная иммунная система — они способны различать патогенные клетки и провоцировать их апоптоз.
Значение губок
Значение губок в природе и жизни человека заключается в следующем:
- они представляют собой среду обитания для некоторых животных;
- они обеспечивают фильтрацию воды от мелких частиц;
- они используются в медицине для лечения гематом и неврологических заболеваний;
- туалетные губки применяются в качестве полировального и шлифовального материала: в ювелирном деле и для изготовления фильтров.
Как видно, эти организмы являются довольно обширной группой животных и важным звеном морской экосистемы.
1.
Тип Губки
PORIFERA или SPONGIA
(от лат. «пористые животные»)
Общая характеристика типа губок
Классификация губок
Значение губок
2.
К губкам относятся животные с выраженной (по
сравнению с простейшими) специализацией клеток тела.
Тело губок пористое, состоит из двух клеточных слоев –
наружного и внутреннего. Между ними находится
студенистое вещество с включениями из игл –
известковых, кремниевых или роговых, а также из
клеток, подобных амебе.
Наружный
слой
образован
плоскими
эпителиальными
клетками,
а
внутренний
–
жгутиковидными клетками с окружающими жгутиками
воронковидными выростами цитоплазмы (служат для
захвата пищи), «амёбы» переваривают её, «звезды»
выделяют вещества для построения скелета.
3.
У губок отсутствуют ткани, органы, у них
высокая
регенерационная
способность
и
взаимопревращаемость
многих
клеток,
отсутствие нервных и мышечных клеток.
Им
свойственно
только
внутриклеточное
пищеварение, имеется минеральный, роговой или
смешанной природы скелет, защищающий тело от
механических повреждений и хищников.
Тело пронизано порами. Через них вода
поступает внутрь тела со взвешенными пищевыми
частицами.
С током воды через тело губок пассивно
осуществляются все функции: питание, дыхание,
выделение, размножение.
3
4.
Бокаловидная
форма губок –
аксон; если состоит как бы из
концентрических
рядов
сросшихся
асконов, то это – сикон.
Если тело
губки
пронизано
многочисленными
трубчатыми,
местами
ветвящимися каналами – Три типа строения губок
это лейкон.
5.
Спикулы губок
Скелетные иглы – могут состоять
из кремнезема или извести
6.
Три класса губок
(около 3 тыс. видов):
Известковые
Обыкновенные
Стеклянные
7.
Clathrina coriacea
(белая губка)
Класс Известковые губки
Представители класса, имеющие
известковый (из карбоната кальция)
скелет, обитают на мелководьях морей
и океанов. Могут жить как одиночно, так
и колониально. Диаметр их непрочного
тела не более 7 см, окраска желто –
серая. Скелетные иглы могут достигать
3 см в длину.
Leucandra heathi,
Leucosolenia botryoides
8.
Aphrocallistes,
Farrea
Класс
Стеклянные
губки
В основном это глубоководные формы, высотой
до 50 см. Тело легко повреждается. Живут в основном
одиночно, колонии редки. Окраска тела – белая,
серая, желтая или коричневая. Скелетные иглы
содержат кремний, по величине бывают очень мелкие
(около 1 мм) и очень крупные (до 30 – 40 см).
9.
Класс
Обыкновенные
губки
Представители этого класса имеют кремневый скелет,
состоящий из органического вещества и кремния, и обитают
не только в морях и океанах, но и в пресных водоемах.
Формы, окраска и размеры этих губок очень разнообразны.
Губки колониальные животные, одиночные – редки.
Скелетные иглы у некоторых видов могут отсутствовать.
Тело у одних особей прочное, у других – мягкое, эластичное.
У некоторых видов губки достигают размеров до 1 м.
10.
губка корзина Венеры
(20-30см)
11.
Бадяга
пресноводная
(используют в
медицине для
лечения
ревматизма,
ушибов, синяков)
12.
Губка туалетная
(в Древнем Риме
использовали как
туалетную бумагу (15-30см))
Используют
в
технических
целях,
медицинских,
парфюмерии, удобрений
13.
Губка кубок Нептуна
(1- 1,5 м)
14.
Пробковая губка интересна своим сожительством с ракомотшельником, прячущим свое мягкое брюшко в спиральной полости
внутри губки.
15.
Обработанная губка легко сжимается, а опущенная в
воду быстро восстанавливает свою форму. Ещё во времена
античности ими мылись в римских банях.
16.
Из
губок
делали
специальные подкладки под
латы и смягчали удары и
впитывали
пот,
но
и
останавливали кровь.
17.
Живые фильтры. Самое глубокое озеро в мире Байкал
известно и как самое большое на Земле хранилище пресной воды.
Её уже более 20 млн. лет фильтруют живущие здесь губки двух
родов – Baicalospongia и Lubomirskia. Именно благодаря
деятельности этих живых фильтров байкальская вода прославилась
своей кристальной чистотой и прозрачностью.
18.
19.
dic.academic.ru › dic.nsf/enc_biology/1240
www.worldofnature.ru
http://www.megabook.ru/DescriptionImage.asp?MID=447600&AID=634372
http://vet.apteka.uz/carstvo_jivotnyh/gubki
http://www.worldofnature.ru/index.php?name=Calcispongia_Calcarea
www.point.ru
www.apus.ru
vet.apteka.uz
chegepe33.narod.ru
chegepe33.narod.ru
www.zooeco.com
www.sealifegifts.net
zoo.rin.ru
http://tana.ucoz.ru/load/174
http://www.clean-face.ru/Bodyaga
www.ksu.ru
www.worldofnature.ru
http://www.medpulse.ru/?area=articleSearchResults&offset=80&w=Оптические
материалы&l=1&action=search
http://ts.1in.kz/index.php?showtopic=3628
http://megasklad.ru/lots/view/353393
http://vash-malish.com.ua/type-4-0-112.html
larece.ruhttp://wiki.omskedu.ru/index.php/Викторина
Губки тип классы
Губки — наиболее низкоорганизованные асимметричные или радиально-симметричные многоклеточные животные (табл. I, рис. 57). Тип губок включает около 5000 видов, подавляющее большинство которых обитает в морях и океанах, пресноводных видов немного (сем. Spongillidae).
Систематическое положение — Лучистые или Радиально-смметричные. Латинское Radialia
Тип Губки Латинское название Porifera или Spongia
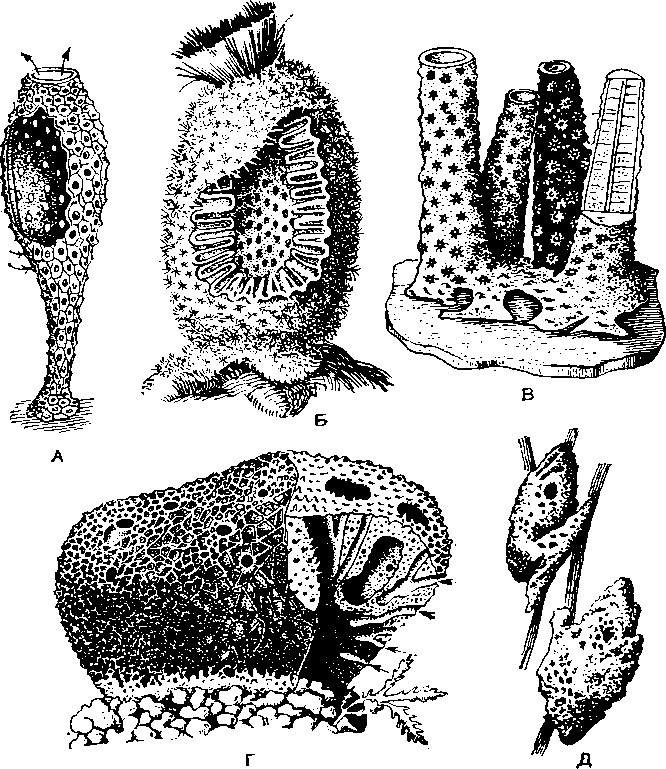
А — Leucosolenia coriacea; Б — Sycon raplianus: В — колония Verongia aerophoba; Г — туалетная губ. (Spongia officinalis): Д — пресноводная бадяга Spongilla.
Общая характеристика губок
Все губ. во взрослом состоянии неподвижные, прикрепленные животные, не имеющие ни нервных клеток, ни органов чувств, ни мускулатуры, ни ротового отверстия. Постоянный ток воды со взвешенными пищевыми частицами проходит через все тело животного. Вода попадает через многочисленные поры (Porifera означает — несущие поры), расположенные на наружной поверхности тела губок, в систему поровых каналов и жгутиковых камер, или полостей, и выводится наружу через устье, или оскулум.
Губки находятся на клеточном уровне организации. Это означает, что их тело представляет собой скопление рыхло расположенных, в разной степени дифференцированных клеток, выполняющих определенные функции: покровную, опорную, вододвигательную, питательную и др. Однако эти клетки еще не образуют настоящих тканей и органов.
У губок различают следующие основные типы клеток:
- Пинакоциты — плоские многоугольные, способные сокращаться клетки, которые выстилают наружную поверхность и поровые каналы Г..
- Воротничков ые клетки, или хоаноциты, цилиндрической или шаровидной формы. Они образуют жгутиковые камеры, а у наиболее примитивных одиночных известковых губок выстилают внутреннюю (гастральную) полость. На свободном конце клетки, обращенном внутрь жгутиковой камеры, расположен жгутик, окруженный цитоплазматическим воротничком (рис. 2). С помощью электронного микроскопа выяснено, что воротничок хоаноцита образован венчиком из 30—40 тесно прилежащих друг к другу тончайших плазматических выростов — щупалец. Жгутик хоаноцита отходит от базального тельца, к которому присоединено чешуевидное парабазальное тельце (рис. 2, В). Такое же ультратонкое строение имеют и воротничковые жгутиковые (Craspedomonadina), что указывает на их филогенетическую близость. Полагают, что Г. и воротничковые жгутиковые имели общего предка. Хоаноциты выполняют вододвигательную и питательную функцию. Биение их жгутиков создает постоянный направленный ток воды в теле Г., а пищевые частицы, приносимые водой, захватываются и перевариваются хоаноцитами. Весь процесс пищеварения у губок проходит внутриклеточно. Непереваренные остатки пищи из пищеварительных вакуолей выбрасываются с током воды через оскулум.
- Амебоциты — крупные блуждающие клетки, они осуществляют фагоцитоз и участвуют в переваривании пищи, получая излишек пищевых частиц от хоапоцитов. Амебоциты постоянно передвигаются в студенистой массе основного вещества Г. — мезоглее, которая заполняет все пространство между поверхностью Г. и системой водоносных каналов. Мезоглея связывает все типы клеток механически и химически, или гуморальн
- Колленциты — звездчатые клетки, соприкасающиеся своими отростками с соседними клетками, они выполняют опорную функцию.
- Склеробласты и сионгиобласты — крупные клетки, формирующие элементы скелета Г. — спикулы и спонгинотше волокна.
- Археоциты — не дифференцированные клетки, способные легко превращаться в клетки любого типа, служат клеточным резервом.
- Половые клетки, образующиеся из археоцитов, обеспечивают половое размножение.
Все типы клеток губок способны при некоторых условиях переходить в амебоидное состояние и превращаться в клетки иного типа. Даже рано обособившиеся хоаноциты, по последним данным, способны терять воротнички и жгутики, уходить в глубь мезоглеи и превращаться там в амебоциты. Губ. можно разрезать на маленькие кусочки и протереть через мелкоячеистое сито (мельничный газ), при этом клетки будут отделены друг от друга. Если протертую массу поместить на дно сосуда, то клетки будут сползаться и образовывать группы, или агрегаты, клеток, которые через несколько дней превратятся в маленьких губок. Такая высокая р е-ге нерационная способность губок объясняется относительно слабой дифференцировкой их клеток и отсутствием настоящих тканей.
Г., как и все многоклеточные, размножаются половым путем. Формирование половых клеток — яйцеклеток и сперматозоидов — происходит обычным для всех многоклеточных способом. Кроме полового размножения, губ. присуще бесполое размножение — путем наружного почкования или образования покоящихся внутренних почек — геммул.
Внешний вид и внутреннее строение
