[править]
Почему у нейрона один аксон?
<- назад
дальше ->
Действительно, а в чем причина этого? Если природа обошлась единственным каналом вывода нервных сигналов из нейрона, значит его вполне достаточно для совершаемых нервной системой актов. Любой нейрон в нервной системе почему-то всегда имеет только один аксон. Если уж и есть нейроны с несколькими аксонами, то только за счет «склеивания» в процессе развития организма нескольких одноаксонных нейронов в один большой нейрон. На первый взгляд получается, что выходная «шина данных» обычного нейрона состоит из единственного «провода», но поскольку хотя бы даже на малых участках нервной системы все-таки предполагается быть транспортировке существенно емких нервных сигналов, вероятно включающих произвольные «порции» потребных сведений, то в свете алгоритмической интерпретации совершаемых нервной системой актов выглядит неясным вопрос о том, как природой решена проблема «разрядности», «структурности» портируемых на выход нейрона данных. В связи с этим в среде лиц, причастных к проектированию самостоятельных «аналогов» нейронов, обычно возникают два противоборствующих объяснения: аксон или является одноканальной шиной для последовательной передачи ненормированной поточной информации, либо алгоритм работы ядра нейрона основан на строгом акцентировании «типоразмера» обслуживаемых данных, четком структурировании выходящего информационного потока.
Попробуем посильно разобрать каждое объяснение с учетом искусственной аппаратной или же программной реализации природных особенностей работы выходного канала нейрона. Той самой особенности, которую приходится поневоле воображать, моделировать в уме с помощью плохо обоснованных допущений, на основе банальных представлений о передаче информации в компьютерных системах, доверяясь мнимой некоторой общности таких представлений со спецификой передачи оперативной информации в нервных сигналах. Начнем со второго объяснения как знакомого всякому, кто хоть мельком касался темы способов представления данных в компьютерах.
Структурирование данных в принципе и есть предварительное описание содержимого информационного потока, коим может выступать тот гипотетически емкий нервный сигнал, только в обычном случае поля структуры (контейнеры «порций» соответствующих сведений) строго фиксированы по емкости несомой в них информации. Структурированные данные несложно передавать одноканальным потоком, и значительный плюс здесь заключен вот в чем: нет нужды вносить в поток дополнительные информационные элементы, отделяющие поля структуры друг от друга, ведь отправляющий и принимающий связаны единым соглашением о порядке и размере следуемых в потоке «порций» данных. Касаемо внутреннего устройства нейрона это означает, что последний в таком случае избавлен от необходимости иметь в своем составе аппаратные или программные компоненты, следящие за разделительными признаками во входном потоке или же, наоборот, формирующие такие признаки в выходном потоке нейрона.
Но при подобных условиях искусственный нейрон не в состоянии передавать или принимать за один «такт» деятельности информации больше или меньше, чем ограничено размером выбранной структуры. Поскольку нейроны в организме конструируются на одной и той же «элементной» базе, а также существует великое разнообразие транспортируемых через них типов информации, то предшествующая проектировка полей структуры информационного потока должна удовлетворять требованиям всех мыслимых нервных узлов и центров, что кажется сложным осуществить одной структурой с запрограммированным порядком и размером полей.
Более выгодной видится поддержка потоков с ненормированным порядком и размером полей, то есть безтиповое структурирование внедрением в поток специальных отметок, идентификаторов, определяющих не только концы «порций» сведений, но также их именные, классификационные приметы. Возможно и даже наиболее вероятно, что именно так или по схожей идее организована методика передачи вместительных нервных сигналов в настоящих нейронах. Подобный поток легко обрабатывать порциями, легко пропускать при необходимости его участки, легко вообще отказаться от обработки в некоторой точке потоке. Заманчивая идея: обрабатывать поток, ориентируясь на какие-то признаки внутри него. Вот только непонятно, как организовать обратную связь с отправителем потока. Как ему сообщить, что нейрон решил прервать обработку потока и отправитель должен прекратить передачу?
Конечно, первое из противоборствующих объяснений привлекает широтой возможностей, в то же время оно вносит в имитацию нейронов дополнительные трудности, обусловленные спецификой обслуживания ненормированных потоков: это и аппаратные (программные) устройства обратной связи; это и устройства отслеживания начала и конца передачи, внедренных идентификаторов и тому подобных вещей. В случае поддержки обратной связи в поток придется внедрить еще и код получателя, ибо у нейрона аксон один, а сообщить получателю свою резолюцию по обработке идущего от него потока нейрон может только с помощью своего выхода. К нему подключены и другие нейроны, ждущие своего потока данных, и в этот момент по выходной шине начинают идти сигналы какому-то неведомому им отправителю. Следовательно, у нейронов должны быть еще и устройства, выделяющие из нервного сигнала код получателя, чтобы по ошибке не отзываться на чужой сигнал.
Вообще кажется странным, почему природа создала один аксон. Что это — экономия, необходимость или неведомая технология обмена нервными сигналами? Экономя на одном аксоне как на последовательной шине ненормированных поточных данных, природа неизбежно вышла бы на весомые затраты в дендритах. Сводя предположительно многоканальную внутреннюю шину данных нейрона в одноканальный выходной сигнал, природа обязана была бы позаботиться, чтобы каждый дендрит нейрона имел аппаратные устройства по «расканаливанию» и введению поточных данных входного сигнала во внутреннюю шину данных. Крупные нейроны головного мозга имеют десятки тысяч дендритов. Выходят неимоверные аппаратные затраты, если необходимо каждый дендрит снабдить таким вот «разводящим» устройством, так сказать, демультиплексором (мультиплексор — микросхема с несколькими входами и одним выходом; управляющими сигналами осуществляется подача на выход данных одного из входных каналов). В этих аналогиях работа аксона напоминает мультиплексирование внутренней шины данных с отсроченным попаданием всех предложенных данных в выходной поток, работа дендрита — демультиплексирование.
Понятно, насколько выгодным в транспортном смысле оказывается одноканальный выход с ненормированным информационным потоком. Да только в программной имитации, в соответствии с доступными сегодня пропускными и вычислительными мощностями, целесообразным пока оказывается склонение к фиксированной размерной структуризации выходного потока. Это прямо повышает производительность запрограммированной нейронной схемы за счет подведения под работу ядра нейрона требования выводить на выход исключительно точные и краткие данные, укладывающиеся в заблаговременно описанную и единую для всех искусственных нейронов структуру информационного потока. Во-первых, это искусственно ограничивает использование памяти эмулирующего устройства (например компьютера), во-вторых, освобождает программные модели нейронов от затратных по времени функций мультиплексирования и демультиплексирования сигналов в аксонах и дендритах. Минус заключен в том, что помимо потери некоторых алгоритмических путей обработки нет никакой гарантии, что избранная статическая структура информационного потока окажется подходящей для большинства целевых нейронных схем.
Дмитрий Сахань, 7 декабря 2002 года
[править]
Справка по разделу -Теория по нейронным микросхемам
Этот неподвластный интеллект
Нейронные микросхемы
Нейронные микросхемы с обратными функциями
Заметки для экспериментов с нейронными микросхемами
Почему у нейрона один аксон?
Каков «алгоритм» работы рецептора?
Следствия в нейронах вместо данных
Каковы настоящие функции нейрона?
Препарируя нейрон
К вопросу о скорости работы нейронов
Рободелам на заметку
Рободелам на заметку 2
Как нейрон фильтрует сигналы
Идея: ядро нейрона помнит правильно выполненное следствие
А не работает ли нейрон по-другому?
К вопросу о механизме обучения нейронов
[править]
Искусственный интеллект в домашних условиях Дмитрий Сахань
I think during the early evolution of nervous systems it was very important to have long axon cells. For example a worm has 3 giant axons which are almost the same length than the worm itself. So there was selection pressure on the length of the axon. The neuron requires a lot of energy to send signals to other neurons. Neurons use energy on nanowatt scale, average human cells use energy on picowatt scale… By long axons it requires less energy to send the message through a single axon than send it through multiple axons. That’s the same why we use limited amount of undersea internet cables — currently about 200. If there is more data than a single cable can handle, only then lay we a parallel cable. So a possible answer is evolution of worm nervous systems combined with energy consumption optimization.
Another answer can be that neurons would be never capable to build multiple action potentials parallel. Maybe they cannot consume enough energy to hold charges separated by multiple axons. I don’t intend to look for the details of that. It is technically possible to have neurons with multiple axons, so you can grow things like that, but it is not clear if their axons can work parallel or not, and if they don’t work parallel does they interfere the communication of each other.
About single axon neural networks there are existing mathematical models. Until there is no mathematical model how a neural network with multiple axons would work, or there is no evidence of it is not possible to build such a thing, you cannot say for sure that it is because of the mathematics of neural networks.
So I have this 3 theories. I think you can find more about this topic by googling a lot, and making new experiments. For me it was enough. 
Глава 10. Нервная ткань
Нервная
ткань высокоспециализированная, из нее
построена вся нервная система. В
центральной нервной системе она образует
серое и белое вещест во головного и
спинного мозга, в периферической—
ганглии, нервы, нервные окончания.
Нервная ткань способна воспринимать
раздражения из внешней и внутренней
среды, возбуждаться под их влиянием,
вырабатывать, проводить
ипередавать
импульсы, организовывать ответные
реакции. Сумма этих свойств нервной
ткани проявляется в основной функции
нервной системы: регуляции и координации
деятельности различных тканей, органов
и систем организма.
Развивается
нервная ткань из нейроэктодермы. Из нее
образуется сначаланервная пластинка,
а затемнервная трубка, вдоль которой с
двух сторон лежатнервные гребни (валики).
В нервной трубке и гребнях формируются
все клетки нервной ткани. Строение
нервной ткани в различных участках
нервной системы сильно различается.
Тем не менее она везде состоит изнейронов
инейроглии. Между ними имеются межклеточные
пространства, заполненные тканевой
жидкостью. Межклеточные пространства
мозга составляют15—20%его объема. В
тканевой жидкости происходит диффузия
веществ между капиллярами и клетками
нервной ткани. Нейроны — нервные клетки,
способные к выработке и проведению
нервного импульса. Нейроглия состоит
из клеток, выполняющих вспомогательные
функции.