Губ. совсем не похожи на животных, они скорее напоминают растения. Недаром их до начала XIX в. относили к сборной группе зоофитов, будто бы промежуточной между растениями и животными. Форма губок крайне разнообразна. Многие из них образуют наросты и корки на субстрате, караваи, глыбы. Немало губок имеет чашевидную, кустистую, трубчатую, грибовидную (рис. 1, А, Б, В; табл. I) форму. Неопределенность формы тела многих губок зависит от того, что они образуют колонии, состоящие из многих особей. При разрастании колонии обычно теряет индивидуальность составляющих ее особей (рис. 1, Г, Д). Отдельные особи ясно различимы только у немногих колониальных губок (рис. 1, В; табл. I). Одиночных губок сравнительно немного. Обычно они имеют более правильную форму — вазы, кубка или бочонка. У одиночных губок можно выявить радиальную многолучевую симметрию.
На ощупь тело большинства губок жесткое, что объясняется присутствием твердого скелета — известкового, кремневого или кремнерогового.
Размеры губок колеблются от нескольких миллиметров до 1—1,5 м в длину и до 2 м в диаметре. Многие морские виды ярко окрашены, чаще в желтые, оранжевые, красные, пурпурно-фиолетовые и коричневые тона. Окраску обусловливают разнообразные пигментные клетки, присутствующие в мезоглее. Некоторые гГ. лишены пигмента и выглядят белыми или сероватыми. Пресноводные Г. бадяги окрашены в коричневатый или зеленый цвет. Последний зависит от присутствия симбиотических одноклеточных зеленых водорослей — зоохлорелл.
Большинство губок имеет резкий, неприятный запах, который зависит от выделения ядовитых веществ. Само тело губок также обладает токсическими свойствами, имеющими, по-видимому, защитное значение и обусловливающими наряду с твердым скелетом несъедобность губок. Предполагают, что запах и токсичность губок обусловлены накоплением значительного количества продуктов метаболизма в отдельных клетках и в мезоглее губ.
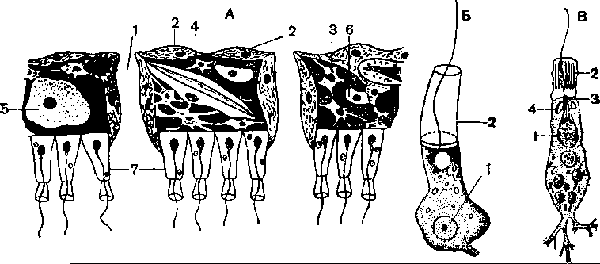
А разрез через часть стенки тела Г. асконоидного типа; 1 — поровый капал; 2 — пина-коциты; 3 — звездчатые клетки мезоглеи; 4 — склеробласт с образующейся внутри клетки спикулой; 5 — яйцевая клетка; 6 — археоцпт; 7 — хоаноциты; Б — хоаноцит; В — ультратонкое строение хоаноцита известковой Г. r.Sycon raplianus); 1—ядро; 2 — плазматический воротничок; 3 — баззльксе тельце; 4 — парабазальное тельце.
Внутреннее строение губок легче всего изучать на одиночных формах. При этом различают три типа их организации: а с коноидный, или аскон, сиконоидный, или сикон, и лейконоидный, или лейкон.
Наиболее просто устроены очень маленькие одиночные (по крайней мере молодые) известковые Г. Leucosolenia coriacea (рис. 1, А; рис. 3, А). Их радиально симметричное тело похоже на вазу. Тонкая стенка тела (около 2 мм) ограничивает центральную гастральную, или атриальную, полость губ.. Последняя одета изнутри почти сплошным слоем хоаноцитов, образующих примитивный рыхлый жгутиковый эпителий, который свидетельствует о зачаточном тканеобразовании у губок. Наружпая дермальная поверхность тела покрыта преимущественно пинакоцитами (рис. 2, Л), перемешанными с клетками других типов. Весьма многочисленны трубкообразные пороциты, соединяющие наружную поверхность гГ. с гастральной полостью. Их внутриклеточный. просвет образует поры, по которым вода поступает в гастральную полость. Между дермальной поверхностью и слоем хоаноцитов в студенистом веществе мезоглеи расположены многочисленные звездчатые колленциты, склеробласты со спикулами и образованные ими трехосные спикулы, различной формы амебоидные клетки — амебоциты и археоциты, веретеновидные сократимые клетки
— миоциты и др.
<
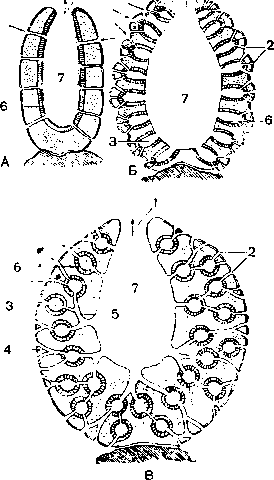
А — асконоидный; Б — сиконоидный; В — лейконоидный; 1 — оскулум; 2 — поры; 3 — жгутиковые камеры; 4 — приносящие каналы; 5 — выносящие каналы; 6 — хоаноциты; 7 — гастральная полость. Стрелки показывают направления . токов воды.
Непрерывное биение жгутиков хоаноцитов прогоняет воду из гастральной полости через выводное отверстие — оскулум — наружу, а через поры в тело Г. поступают все новые порции воды.
Этот тип организации называется аскон, он встречается у небольшого числа видов известковых губок или у молодых губок, строение которых во взрослом состоянии усложняется. Большинство губок устроено сложнее.
Усложнение организации губок (тип сикон) наблюдается у небольших известковых губок из рода Sycon и др. (рис. 1, Б; 39, Б). Стенка тела этих губок утолщается, в ней образуются многочисленные цилиндрические впячивания атриальной полости. Хоаноциты располагаются только в этих впячиваниях, которые превращаются в жгутиковые каналы. Атриальную полость сиконоидной Г. выстилают только пина-коциты. Вода поступает через межклеточные поры в норовые каналы, выстланные пинакоцитами, затем в жгутиковые каналы, а оттуда в атриальную полость и выводится через оскулум наружу.
Большинству крупных одиночных и колониальных губок свойственно дальнейшее усложнение организации. При этом жгутиковые каналы разбиваются на маленькие шарообразные жгутиковые камеры, которые лежат в несколько слоев внутри еще более утолщенной стенки тела Г.. Все жгутиковые камеры связаны с внешней средой сложной системой приводящих каналов, а с атриальной полостью — системой отводящих каналов. Этот наиболее сложный тип строения называют лейконоидным (рис. 59, В). Прогрессивное увеличение числа маленьких жгутиковых камер приводит к значительному повышению их фильтрационной способности. Так, у небольшой Г. Leucandra aspera, высотой 7 см и диаметром 1 см, подсчитано 2 250 000 жгутиковых камер с общей поверхностью 52,5 см2, 81 000 приносящих и 5200 отводящих каналов. За сутки такая губ. фильтрует 22,5 л воды.
У многих губок вторично образуются защитные дермальные мембраны, или корковый слой, покрывающий снаружи тело губ.. Атриальная полость может сильно сокращаться, мало отличаясь от отводящих каналов, особенно у колониальных форм. У последних обычно невозможно различать отдельные особи, входящие в состав колонии. Об их количестве можно судить лишь по числу оскулумов Г..
Питание губок осуществляется хоаноцитами и амебоцитами. Хоа-ноциты захватывают мелкие пищевые частицы (детрит, бактерии, одноклеточные водоросли и т. п.) короткими псевдоподиями, образующимися несколько ниже воротничка. При этом формируются типичные пищеварительные вакуоли, совершающие круговые движения в цитоплазме хоа-ноцита. Непереваренные остатки пищи выбрасываются из сократившихся в размерах вакуолей в зоне воротничка. Крупные хоаноциты известковых губок захватывают пищевые частицы, а затем передают их прилегающим к ним амебоцитам, которые их переваривают и разносят питательный материал по телу. Более крупные частицы фагоцитируются непосредственно амебоцитами, подходящими к стенкам каналов.
Дыхание и выделение у губок осуществляются диффузно. Кислород, растворенный в воде, которая проходит через все тело губ., используется ею для дыхания (газообмена). Жидкие продукты метаболизма и экскременты из пищеварительных вакуолей хоаноцитов и амебоцитов поступают через систему каналов в атриальную полость, а оттуда с током воды выбрасываются через оскулум наружу.
До настоящего времени не выяснено, каким путем осуществляется координация между всеми клеточными элементами губок. Французские ученые (Тюзе, Паван, Грассе и др.) описали биполярные и мультиполяр-ные паукообразные и пузыревидные клетки, образующие связи с клетками других типов. Они считают их за нервные клетки, образующие диффузную нервную систему. Однако у губок крайне низка способность реагировать даже на грубые внешние раздражения (химические, термические и др.). Губ. можно проткнуть раскаленным металлическим стержнем, однако реакция на это раздражение будет распространяться по телу очень медленно и на незначительное расстояние. Все движения взрослой губ. сводятся к медленному закрыванию пор и оскулума. Все это дает основание большинству ученых отрицать наличие у губок как нервной, так и мышечной системы.
Скелет
Подавляющее большинство губок имеет скелет. Особенности в строении и составе скелетных образований являются важнейшим систематическим признаком. Скелет у различных групп губок может быть известковым, кремневым, кремнероговым и роговым, или спонгиновым.
Известковый скелет свойствен сравнительно небольшой группе исключительно морских, чаще одиночных губок (Leucosolenia, Sycon и др.). Он образован различными по форме иглами, или спикулами, состоящими из углекислого кальция. Различают одно-, трех- и четырехлучевые иглы (рис. 4, А). У сикона и других губок (рис. 57) они окружают жгутиковые камеры и оскулум, причем концы их могут торчать наружу.
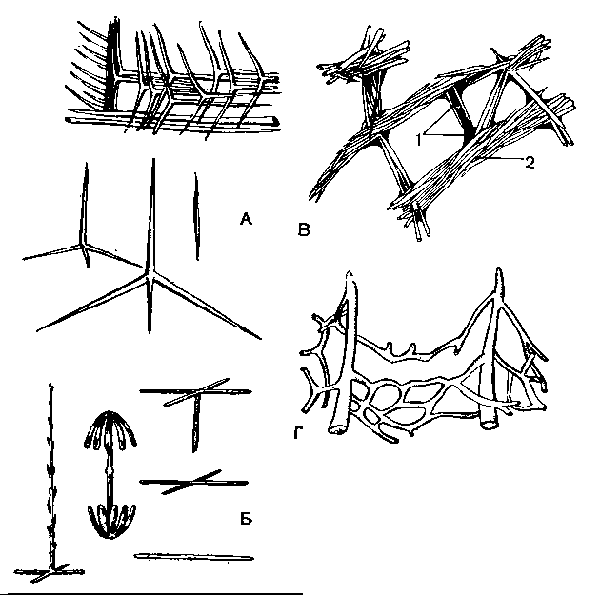
А — спикулы известковых губок; Б — кремневые спикулы стеклянных губок; В — скелет бадяги; 1 — спонгиновые волокна; 2— кремневые спикулы; Г — спонгиновый скелет туалетной губки.
Спикулы образуются особыми клетками мезоглеи — склеробла-стами. Отложение углекислого кальция и начало образования спикулы происходит внутри склеробласта. Когда спикула достигает размеров, превышающих размеры склеробласта, последний отмирает, а игла остается в мезоглее.
Формирование трех- и четырехлучевых спикул происходит при совместной деятельности нескольких склеробластов, образующих однолучевые спикулы, которые одновременно спаиваются своими концами под определенным углом.
Кремневый скелет образован спикулами, состоящими из кремнезема, который отлагается в склеробласте вокруг осевой нити. Последняя состоит из органического вещества.
Кремневые спикулы бывают двух родов: более крупные — макросклериты, составляющие основу скелета, и мелкие — микросклериты. Кремневые спикулы весьма разнообразны по форме (рис. 60, Б). Макросклериты могут быть одно-, трех- и четырехосными, достигая у некоторых губок больших размеров. Так, у стеклянной Г. Hyalonema (рис. 63, Б) «корневые» нити имеют длину в несколько десятков сантиметров, а у губ. Monorhaphis так называемая осевая спикула достигает длины 3 м
Кремнероговой скелет имеется у бадяг и некоторых других губок (рис. 4, В). Он состоит из кремневых спикул и волокон, образованных особым органическим веществом — спонгином. Спонгин — сложное органическое вещество, по составу близкое к шелку, содержащее иод (до 14%). Волокна спонгина образуются не внутри отдельных клеток, а межклеточно. Мелкие клетки спонгиобласты как футляром одевают растущее волокно.