Вопрос 66
Нейроглия
заполняет в нервной ткани все пространства
между нейронами, их отростками,
кровеносными капиллярами. Тесно прилегает
к перечисленным структурам, образуя их
оболочки. Она выполняет разнообразные
функции: опорную, изолирующую,
разграничительную, трофическую, защитную,
обменную, гомеостатическую. Нейроглиальные
клетки — глиоциты
— называют
вспомогательными клетками нервной
ткани, так как они не проводят нервный
импульс. Тем не менее их функции жизненно
необходимы, поскольку отсутствие или
повреждение нейроглии делает невозможной
работу нейронов. Существуют две
разновидности нейроглии: макроглия и
микроглия.
Макроглия
(глиоциты), как и нейроны, развивается
из клеток нервной трубки. Среди глиоцитов
различают: эпендимоциты, астроциты,
олигодендроциты.
Эпендимоциты
— глиальные клетки кубической или
цилиндрической формы, на их апикальном
полюсе имеются реснички, от базального
полюса отходит длинный отросток, который
пронизывает всю толщу мозга. Они плотно
прилегают друг к другу, выстилая сплошным
слоем стенки желудочков мозга и
спинномозгового канала. Движениями
ресничек создается ток цереброспинальной
жидкости. В некоторых эпендимоцитах
обнаруживают секреторные гранулы.
Предполагают, что эпендимоциты выделяют
секрет в цереброспинальную жидкость и
регулируют ее состав.
Вопрос 67
Строение
и виды нейронов. Нейрон (нейроцит) —
основная структурная
ифункциональная
единица нервной ткани (рис. 32). В нем
различают тело
— перикарион
и отростки. Нейроны разных отделов
нервной системы отличаются друг от
друга по функции, форме, размерам,
количеству и характеру ветвления
отростков, по выделяемому медиатору.
По функции нейроны бывают чувствительные
(рецепторные, или афферентные),двигательные
(эффекторные, или эфферентные) и вставочные
(ассоциативные). Размеры нейроцитов
колеблются от 4 мкм уклеток-зеренмозжечка
до 130 мкм (у гигантопирамидных клеток
коры)
. Нейроны
в основном одноядерные клетки. Ядро
крупное, округлое, расположено обычно
в центре клетки. Кариоплазма светлая,
так как хроматин не образует крупных
глыбок. Содержит 1—2крупных ядрышка.
Комплекс Гольджи расположен вокруг
ядра. Много митохондрий, микротрубочек,
есть центросома, лизосомы. Хорошо
представлен аппарат синтеза белка:
рибосомы и гранулярная цитоплазматическая
сеть. Адсорбция основных красителей на
скоплениях этих органелл образует
характерную картину в виде крупных
глыбок, напоминающую шкуру тигра (при
изучении в световой микроскоп),
за что и
названа тигроидным(базофильным) веществом
или субстанцией Ниссля(по имени описавшего
ее гистолога). Есть и специальные
орга-неллы
—
нейрофиламенты. Пучки нейрофиламентов
и микротрубочек (нейротубул) благодаря
адсорбции на них красителей видны в
световой микроскоп в виденейрофибрилл.
Эти органеллы участвуют в формировании
цитоокелета, в передвижении веществ по
клетке и ее отросткам.
Форма
перикариона во многом определяется
количеством отростков. Различают
униполярные — с одним отростком,
ложно-униполярные,биполярные — с двумя
отростками и мультиполярные нейроны —
с несколькими(3—20)отростками. Тела
униполярных и ложноуниполярных клеток
округлые, биполярных — веретеновидные,
мультиполярных — разнообразные. Отростки
— обязательная принадлежность нейронов.
Без них нейроциты не могут выполнять
свои функции, так как отростки обеспечивают
проведение нервного импульса от одной
части тела в другую. Их длина бывает от
нескольких микрометров до1—2м. По
морфологическим и функциональным
свойствам отростки неравнозначны. В
нейроне различаютдендриты иаксон
(нейрит). Аксон в клетке всегда один,
дендритов может быть разное количество.
По аксону возбуждение распространяется
от тела, по дендриту— к телу нервной
клетки. Дендриты, как правило, сильно
ветвятся и в них присутствуют все
органеллы, которые есть и в теле клетки.
Аксон не ветвится, но может отдавать
коллатерали — ответвления, идущие
параллельно. В нем нет базофильного
вещества. Нейрофиламенты и нейротрубочки
располагаются упорядочение — вдоль
аксона. Униполярными считаются
недифференцированные нервные клетки
на ранней стадии развития, когда дендриты
еще не образовались. Среди дифференцированных
клеток униполярные нейроны встречаются
редко.
От тела
ложноуниполярного нейрона отходит один
отросток, который Т- образно разветвляется
на дендрит и нейрит. Такие клетки
распространены в спинномозговых узлах
(ганглиях). Это чувствительные нейроны,
дендриты которых идут на периферию, где
заканчиваются в органах чувствительными
нервными окончаниями (рецепторами), а
нейриты несут возбуждение от тела клетки
в центральную нервную систему. Как
видим, эти клетки по своим
структурно-функциональнымсвойствам
приближаются кбиполярным нейронам,
которые встречаются в органе зрения,
обоняния и среди ассоциативных нейронов.