Роговой, или спонгиновый, скелет, свойственный небольшому числу губок (туалетная губ.), вследствие редукции кремневых спикул состоит только из спонгина (рис. 60, Г).
Некоторые очень мелкие губ. полностью лишены скелетных образований. По-видимому, утеря минерального скелета роговыми и бесскелет-ными губками произошла вторично.
Губкам свойственно как бесполое, так и половое раз-Размпожение множение. Бесполое размножение губок проис-и развитие ходит двумя способами: наружным и внутренним почкованием.
Наружное почкование заключается в образовании небольших вздутий — почек, затем разрастающихся и формирующих собственный оскулум. У большинства губок дочерние особи не отделяются от материнской, и образуется колония. У одиночных губок дочерние особи отделяются от материнской и переходят к самостоятельному существованию.
Внутреннее почкование наблюдается у пресноводных бадяг и некоторых морских губок. У бадяг осенью археоциты собираются в группы и вокруг них образуется двойная хитиноидная оболочка. Наружный слой этой оболочки имеет воздушные камеры. Склеробласты, окружающие внутреннюю почку, или гем му л у (рис. 61), образуют особые склериты, так называемые амфидиски, обычно состоящие из колонок с звездообразными расширениями на концах. Амфидиски располагаются между двумя слоями оболочки.
Осенью колонии бадяг погибают, а внутри их сохраняются жизнеспособные геммулы. Они хорошо переносят промерзание и высыхание водоема. Геммулы могут оставаться на месте старой колонии или, при разрушении ее, переноситься течением, ветром или водоплавающими птицами. Таким образом, геммулы обеспечивают как переживание губок в течение неблагоприятного времени года, так и их расселение.
Весной из геммул выходят археоциты и образуют новую губ., причем содержимое нескольких геммул часто сливается вместе и развивается в одну колонию.
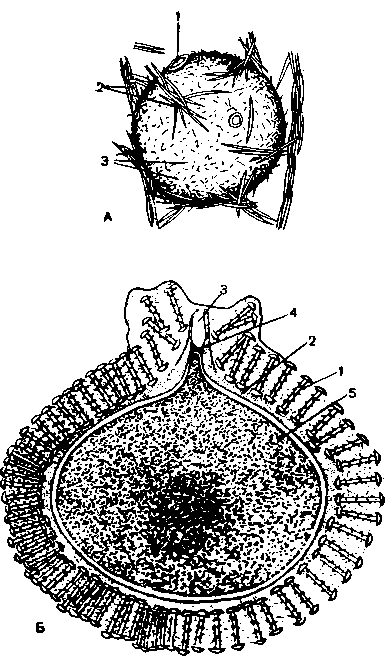
А — геммула озерной бадяги (Spongilla lacust-ris); 1 — поровое отверстие; 2 — мегасклеры; 3 — микросклеры; Б — разрез через геммулу бадяги Ephydatia blembingia; 1 — амфидиски; 2 — внутренняя кутикулярная оболочка; 3 — поровое отверстие; 4 — замыкающая его перегородка; 5 — внутренняя масса клеток.
Половоеразмножение свойственно всем губ. Значительная часть губок — гермафродиты, но многие, в том числе пресноводные бадяги, раздельнополы. У раздельнополых губок одни колонии образуют только яйцевые клетки, а другие — только сперматозоиды. Половые клетки образуются в мезоглее из археоцитов. Женские половые клетки — яйца, или ооциты, остаются в мезоглее, где и происходит их оплодотворение. Мужские половые клетки — сперматозоиды — выходят в воду и проникают с током воды в соседние колонии.
Оплодотворение у всех губок проходит очень своеобразно. Сперматозоид проникает через воротничок внутрь хоаноцита, где теряет хвост и инцистируется. Такой хоаноцит со сперматозоидом внутри переходит в амебоидное состояние и превращается в клетку носительницу. Последняя мигрирует в мезоглею, где поглощается ооцитом. При этом сливаются гаплоидные ядра ооцита и сперматозоида, т. е. происходит процесс оплодотворения.
Дробление оплодотворенного яйца у губок происходит обычно внутри материнского организма, где образуются личинки. Развитие у губок протекает очень разнообразно и подчас весьма сложно.
У некоторых известковых губок (Sycon raphanus) дробление яйца проходит в мезоглее под слоем хоаноцитов (рис. 6, А). В результате его образуется бластула, у которой восемь крупных клеток тесно прилегают к хоаноцитам, а мелкие, более многочисленные жгутиковые клетки составляют ее вторую часть, расположенную свободно в мезоглее. Жгутики мелких клеток направлены внутрь полости бластулы. Затем в центре бластулы раздвигается группа крупных клеток и образуется отверстие, через которое становятся видны усиленно размножающиеся жгутиковые клетки. Внутренняя часть личинки со жгутиковыми клетками выгибается в виде купола по направлению к слою хоаноцитов, а крупные клетки отходят от хоаноцитов, располагаются по краям отверстия, а затем смыкаются на полюсе, противоположном их первоначальному положению, где ранее были мелкие клетки. Этот процесс называется экскурвацией или вы-ворачиванием поверхностей зародыша. Такого изменения, или инверсии, поверхностей бластулы у других многоклеточных не наблюдается. У простейших аналогичный процесс известен только при формировании колоний вольвокса. Крупные клетки впячиваются при этом внутрь образовавшейся амфибластулы (рис. 62, А). Затем они вновь выпячиваются, и шаровидная личинка покидает материнский организм. Амфи-бластула свободно плавает за счет биения жгутиков, расположенных на ее переднем полюсе, а задний полюс образован крупными зернистыми клетками (рис. 6, А).
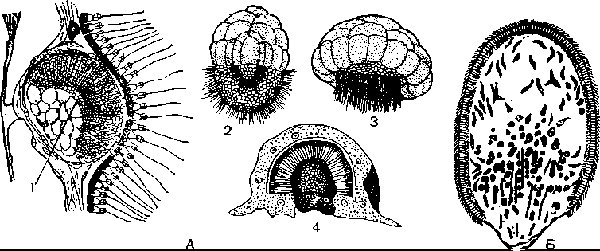
А— развитие губки Sycon raphanus; l — амфнбластула в теле материнской губ.и (видно впя-чивание крупных клеток); 2 — плавающая амфнбластула; 3 — впячпвание мерцательных клеток; 4 — прикрепившаяся личинка; Б — личинка кремнероговой губ. МухШа.
После короткого периода подвижного существования амфнбластула прикрепляется передним полюсом к поверхности дна. При этом мелкие жгутиковые клетки впячиваются внутрь полости амфибластулы, а крупные зернистые образуют наружный слой, т. е. происходит процесс гаст-руляции. Бластопор, которым личинка прикрепилась к субстрату, закрывается, жгутиковые клетки теряют жгутики (де дифференцируются) и усиленно размножаются. Позднее из них образуются хоаноциты жгутиковых каналов, а наружные крупные зернистые клетки превращаются во все остальные клетки тела Г. (рис. 6, А).
У некоторых обыкновенных губок, например у Oscarella lobularis и других, в материнском организме развивается однослойная бластула — целобластула, равномерно покрытая жгутиками. Она покидает материнскую губ. и свободно плавает. Ее задний полюс образован более длинными красноватыми клетками, а клетки переднего полюса бесцветные. Затем она садится на дно передним полюсом, бесцветные клетки которого впячиваются внутрь, т. е. происходит типичная гаструляция путем инвагинации. Наружные красные клетки теряют жгутики, часть из них распадается. Бластопор закрывается, и происходит сложный процесс формирования жгутиковых камер за счет жгутиковых клеток переднего полюса, все остальные части тела Г. (система каналов, выстилка наружной поверхности и атриальной полости, скелет и т. п.) образуются путем размножения и дифференцировки клеток заднего полюса целобластулы.
У части обыкновенных губок (Monaxonida, Keratosa) губку-мать покидает более сложно организованная личинка — паренхимула (рис. 62, Б). Снаружи она почти целиком покрыта жгутиковыми клетками (у некоторых видов на заднем полюсе нет жгутиков), а внутри ее находятся рыхло расположенные клетки, которые у отдельных видов способны дифференцироваться на амебоциты и склеробласты. Еще до вылупления личинки из материнской губ. в ней начинается формирование спикул.
После короткого периода свободного плавания паренхимула прикрепляется передним концом к дну и сильно уплощается. Все жгутиковые клетки ее мигрируют внутрь тела (поодиночке или группами), где их жгутики исчезают. Позднее эти клетки превращаются в хоаноциты и образуют жгутиковые камеры. Одновременно внутренние клетки обрастают всю наружную поверхность Г.. Часть внутренних клеток мезоглеи выстилает приводящие и отводящие каналы, атриальную полость, прорывающийся оскулум. Этот способ развития губ. соответствует поздней гаст-руляции путем мультиполярной иммиграции и эпиболии прочих многоклеточных.
У пресноводных губок бадяг (Spongilla lacustris) наблюдается еще большая эмбрионизация развития. В материнском организме губ. в паренхимуле происходит формирование не только амебоцитов и склеробла-стов, но и жгутиковых камер. Такую паренхимулу правильнее считать молодой губ., еще не имеющей системы каналов. Она появляется из оскулума матери, покрытая жгутиковыми клетками. После ее фиксации на грунте водоема жгутиковые клетки мигрируют внутрь мезоглеи, где их фагоцитируют амебоциты.
Таким образом, в данном случае жгутиковые клетки выступают как временный личиночный орган движения. Дальнейшее развитие такой губ. сводится к построению системы каналов, выстилке дермальной поверхности, атриальной полости и т. п.
Некоторые ученые рассматривают первое впячивание крупных клеток амфибластулы известковых губок, проходящее в материнской губке, как истипную гаструляцию. Тогда впячивание мелких жгутиковых клеток ее заднего полюса после оседания амфибластулы на дно будет вторичным, а не настоящей гаструляцией. Соответственно эти ученые считают губок животными с вывернутыми слоями. При развитии обыкновенных губок поверхностные жгутиковые клетки паренхимулы мигрируют внутрь и превращаются в пищеварительные клетки — хоаноциты. Это тоже как бы подтверждает вывертывание зародышевых слоев тела, в котором видели коренное отличие губок от всех остальных многоклеточных животных. Более подробное изучение эмбрионального развития губок показало, что у некоторых форм, описанных выше, никакого выворачивания слоев не происходит. Более того, у пресноводных губок поверхностные и жгутиковые клетки играют роль провизорного личиночного оргапа передвижения и в построении тела взрослой Г. участия не принимают.
Поверхностные жгутиковые клетки личинок губок можно рассматривать как кин облает — двигательный слой. При переходе к сидячему образу жизни он у взрослой губ. уходит внутрь и выполняет вододвигательную функцию. Фагоцитобласт — внутренний слой — дает все типы клеток губок, как типичных фагоцитов, так и выстилающих пограничные поверхности (наружную дермальную поверхность и систему каналов). Только в этом смысле можно говорить об извращении первичных слоев у губок. Никакой экто- и энтодермы, т. е. настоящих покровных и пищеварительных тканей, у губок еще нет, как нет у них и настоящих зародышевых листков. О крайне низком уровне клеточной интеграции свидетельствует и разнообразие типов их индивидуального развития.

Классификация
Основным признаком, на котором основывается классификация губок, служит строение их скелета. Тип губок разделяется на три класса: 1. Известковые губки (Calcarea); 2. Стеклянные губки (Hyalospongia); 3. Обыкновенные губки (Demospongia).
По современной классификации:
Тип/Отдел Porifera • 9,092 из предполагаемых 8,659 видов (100%) и ископаемые
Класс Calcarea — Известковые губ. • 775 современных вида; и ископаемые
Класс Demospongiae — Обыкновенные губ. • 7,532 современных вида; и ископаемые
Класс Hexactinellida Шестилучевы́е гу́б., или стекля́нные гу́б. • 664 современных вида и ископаемые
Класс Homoscleromorpha • 120 современных вида;
Класс Not assigned • 1 современный вид
Класс Calcarea — Известковые губки
Известковые губ. — исключительно морские мелкие, часто одиночные формы. Их скелет образован известковыми спикулами. Обычно это обитатели мелководий: Leucosolenia, Sycon, Sycetta, Leuconia и др« (рис. 1, А, Б).
Класс Hexactinellida Шестилучевы́е гу́бки, или стекля́нные гу́бки
К этому классу относятся также только морские, но глубоководные губ. с кремневым скелетом, образованным из спаянных вместе трехосных спиьул (макросклеритов). К ним принадлежат Eupiectella (рис.7, А), Hyalonema (рис. 7, Б) с пучком длинных хвостовых кремневых игл, которыми они удерживаются в илистом грунте, Monorhaphis и др.
Класс Demospongiae — Обыкновенные губки
Наиболее сложно организованные (лейконоидные) губки с кремневым скелетом из четырехосных или одноосных макросклеритов, часто укрепленных спонгиновыми волокнами. Обыкновенные Г. наиболее многочисленны и разнообразны. Этот класс включает несколько отрядов.
Скелет этих морских губок образован исключительно четырехосными спикулами. В Баренцевом море в большом количестве встречается Geodia (табл. I), образующая крупные шаровидные колонии, до 50 см в диаметре. Некоторые представители этого отряда — Oscarella, Halisarca — лишены спикул; их называют мясистыми губками.
ОТРЯД 2. ОДНООСНЫЕ ГУБКИ (MONAXONIDA)
Скелет этих губок образован одноосными кремневыми спикулами, связанными сетью спонгиновых волокон в прочную массу. У некоторых видов спикул становится мало, а спонгиновых волокон больше.
Одноосные крайне разнообразны, к ним относятся как морские, так и пресноводные виды. Среди морских губок известен кубок Нептуна (Poterion neptuni), который обитает в Тихом океане и достигает 1,5 м. Интересны также сверлящие губ. (Cliona), поселяющиеся в известняках, на раковинах моллюсков и на коралловых полипах.
Большая группа широкораспространенных пресноводных губок бадяг (сем. Spongillidae) образует корки или массивные бесформенные наросты (рис. 57, Д) на дне водоемов (на камнях, стеблях водных растений и т. п.), часто окрашенные в зеленый или бурый цвет. Байкальские Г. (табл. I) на небольших глубинах образуют кустистые заросли до 1 м высотой.
ОТРЯД 3. РОГОВЫЕ ГУБКИ (KERATOSA)
Обыкновенные губки, скелет которых не содержит кремневых спикул, а образован только сетью спонгиновых волокон. Обычно массивные крупные формы: конская губка (Hyppospongia communis), достигающая 90 см в диаметре, веронгия (Verongia, рис. 57, В) и др. Наибольшее значение имеет туалетная губка (Spongia officinalis), обычная в Средиземном море, которая издавна служит предметом промысла (рис. 57, Г).
Класс Homoscleromorpha
Отряд морских губок, выделяемый в монотипический класс Homoscleromorpha, ранее рассматривавшийся в составе класса обыкновенных губок. Цитологическая организация отличается от всех остальных губок. … По современным филогенетическим данным, Homoscleromorpha — чётко ограниченная монофилетическая группа губок, независимая от обыкновенных губок. По данным сайта World Porifera Database, на июль 2016 года в отряд включают следующие таксоны до рода включительно: Семейство Oscarellidae Lendenfeld, 1887.
ФИЛОГЕНИЯ ГУБОК
Губ.. — очень своеобразная группа многоклеточных животных. По уровню организации они занимают промежуточное положение между простейшими и кишечнополостными. Общность ультратонкого строения во-ротничковых клеток губок и воротничковых жгутиконосцев Craspedomo-iiadina говорит о том, что они произошли от одного предка.
Будучи типичными многоклеточными животными, Г. обладают некоторыми чертами простейших:
1. Хоаноциты расположены рыхло, они могут уходить в мезоглею и переходить в амебоидную форму.
2. Многие типы клеток губок имеют амебоидную природу. Так, женские половые клетки — ооциты — двигаются при помощи псевдоподий, вылезают в атриальную полость в поисках питательных клеток и уползают обратно.
3. При оплодотворении ооцита в нем образуется углубление — клеточный рот; он захватывает инцистированный сперматоцит, как амеба захватывает добычу.
4. Процесс инверсии поверхностей, имеющий место при развитии ам-фибластулы известковых губок, известен только у простейших (Volvoci-dae). Последнее вовсе не говорит о родстве губок и вольвоксовых. По-видимому, губ. произошли от колоний примитивных воротничковых жгутиковых, подобных вольвоксу.
[sch:11]
Губ. ничем принципиально не отличаются от остальных многоклеточных животных. Их хоаноциты, или пищеварительные клетки, располагаются в один слой, как и у остальных Metazoa. Воротничковые клетки имеются и у других многоклеточных: у некоторых иглокожих (морские ежи, голотурии) и кишечнодышащих. Образование половых клеток (сперматогенез и оогенез) и строение сперматозоидов не отличаются от таковых даже высокоорганизованных многоклеточных. У некоторых обыкновенных губок (Demospongia) ^совсем не происходит выворачивания первичных (зародышевых) слоей При развитии известковых губок проявляется четырехлучевая симметрия (четыре крупные клетки образуют крест), весьма характерная для кишечнополостных, личинок полихет, га-стропод и сипункулид. О родстве губок с остальными многоклеточными свидетельствует также сходство личинок губок с личинками кишечнополостных и иглокожих.
Все приведенные сведения подтверждают теорию общего происхождения губок и остальных многоклеточных.
Отсутствие нервной и мышечной систем у губок говорит об их примитивности. Губок можно поставить у самого начала общего ствола Metazoa.
Глядя на причудливую форму и яркую окраску странных объектов, прикреплённых к стенкам морских гротов, коралловым полипам или просто ко дну, трудно поверить, что это не грибы или растения. Но, несмотря на всю внешнюю необычность, губки (Porifera — «несущий поры») — это многоклеточные животные, на взрослой стадии ведущие малоподвижный образ жизни. Они первыми из Metazoa освоили способ питания взвешенными органическими веществами и успешно используют его до сих пор. Поэтому им удобно вести прикреплённый образ жизни.
Основные особенности животных типа Губки
Всего описано около 15 000 видов губок. Подавляющая их часть обитает в морях, только около 219 видов освоили пресные водоёмы. Они встречаются везде, где есть прочный субстрат, подходящий для прикрепления: коралловые рифы, скалы, затопленные брёвна, раковины моллюсков. Некоторые селятся на иле или песке. Во взрослом состоянии почти все они неподвижны, только малая их часть способны на весьма ограниченные перемещения и движения отдельными участками тела.