Самыми распространенными
являютсямультиполярные нейроны. Это
все двигательные (моторные) и большинство
ассоциативных нейронов. Среди их
отростков только один аксон, а остальные
дендриты. У ассоциативных нейронов
аксон не покидает центральной нервной
системы, у двигательных — идет на
периферию—корганам (мышцам, железам),
где и оканчивается двигательным нервным
окончанием.
Нервные
клетки рано дифференцируются в онтогенезе,
утрачивают способность к делению, в
норме продолжительность их жизни равна
продолжительности жизни особи. Для
поддержания жизнедеятельности и
способности к выполнению функций на
протяжении столь длительного времени
в нейронах развита система внутриклеточной
регенерации. При этом макромолекулы и
их ансамбли постоянно разрушаются и
создаются вновь. Белковые синтезы идут
в основном в теле клетки. Высокий уровень
жизнедеятельности отростков поддерживается
постоянным таком цитоплазмы в отростки
и обратно.
Плазмолемма
нейрона выполняет все функции, присущие
ей в любых клетках. Кроме того, она
способна к возбуждению при деполяризации
(снижении величины заряда) в результате
перемещения ионов Na+ в клетку. Деполяризация
возникает локально (в одном месте) и
волнообразно перемещается от дендрита
к телу и аксону. С какой скоростью
движется волна деполяризации, с такой
же скоростью передается и нервный
импульс. Торможение наступает при
противоположном явлении: увеличении
заряда мембраны под влиянием ионных
потоков (О-—в клетку и К+ — из клетки).
В нервной ткани нейроны образуют
ансамбли, характерные для определенных
участков нервной системы. Характер их
расположения носит названиецитоархи-
тектоника.
Передача
нервного импульса от одного нейрона к
другому осуществляется в месте их
контакта—синапсе(sinapsis — соединение)
(рис. 33). В зависимости от того, какие
участки нейронов вступают в контакт,
различаютаксодендритические (аксон
одного нейрона контактирует с дендритом
другого нейрона),аксосоматические
(аксон контактирует с телом другого
нейрона) и аксоаксо-нальные (контактируют
аксоны двух нейронов) синапсы. Описаны
такжедендросоматические идендродендритные
синапсы. Примерно поло-
вина
поверхности тела нейрона и почти вся
поверхность его дендритов бывает занята
синапсами.
В результате
каждый нейрон имеет обширные контакты.
Так, на одной грушевидной клетке мозжечка
насчитывают до 200 000 синапсов. Синапсы
бывают как возбудительные, так и
тормозные.
У всех
синапсов общие принципы строения:
концевые веточки аксона, передающего
импульс нейрона в месте синапса, образуют
колбовидные утолщения — это пресинаптический
полюс. В нем содержится много митохондрий
исинаптических пузырьков, которые
различаются по виду и размерам в
зависимости от содержащегося в
нихмедиатора — вещества, возбуждающего
второй нейрон. Это могут быть серотонин,
ацетилхоллин, адреналин и другие
вещества. Участок второго нейрона,
воспринимающий импульс,
называетсяпостсинаптическим полюсом.
В нем нет синаптических пузырьков и
митохондрий. Между двумя полюсами
находится узкаясинап-
тическая
щель (около 20 им), ограниченная
контактирующими мембранамидвух полюсов:
пресинаптическойи постсинаптической.Эти
мембраны имеют утолщения и другие
специальные структурные приспособления,
обеспечивающие успешную передачу
нервного импульса только в одном
направлении. Нервный импульс, пришедший
в пресинаптический полюс, приводит к
выбросу медиатора в синаптическую щель.
Вызванный им нервныйим- пульс переходит
на второй нейрон.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
I think during the early evolution of nervous systems it was very important to have long axon cells. For example a worm has 3 giant axons which are almost the same length than the worm itself. So there was selection pressure on the length of the axon. The neuron requires a lot of energy to send signals to other neurons. Neurons use energy on nanowatt scale, average human cells use energy on picowatt scale… By long axons it requires less energy to send the message through a single axon than send it through multiple axons. That’s the same why we use limited amount of undersea internet cables — currently about 200. If there is more data than a single cable can handle, only then lay we a parallel cable. So a possible answer is evolution of worm nervous systems combined with energy consumption optimization.
Another answer can be that neurons would be never capable to build multiple action potentials parallel. Maybe they cannot consume enough energy to hold charges separated by multiple axons. I don’t intend to look for the details of that. It is technically possible to have neurons with multiple axons, so you can grow things like that, but it is not clear if their axons can work parallel or not, and if they don’t work parallel does they interfere the communication of each other.
About single axon neural networks there are existing mathematical models. Until there is no mathematical model how a neural network with multiple axons would work, or there is no evidence of it is not possible to build such a thing, you cannot say for sure that it is because of the mathematics of neural networks.
So I have this 3 theories. I think you can find more about this topic by googling a lot, and making new experiments. For me it was enough.