Большая часть представителей типа предпочитает мелководье. Но некоторые, например, почти все стеклянные губки, встречаются на большой глубине. Размер тела губок варьирует от нескольких миллиметров до трёх метров и более в диаметре и в высоту (как, например, у бочонковидной губки Spheciospongia vesparium). Они живут колониями или одиночно.
- В процессе эволюции губки приобрели многоклеточное тело, идеально приспособленное к добыванию пищи путём фильтрации. Они непрерывно перестраиваются, чтобы настроить фильтрационную систему. Перестройка осуществляется подвижными амёбоидными клетками, способными трансформироваться (превращаться) в другие типы клеток.
- У них нет постоянных тканей, так же как и клетки, они непрерывно изменяются. Это даёт возможность некоторым учёным предполагать, что губки — переходная форма между колониями простейших и многоклеточными животными. Чаще говорят, что у губок нет настоящих тканей, так же написано и в школьных учебниках, это утверждение мы прибережём и для всех экзаменов и проверочных.
- Тело губок может быть радиально-симметричным (шар, конус, цилиндр), но чаще всего оно асимметрично. Общей закономерности в типе симметрии нет даже среди особей одного вида.
- Они бывают ярко окрашенными: пурпурными, красными, зелёными, жёлтыми, оранжевыми. Но встречаются и представители с более скромной расцветкой: серой или коричневой. Окраска обеспечивается наличием клеточных пигментов или симбиотических организмов.
- Им обычно (не всегда) присущ неограниченный рост, что характерно не для животных, а для растений.
- У губок хорошо развита способность к регенерации.
- Большинство имеют скелетные элементы из органического, минерального веществ или из того и другого вместе.
- Для них характерно внутриклеточное пищеварение.

Внешнее строение губок
Приспособленное к фильтрационному типу питания тело губок соответствует одному из трёх вариантов строения: асконоидному, сиконоидному или лейконоидному. Недавно наряду с выделением нового класса губок добавился и ещё один тип их строение — силлиибидный. Наиболее простой, асконоидный тип, представляет собой полый цилиндр, основанием прикреплённый к субстрату.

Все губки — двухслойные животные:
- поверхность их тела покрыта пинакодермой (эктодерма) — слоем плоских клеток;
- внутренняя полость губки носит название атриум, или спонгоцель (парагастральная (атриальная) полость) она выстлана хоанодермой (энтодерма), состоящей из одного слоя жгутиковых клеток, схожих по строению с воротничковыми жгутиконосцами.
Все стенки цилиндра-бокала пронизаны порами — остиями. На верхнем конце тела губки расположено наиболее крупное отверстие-устье — оскулум. Хоанодерма движением жгутиков создаёт ток воды в одном направлении. Она входит в тело губки через остии, попадает в атриум и удаляется через оскулум. Места, где циркулирует вода в теле губки, называется водоносной системой.

Губки с аксоноидным строением не бывают крупными. Их цилиндрические тела обычно не превышают 1 мм в диаметре. Такой тип строения накладывает ограничения на их размер, поскольку увеличение диаметра тела в этом случае нарушает оптимальное соотношение площади поверхности и объема. У особей рода Leucosolenia тело имеет вид отдельных трубок, часто соединённых своими основаниями, напоминающих пучки. У видов рода Clathrina оно организовано в форме трубчатой сети.
Чтобы увеличить свои размеры, губкам пришлось изменить строение тела. Сиконоидный тип строения позволяет увеличить площадь поверхности и сократить объём атриума за счёт глубоких трубчатых впячиваний на внутренней и внешней поверхности тела. Впячивания хоанодермы (камеры и отводящие, или жгутиковые каналы) направлены от центра наружу. Впячивания пинакодермы называются приводящими каналами. Многочисленные отверстия — прозопиле, соединяют приводящие каналы и жгутиковые камеры.

Вода в теле сиконоидных губок проходит следующий путь: остии — приводящие каналы — прозопили — жгутиковые каналы — атриум — оскулум. Губки с таким строением тела имеют больший диаметр — от 1 до нескольких см. Сиконоидное строение характерно для родов Grantia и Sycon.
Наибольшего размера (от нескольких см до 1 метра и более) достигают представители типа с лейконоидным телом. Вода в их, лишённых крупных полостей телах, проходит по сложной сети сосудов. Вместо одной полости в них расположены многочисленные (до 10 000 шт.) небольшие сферические камеры, покрытые жгутиковыми клетками, к которым подходят приводящие и отводящие каналы. Оскулумов, как и отверстий отводящих каналов, на теле таких губок много.