Not to be confused with Neutron.
| Neuron | |
|---|---|

Anatomy of a multipolar neuron |
|
| Identifiers | |
| MeSH | D009474 |
| NeuroLex ID | sao1417703748 |
| TA98 | A14.0.00.002 |
| TH | H2.00.06.1.00002 |
| FMA | 54527 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A neuron, neurone, or nerve cell is an electrically excitable cell that fires electric signals called action potentials. Neurons communicate with other cells via synapses — specialized connections that commonly use minute amounts of chemical neurotransmitters to pass the electric signal from the presynaptic neuron to the target cell through the synaptic gap. The neuron is the main component of nervous tissue in all animals except sponges and placozoa. Non-animals like plants and fungi do not have nerve cells.
Neurons are typically classified into three types based on their function. Sensory neurons respond to stimuli such as touch, sound, or light that affect the cells of the sensory organs, and they send signals to the spinal cord or brain. Motor neurons receive signals from the brain and spinal cord to control everything from muscle contractions to glandular output. Interneurons connect neurons to other neurons within the same region of the brain or spinal cord. When multiple neurons are functionally connected together, they form what is called a neural circuit.
A typical neuron consists of a cell body (soma), dendrites, and a single axon. The soma is a compact structure, and the axon and dendrites are filaments extruding from the soma. Dendrites typically branch profusely and extend a few hundred micrometers from the soma. The axon leaves the soma at a swelling called the axon hillock and travels for as far as 1 meter in humans or more in other species. It branches but usually maintains a constant diameter. At the farthest tip of the axon’s branches are axon terminals, where the neuron can transmit a signal across the synapse to another cell. Neurons may lack dendrites or have no axon. The term neurite is used to describe either a dendrite or an axon, particularly when the cell is undifferentiated.
Most neurons receive signals via the dendrites and soma and send out signals down the axon. At the majority of synapses, signals cross from the axon of one neuron to a dendrite of another. However, synapses can connect an axon to another axon or a dendrite to another dendrite.
The signaling process is partly electrical and partly chemical. Neurons are electrically excitable, due to maintenance of voltage gradients across their membranes. If the voltage changes by a large enough amount over a short interval, the neuron generates an all-or-nothing electrochemical pulse called an action potential. This potential travels rapidly along the axon and activates synaptic connections as it reaches them. Synaptic signals may be excitatory or inhibitory, increasing or reducing the net voltage that reaches the soma.
In most cases, neurons are generated by neural stem cells during brain development and childhood. Neurogenesis largely ceases during adulthood in most areas of the brain.
Nervous system[edit]
Schematic of an anatomically accurate single pyramidal neuron, the primaryious excitatory neuron of cerebral cortex, with a synaptic connection from an incoming axon onto a dendritic spine
Neurons are the primary components of the nervous system, along with the glial cells that give them structural and metabolic support.[1] The nervous system is made up of the central nervous system, which includes the brain and spinal cord, and the peripheral nervous system, which includes the autonomic and somatic nervous systems.[2] In vertebrates, the majority of neurons belong to the central nervous system, but some reside in peripheral ganglia, and many sensory neurons are situated in sensory organs such as the retina and cochlea.
Axons may bundle into fascicles that make up the nerves in the peripheral nervous system (like strands of wire make up cables). Bundles of axons in the central nervous system are called tracts.
Anatomy and histology[edit]

Diagram of the components of a neuron
Neurons are highly specialized for the processing and transmission of cellular signals. Given their diversity of functions performed in different parts of the nervous system, there is a wide variety in their shape, size, and electrochemical properties. For instance, the soma of a neuron can vary from 4 to 100 micrometers in diameter.[3]
- The soma is the body of the neuron. As it contains the nucleus, most protein synthesis occurs here. The nucleus can range from 3 to 18 micrometers in diameter.[4]
- The dendrites of a neuron are cellular extensions with many branches. This overall shape and structure are referred to metaphorically as a dendritic tree. This is where the majority of input to the neuron occurs via the dendritic spine.
- The axon is a finer, cable-like projection that can extend tens, hundreds, or even tens of thousands of times the diameter of the soma in length. The axon primarily carries nerve signals away from the soma and carries some types of information back to it. Many neurons have only one axon, but this axon may—and usually will—undergo extensive branching, enabling communication with many target cells. The part of the axon where it emerges from the soma is called the axon hillock. Besides being an anatomical structure, the axon hillock also has the greatest density of voltage-dependent sodium channels. This makes it the most easily excited part of the neuron and the spike initiation zone for the axon. In electrophysiological terms, it has the most negative threshold potential.
- While the axon and axon hillock are generally involved in information outflow, this region can also receive input from other neurons.
- The axon terminal is found at the end of the axon farthest from the soma and contains synapses. Synaptic boutons are specialized structures where neurotransmitter chemicals are released to communicate with target neurons. In addition to synaptic boutons at the axon terminal, a neuron may have en passant boutons, which are located along the length of the axon.
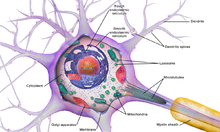
The accepted view of the neuron attributes dedicated functions to its various anatomical components; however, dendrites and axons often act in ways contrary to their so-called main function.[5]
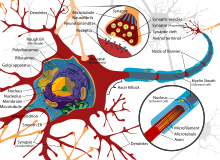
Diagram of a typical myelinated vertebrate motor neuron
Axons and dendrites in the central nervous system are typically only about one micrometer thick, while some in the peripheral nervous system are much thicker. The soma is usually about 10–25 micrometers in diameter and often is not much larger than the cell nucleus it contains. The longest axon of a human motor neuron can be over a meter long, reaching from the base of the spine to the toes.