Любое увеличение их тела просто приводит к появлению дополнительных жгутиковых камер. Форма лейконоидных губок разнообразна: они бывают корковыми, подушковидными, ветвистыми, пальцевидными, плоскими, трубчатыми и похожими на вазы.
В настоящее время самой крупной губкой считается вид, пока не имеющий чёткого описания и названия из семейства Rossellidae (найден он на глубине 2,1 км вблизи Гавайских островов). Он достигает 3,5 м в длину, 2 м в высоту и 1,5 м в ширину.

Клеточное строение губок
У асконоидных губок стенка самая тонкая, у лейконоидных наиболее толстая. Но качественно стенка отличается только у стеклянных губок. Она состоит не из отдельных клеток, как у всех остальных, а представляет собой многоядерную цитоплазматическую массу, ограниченную поверхностной мембраной (синцитий). У обыкновенных и известковых губок стенка состоит из клеток, образующих ложные ткани двух разных типов: «соединительную» и «эпителиальную». «Эпителиальная ткань» чаще лишена характерных для эпителия межклеточных контактов и не подстилается базальной пластинкой.
У губок «эпителиальные ткани» — это:
- пинакодерма, она выстилает отводящие и приводящие каналы, покрывает наружную поверхность тела;
- жгутиковая хоанодерма, покрывающая атриум у асконоидных губок и жгутиковые камеры у лейконоидных и сиконоидных.

- Пинакодерма состоит из отдельных клеток двух типов:
- пинакоцитов. Они уплощённые, похожие на чешуйки, плотно пригнанные друг к другу. Как правило они лишены жгутиков. Только у губок родов Plakina и Oscarella каналы выстланы жгутиковыми эндопинакоцитами;
- пороцитов — образуют остии у всех асконоидных и многих других губок. А также апопили и прозопили многих лейконоидных и сиконоидных губок. Но у многих остии могут быть просто щелями между соседними пинакоцитами, а прозопили — между хоаноцитами. Пороциты могут сокращаться при помощи цитоплазматических волокон, поэтому отверстия представляют собой клапаны организма, работающие по типу сфинктеров.
2. Хоанодерма состоит из хоаноцитов — одножгутиковых воротничковых клеток. По строению они похожи на хоанофлагеллят.
3. Между пинакодермой и хоанодермой лежит мезохил — слой, похожий на соединительную ткань. Особенно хорошо он выражен у туалетных губок, представленный густой волокнистой сетью.
Мезохил — единственный слой тела губок, который не омывается водой, да и вообще он слоем не считается (помните, что губки двухслойны?). Он состоит из совокупность межклеточного матрикса и населяющих его разнообразных клеток. Среди его многочисленных клеток есть:
- похожие на макрофаги археоциты. Они крупные, с большим ядром и крупными лизосомами, способные к амёбоидному движению. Археоциты могут превращаться в любые другие клетки, встречающиеся у губок;
- лофоциты — другие амёбоидные клетки, внешне похожие на археоциты, но при движении по мезохилу они оставляют за собой нити коллагена. Их функция — пополнение коллагеновых волокон тела губки;
- спонгоциты — есть только у Demospongiae, они также внешне похожи на археоциты. Но выделяют коллаген, который, застывая, превращается в спонгин в виде толстых скелетных волокон;
- склероциты формируют у многих губок минеральный скелет;
- миоциты напоминают мышечные клетки, содержат актин и миозин и способны к слабому сокращению. У некоторых обыкновенных губок они в большом количестве скапливаются вокруг оскулумов, регулируя их диаметр;
- ооциты и сперматоциты — предшественники половых клеток, тоже расположены в мезохиле. Там они претерпевают гаметогенез и дают начало яйцеклеткам и сперматозоидам.

Скелет
Губки подвергаются натиску течения воды, и чтобы ему противостоять, они обзавелись скелетными элементами. Они бывают жёсткими и хрупкими или эластичными как резина — минеральными и органическими (из спонгина). Чаще они расположены в мезохиле (эндоскелет), но есть виды с наружным скелетными элементами (экзоскелет), покрывающими либо всё тело губки, либо только отдельные его участки. Жёсткость скелета сильно отличается даже у губок одного вида, она зависит от формы их тела.

Скелетом мягких корковых форм, таких, как Oscarella и Halisarca, является студенистый мезохил, пронизанный лишь тонкими коллагеновыми волокнами. Чаще всего мезохил бывает укреплённым минеральными спикулами или спонгином или тем и другим одновременно. Концы спикул могут выходит наружу из пинакодермы и осуществлять дополнительную механическую защиту губки.
Скелет из спикул может представлять собой жёсткую хрупкую решётку или каркас, как у стеклянной губки Euplectella aspergillum или известковой Minchinella sp. Коралловые губки Demospongia имеют массивный экзоскелет из СаСО3, дополняемый спикулами из кремнезёма, как и у большинства обыкновенных губок.
У некоторых губок нет спикул, зато есть органический спонгин. Их тело пористое и эластичное, легко сжимается. Большое количество спонгина делает тело похожим на резину. Иногда в спонгине заключаются инородные частицы, например песчинки. У большинства губок присутствует и спонгин и спикулы.
Спикулы — это элементы скелета, построенные из кремнёзема (SiО2) или углекислого кальция (СаСО3). Их состав, размер и форма используются в систематике губок на всех таксономических уровнях. Они бывают крупными и мелкими, в форме игл с прямым и закруглённым концом и трёхлучевыми.

Фильтрация
По способу питания и по образу жизни все губки относятся к организмам-биофильтраторам. Ток воды не только приносит пищевые частички к клеткам их тела, но и снабжает их кислородом (дышат они путём диффузии), удаляет продукты обмена веществ, выносит личинок и половые клетки. Объём жидкости, который прогоняют через себя губки, поразителен. В среднем каждые 5 сек. губка прокачивает количество воды, равное объёму своего тела.
Морская губка Geodia cydonium весом 1 кг процеживает около 24 тысяч литров воды в сутки, а пресноводная Spongilla lacustris фильтрует воду со скоростью 0,3 мл/с на 1 г сырого веса, что для губки весом 1 кг составляет примерно 25 тысяч литров воды в сутки.
Они могут снижать скорость фильтрации и совсем прекращать поток воды, чтобы, например, предотвратить попадание ила в тело. Осуществляют они это при помощи регуляции диаметра оскулума.
Направленный ток воды создаётся при помощи хоаноцитов. Оскулумы у многих видов губок расположены на вершине и работают по типу «вытяжных труб». У основания губок скорость течения воды ниже, чем рядом с оскулумами. В результате создаётся разность давления и поток устремляется из области высокого давления к области низкого — от остий к оскулумам.
Так как у губок нет органов, то их функции заменяет водоносная система. Но отсутствие дифференцировки тела не означает, что их можно считать низшими, плохо приспособленными к среде обитания. Наоборот они чрезвычайно быстро приспосабливаются к изменениям путём перестроения тела, восстановления его повреждённых участков и размножения бесполым путём.
Питание
У губок внутриклеточное пищеварение. Все их клетки способны поглощать пищу путём фагоцитоза. Из воды, протекающей через их тело, они отфильтровывают частицы размером от 50 до 1 мкм. В эти размеры попадают одноклеточные планктонные организмы, такие, как вирусы, водоросли, грибки, бактерии, динофлагелляты, мелкие частицы детрита и др. Улавливают их постепенно сужающиеся вглубь приводящие каналы и хоаноциты. Далее археоциты, экзо- и эндопинатоциты их фагоцитируют и переваривают в пищеварительных вакуолях.
Есть хищные виды, относящиеся к роду Cladorhizidae. Они ловят мелких животных, типа рачков, которые прилипают к клейким нитям, раскинутым губкой. Пойманное животное подтягивается к стенкам тела губки и переваривается при помощи археоцитов.
У многих губок есть автотрофные симбионты (зоохлореллы, зооксантеллы, цианобактерии), от которых они получают питательные вещества. Некоторые губки с Большого Барьерного рифа (Австралия) за счет симбиотических цианобактерии покрывают от 48 до 80 % своих энергетических потребностей.

Токсичность губок
Многие губки вырабатывают и накапливают на поверхности тела продукты обмена, которые часто бывают токсичными для определённых хищников. Например, 9 из 16 антарктических губок и 27 из 36 карибских губок токсичны для рыб. Но они отпугивают не всех животных. Да и среди рыб встречаются губкоядные виды (рыбы-ангелы и единороговые). Охотно питаются губками морские черепахи, особенно бисса.
Некоторые губки используют вырабатываемые ими вещества в борьбе с конкурентами за пространство. Так, карибский вид Chondrilla nucula таким способом убивает находящиеся поблизости кораллы и после обрастает вокруг их прочных скелетов.

Неприятным сильным запахом обладает «чесночная» губка Lissodendoryx isodictyalis. А прикосновение к карибской «огненной» губке Tedania ignis вызывает сильное раздражение на коже.
Движение губок
Подавляющее большинство губок — прикреплённые животные. Но некоторые из них способны к ограниченному передвижению. Морские (Chondrilla, Hymeniacidon, Tethya) и пресноводные (Ephydatia) перемещаются по субстрату со скоростью от 1 до 4 мм в день. Двигаются они при помощи более или менее согласованного перемещения амёбоидных клеток. Также губки могут сокращаться и регулировать размер оскулума при помощи миоцитов.
Отличительная особенность губок в способности многих их клеток свободно перемещаться. Амёбоидные клетки мезохила двигаются постоянно. Хоаноциты и эндопиканоциты также способны менять своё место расположения в ходе перестройки водоносной системы.
Размножение
Губки размножаются бесполым и половым путём и в жизненном цикле многих из них происходит чередование половой и бесполой стадий. Бесполое размножение осуществляется путём фрагментации, почкования или образования зимующих пропагул (геммул).
- Фрагментация — размножение частями тела. Оторвавшиеся или оставшиеся после поедания хищниками куски губки благодаря способности к регенерации легко выживают и дают начало новому организму. Даже протёртая через мелкоячеистое сито, т.е. практически разделённая на отдельные клетки, губка превращается в целое нормально функционирующее животное.
- Почкование встречается у губок достаточно редко. У асконоидной Clathrina на свободных концах трубок образуются вздутия, там формируются почки, которые отделяются и развиваются в отдельный организм. У некоторых видов Tethya почки образуются на стебельках. Виды родов Oscarella и Aplysilla формируют папиллы, также отделяющиеся и становящиеся самостоятельными дочерними губками.
- Геммулы — клетки, напоминающие споры. Их образуют многие пресноводные и некоторые морские виды. Обычно это происходит осенью. Геммулы впадают в диапаузу и переживают зиму. Весной они «прорастают» и вырастают в целую губку. В теле погибающей губки геммулы формируются в мезохиле из группы археоцитов. Спонгоциты образуют вокруг них спонгиновую оболочку, в которую часто инкрустируются спикулы. Оболочка покрывает клетки полностью, оставляя только отверстие (микропиле) на одном из полюсов.
Половое размножение
Почти все губки (за редким исключением) гермафродиты. В подходящий период все они выбрасывает сперматозоиды, которые током воды переносятся к другой губке, где их захватывают хоаноциты, теряя жгутик и превращаясь в клетку-носителя. Они переносят сперматозоиды к яйцеклеткам. Внутри мезохила происходит слияние сперматозоида с яйцеклеткой (оплодотворение). У некоторых губок зиготы выходят в окружающую среду и развиваются там. У других наблюдается живорождение (личинкорождение), зигота развивается в личинку внутри материнского организма.

Личинки губок весьма разнообразны. И называются они тоже по-разному:
- целобластула (у известковых губок) — пустой внутри шар, состоящий из одного слоя жгутиковых клеток. Личинка ведёт планктонный образ жизни, а многие её клетки теряют жгутики, становятся амёбоидными, переходят внутрь шара, постепенно заполняя его. Заполненная клетками целобластула носит название стерробластулы;
- амфибластула (у губок родов Grantia, Sycon и Leucosotenia) — полый шар из двух типов клеток: на одном конце жгутиковых, на другом безжгутиковых с зернистой цитоплазмой. Формируются они в теле губки;
- паренхимула (у большинства обыкновенных губок) — масса клеток без внутренней полости. Снаружи расположены жгутиковые клетки, между которыми попадаются безжгутиковые везикулярные клетки. Внутри личинки располагаются разные клетки: склероциты, колленциты, пинакоциты, археоциты и даже жгутиковые камеры. Паренхимула — это почти готовая губка, приспособленная для плавания;
- трихимелла (у стеклянных губок) — это почти стерробластула, но с пояском жгутиковых клеток по экватору. Внутри неё есть клетки, содержащие желток, склероциты (со спикулами), другие клетки и жгутиковые камеры.

Личинки всех губок живут недолго. Вскоре они оседают и принимаются ползать по дну в поисках подходящего для прикрепления места. Прикрепляются, претерпевают метаморфоз и становятся молодыми губками.
Значение губок
Как биофильтраторы они выполняют важнейшую роль в очистке воды. Очищают миллионы тонн воды, улучшая санитарное состояние водной среды. Кроме микроорганизмов и жидкой органики, они адсорбируют также тяжёлые металлы и галогены.

Пресноводные губки служат индикаторами загрязнения воды. Их кремниевые спикулы отличаются быстрым реагированием на загрязнения. Многие губки разрушают известковые кораллы и раковины моллюсков.
У острорылого краба (Oregonia gracilis) оригинальный способ маскировки. Он селит на свой панцирь прикреплённых животных, в том числе и губок или растения, которые разрастаясь, скрывают его от хищников.
Многие мелкие животные используют тело губок как убежище. В их водоносных каналах селятся ракообразные и офиуры.

Класс Известковые губки (Calcarea)
- включает 680 видов, объединённых в 5 отрядов (8,2 % от всех описанных видов губок);
- исключительно морские животные, встречающиеся как на литорали, так и батиали;
- могут иметь асконоидный, сиконоидный и лейконоидный типы строения;
- спикулы из кальцита (карбонат кальция) — известковый скелет, чаще сливаются в триаксоны, тетраксоны или монаксоны;
- мезохил представлен тонким слоем;
- хоаноциты крупные, сравнимые по величине с археоцитами и пинакоцитами.

Класс Homoscleromorpha
Относительно молодой таксон, выделенный совсем недавно, ранее представителей класса причисляли к обыкновенным губкам. Их особенности:
- 6 родов, 2 семейства и 1 отряд;
- обитают в морях на небольшой глубине;
- тело гладкое в виде корки или комка;
- строение лейконоидное или силлиибидное;
- спикулы четырёхлучевые с ветвящимися и изгибающимися краями, состоят из кремнезёма;
- клетки хоанодермы и пинакодермы подстилаются базальной пластинкой как у эуметазоев;
- пинакоциты имеют жгутики;
- бесполое размножение при помощи почкования и фрагментации;
- живородящие, личинка — цинктобластула.
Класс Обыкновенные губки (Demospongiae)
- примерно 6900 видов, объединённых в 12 отрядов (83,3 % всех современных губок);
- обитают в пресных и солёных водах;
- тип организации — лейконоидный;
- скелет из кремнезёмных спикул, спонгина или из того и другого вместе, а иногда отсутствует. У некоторых реликтовых видов есть наружный известковый плотный скелет;
- мезохил развит хорошо;
- хоаноциты мельче других клеток;
- представители: оранжево-красные морские апельсины (Tethya), комковидные пробковые губки (сем. Suberitidae), сверлящие губки (сем. Clionidae), туалетные губки, бадяги, байкальские губки и др.

Класс Стеклянные губки (Hexactinellida, Hyalospongia)
- 600 видов (7,3 % от всех описанных губок);
- обитают в морях на значительной глубине (от 5 до 6770 м.);
- тело мешковидное, трубчатое, иногда в виде бокала;
- почти всегда одиночные организмы с сиконоидным типом строения;
- спикулы шестилучевые из кремнезёма. Они спаиваются и образуют прочную, но хрупкую скелетную решётку;
- большая часть клеток объединяется в синцитий;
- мезохил развит слабо.

У некоторых видов рода тело цилиндрическое, до 1 м. в высоту, иглы у основания, втыкающиеся в грунт, достигают 3 метров длины.