Sensory neurons can have axons that run from the toes to the posterior column of the spinal cord, over 1.5 meters in adults. Giraffes have single axons several meters in length running along the entire length of their necks. Much of what is known about axonal function comes from studying the squid giant axon, an ideal experimental preparation because of its relatively immense size (0.5–1 millimeter thick, several centimeters long).
Fully differentiated neurons are permanently postmitotic[6] however, stem cells present in the adult brain may regenerate functional neurons throughout the life of an organism (see neurogenesis). Astrocytes are star-shaped glial cells. They have been observed to turn into neurons by virtue of their stem cell-like characteristic of pluripotency.
Membrane[edit]
Like all animal cells, the cell body of every neuron is enclosed by a plasma membrane, a bilayer of lipid molecules with many types of protein structures embedded in it.[7] A lipid bilayer is a powerful electrical insulator, but in neurons, many of the protein structures embedded in the membrane are electrically active. These include ion channels that permit electrically charged ions to flow across the membrane and ion pumps that chemically transport ions from one side of the membrane to the other. Most ion channels are permeable only to specific types of ions. Some ion channels are voltage gated, meaning that they can be switched between open and closed states by altering the voltage difference across the membrane. Others are chemically gated, meaning that they can be switched between open and closed states by interactions with chemicals that diffuse through the extracellular fluid. The ion materials include sodium, potassium, chloride, and calcium. The interactions between ion channels and ion pumps produce a voltage difference across the membrane, typically a bit less than 1/10 of a volt at baseline. This voltage has two functions: first, it provides a power source for an assortment of voltage-dependent protein machinery that is embedded in the membrane; second, it provides a basis for electrical signal transmission between different parts of the membrane.
Histology and internal structure[edit]

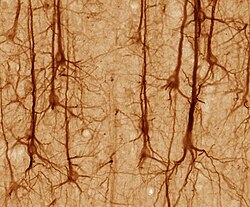
Golgi-stained neurons in human hippocampal tissue

Actin filaments in a mouse cortical neuron in culture
Numerous microscopic clumps called Nissl bodies (or Nissl substance) are seen when nerve cell bodies are stained with a basophilic («base-loving») dye. These structures consist of rough endoplasmic reticulum and associated ribosomal RNA. Named after German psychiatrist and neuropathologist Franz Nissl (1860–1919), they are involved in protein synthesis and their prominence can be explained by the fact that nerve cells are very metabolically active. Basophilic dyes such as aniline or (weakly) haematoxylin[8] highlight negatively charged components, and so bind to the phosphate backbone of the ribosomal RNA.
The cell body of a neuron is supported by a complex mesh of structural proteins called neurofilaments, which together with neurotubules (neuronal microtubules) are assembled into larger neurofibrils.[9] Some neurons also contain pigment granules, such as neuromelanin (a brownish-black pigment that is byproduct of synthesis of catecholamines), and lipofuscin (a yellowish-brown pigment), both of which accumulate with age.[10][11][12] Other structural proteins that are important for neuronal function are actin and the tubulin of microtubules. Class III β-tubulin is found almost exclusively in neurons. Actin is predominately found at the tips of axons and dendrites during neuronal development. There the actin dynamics can be modulated via an interplay with microtubule.[13]
There are different internal structural characteristics between axons and dendrites. Typical axons almost never contain ribosomes, except some in the initial segment. Dendrites contain granular endoplasmic reticulum or ribosomes, in diminishing amounts as the distance from the cell body increases.
Classification[edit]

Neurons vary in shape and size and can be classified by their morphology and function.[15] The anatomist Camillo Golgi grouped neurons into two types; type I with long axons used to move signals over long distances and type II with short axons, which can often be confused with dendrites. Type I cells can be further classified by the location of the soma. The basic morphology of type I neurons, represented by spinal motor neurons, consists of a cell body called the soma and a long thin axon covered by a myelin sheath. The dendritic tree wraps around the cell body and receives signals from other neurons. The end of the axon has branching axon terminals that release neurotransmitters into a gap called the synaptic cleft between the terminals and the dendrites of the next neuron.[citation needed]
Structural classification[edit]
Polarity[edit]
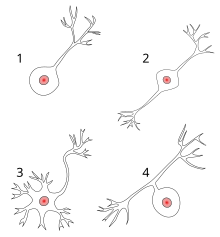
Most neurons can be anatomically characterized as:[citation needed]
- Unipolar: single process
- Bipolar: 1 axon and 1 dendrite
- Multipolar: 1 axon and 2 or more dendrites
- Golgi I: neurons with long-projecting axonal processes; examples are pyramidal cells, Purkinje cells, and anterior horn cells
- Golgi II: neurons whose axonal process projects locally; the best example is the granule cell
- Anaxonic: where the axon cannot be distinguished from the dendrite(s)
- Pseudounipolar: 1 process which then serves as both an axon and a dendrite
Other[edit]
Some unique neuronal types can be identified according to their location in the nervous system and distinct shape. Some examples are:[citation needed]
- Basket cells, interneurons that form a dense plexus of terminals around the soma of target cells, found in the cortex and cerebellum
- Betz cells, large motor neurons
- Lugaro cells, interneurons of the cerebellum
- Medium spiny neurons, most neurons in the corpus striatum
- Purkinje cells, huge neurons in the cerebellum, a type of Golgi I multipolar neuron
- Pyramidal cells, neurons with triangular soma, a type of Golgi I
- Renshaw cells, neurons with both ends linked to alpha motor neurons
- Unipolar brush cells, interneurons with unique dendrite ending in a brush-like tuft
- Granule cells, a type of Golgi II neuron
- Anterior horn cells, motoneurons located in the spinal cord
- Spindle cells, interneurons that connect widely separated areas of the brain
Functional classification[edit]
Direction[edit]
- Afferent neurons convey information from tissues and organs into the central nervous system and are also called sensory neurons.
- Efferent neurons (motor neurons) transmit signals from the central nervous system to the effector cells.
- Interneurons connect neurons within specific regions of the central nervous system.
Afferent and efferent also refer generally to neurons that, respectively, bring information to or send information from the brain.
Action on other neurons[edit]
A neuron affects other neurons by releasing a neurotransmitter that binds to chemical receptors. The effect upon the postsynaptic neuron is determined by the type of receptor that is activated, not by the presynaptic neuron or by the neurotransmitter. A neurotransmitter can be thought of as a key, and a receptor as a lock: the same neurotransmitter can activate multiple types of receptors. Receptors can be classified broadly as excitatory (causing an increase in firing rate), inhibitory (causing a decrease in firing rate), or modulatory (causing long-lasting effects not directly related to firing rate).[citation needed]
The two most common (90%+) neurotransmitters in the brain, glutamate and GABA, have largely consistent actions. Glutamate acts on several types of receptors, and has effects that are excitatory at ionotropic receptors and a modulatory effect at metabotropic receptors. Similarly, GABA acts on several types of receptors, but all of them have inhibitory effects (in adult animals, at least). Because of this consistency, it is common for neuroscientists to refer to cells that release glutamate as «excitatory neurons», and cells that release GABA as «inhibitory neurons». Some other types of neurons have consistent effects, for example, «excitatory» motor neurons in the spinal cord that release acetylcholine, and «inhibitory» spinal neurons that release glycine.[citation needed]
The distinction between excitatory and inhibitory neurotransmitters is not absolute. Rather, it depends on the class of chemical receptors present on the postsynaptic neuron. In principle, a single neuron, releasing a single neurotransmitter, can have excitatory effects on some targets, inhibitory effects on others, and modulatory effects on others still. For example, photoreceptor cells in the retina constantly release the neurotransmitter glutamate in the absence of light. So-called OFF bipolar cells are, like most neurons, excited by the released glutamate. However, neighboring target neurons called ON bipolar cells are instead inhibited by glutamate, because they lack typical ionotropic glutamate receptors and instead express a class of inhibitory metabotropic glutamate receptors.[16] When light is present, the photoreceptors cease releasing glutamate, which relieves the ON bipolar cells from inhibition, activating them; this simultaneously removes the excitation from the OFF bipolar cells, silencing them.[citation needed]
It is possible to identify the type of inhibitory effect a presynaptic neuron will have on a postsynaptic neuron, based on the proteins the presynaptic neuron expresses. Parvalbumin-expressing neurons typically dampen the output signal of the postsynaptic neuron in the visual cortex, whereas somatostatin-expressing neurons typically block dendritic inputs to the postsynaptic neuron.[17]
Discharge patterns[edit]
Neurons have intrinsic electroresponsive properties like intrinsic transmembrane voltage oscillatory patterns.[18] So neurons can be classified according to their electrophysiological characteristics:
- Tonic or regular spiking. Some neurons are typically constantly (tonically) active, typically firing at a constant frequency. Example: interneurons in neurostriatum.
- Phasic or bursting. Neurons that fire in bursts are called phasic.
- Fast spiking. Some neurons are notable for their high firing rates, for example some types of cortical inhibitory interneurons, cells in globus pallidus, retinal ganglion cells.[19][20]
Neurotransmitter[edit]
![]()
Synaptic vesicles containing neurotransmitters
Neurotransmitters are chemical messengers passed from one neuron to another neuron or to a muscle cell or gland cell.
- Cholinergic neurons – acetylcholine. Acetylcholine is released from presynaptic neurons into the synaptic cleft. It acts as a ligand for both ligand-gated ion channels and metabotropic (GPCRs) muscarinic receptors. Nicotinic receptors are pentameric ligand-gated ion channels composed of alpha and beta subunits that bind nicotine. Ligand binding opens the channel causing influx of Na+ depolarization and increases the probability of presynaptic neurotransmitter release. Acetylcholine is synthesized from choline and acetyl coenzyme A.
- Adrenergic neurons – noradrenaline. Noradrenaline (norepinephrine) is released from most postganglionic neurons in the sympathetic nervous system onto two sets of GPCRs: alpha adrenoceptors and beta adrenoceptors. Noradrenaline is one of the three common catecholamine neurotransmitter, and the most prevalent of them in the peripheral nervous system; as with other catecholamines, it is synthesised from tyrosine.
- GABAergic neurons – gamma aminobutyric acid. GABA is one of two neuroinhibitors in the central nervous system (CNS), along with glycine. GABA has a homologous function to ACh, gating anion channels that allow Cl− ions to enter the post synaptic neuron. Cl− causes hyperpolarization within the neuron, decreasing the probability of an action potential firing as the voltage becomes more negative (for an action potential to fire, a positive voltage threshold must be reached). GABA is synthesized from glutamate neurotransmitters by the enzyme glutamate decarboxylase.
- Glutamatergic neurons – glutamate. Glutamate is one of two primary excitatory amino acid neurotransmitters, along with aspartate. Glutamate receptors are one of four categories, three of which are ligand-gated ion channels and one of which is a G-protein coupled receptor (often referred to as GPCR).
-
- AMPA and Kainate receptors function as cation channels permeable to Na+ cation channels mediating fast excitatory synaptic transmission.
- NMDA receptors are another cation channel that is more permeable to Ca2+. The function of NMDA receptors depend on glycine receptor binding as a co-agonist within the channel pore. NMDA receptors do not function without both ligands present.
- Metabotropic receptors, GPCRs modulate synaptic transmission and postsynaptic excitability.
- Glutamate can cause excitotoxicity when blood flow to the brain is interrupted, resulting in brain damage. When blood flow is suppressed, glutamate is released from presynaptic neurons, causing greater NMDA and AMPA receptor activation than normal outside of stress conditions, leading to elevated Ca2+ and Na+ entering the post synaptic neuron and cell damage. Glutamate is synthesized from the amino acid glutamine by the enzyme glutamate synthase.
- Dopaminergic neurons—dopamine. Dopamine is a neurotransmitter that acts on D1 type (D1 and D5) Gs-coupled receptors, which increase cAMP and PKA, and D2 type (D2, D3, and D4) receptors, which activate Gi-coupled receptors that decrease cAMP and PKA. Dopamine is connected to mood and behavior and modulates both pre- and post-synaptic neurotransmission. Loss of dopamine neurons in the substantia nigra has been linked to Parkinson’s disease. Dopamine is synthesized from the amino acid tyrosine. Tyrosine is catalyzed into levodopa (or L-DOPA) by tyrosine hydroxlase, and levodopa is then converted into dopamine by the aromatic amino acid decarboxylase.
- Serotonergic neurons—serotonin. Serotonin (5-Hydroxytryptamine, 5-HT) can act as excitatory or inhibitory. Of its four 5-HT receptor classes, 3 are GPCR and 1 is a ligand-gated cation channel. Serotonin is synthesized from tryptophan by tryptophan hydroxylase, and then further by decarboxylase. A lack of 5-HT at postsynaptic neurons has been linked to depression. Drugs that block the presynaptic serotonin transporter are used for treatment, such as Prozac and Zoloft.
- Purinergic neurons—ATP. ATP is a neurotransmitter acting at both ligand-gated ion channels (P2X receptors) and GPCRs (P2Y) receptors. ATP is, however, best known as a cotransmitter. Such purinergic signalling can also be mediated by other purines like adenosine, which particularly acts at P2Y receptors.
- Histaminergic neurons—histamine. Histamine is a monoamine neurotransmitter and neuromodulator. Histamine-producing neurons are found in the tuberomammillary nucleus of the hypothalamus.[21] Histamine is involved in arousal and regulating sleep/wake behaviors.
Multimodel classification[edit]
Since 2012 there has been a push from the cellular and computational neuroscience community to come up with a universal classification of neurons that will apply to all neurons in the brain as well as across species. This is done by considering the three essential qualities of all neurons: electrophysiology, morphology, and the individual transcriptome of the cells. Besides being universal this classification has the advantage of being able to classify astrocytes as well. A method called Patch-Seq in which all three qualities can be measured at once is used extensively by the Allen Institute for Brain Science.[22]
Connectivity[edit]

A signal propagating down an axon to the cell body and dendrites of the next cell

Neurons communicate with each other via synapses, where either the axon terminal of one cell contacts another neuron’s dendrite, soma or, less commonly, axon. Neurons such as Purkinje cells in the cerebellum can have over 1000 dendritic branches, making connections with tens of thousands of other cells; other neurons, such as the magnocellular neurons of the supraoptic nucleus, have only one or two dendrites, each of which receives thousands of synapses.
Synapses can be excitatory or inhibitory, either increasing or decreasing activity in the target neuron, respectively. Some neurons also communicate via electrical synapses, which are direct, electrically conductive junctions between cells.[23]
When an action potential reaches the axon terminal, it opens voltage-gated calcium channels, allowing calcium ions to enter the terminal. Calcium causes synaptic vesicles filled with neurotransmitter molecules to fuse with the membrane, releasing their contents into the synaptic cleft. The neurotransmitters diffuse across the synaptic cleft and activate receptors on the postsynaptic neuron. High cytosolic calcium in the axon terminal triggers mitochondrial calcium uptake, which, in turn, activates mitochondrial energy metabolism to produce ATP to support continuous neurotransmission.[24]
An autapse is a synapse in which a neuron’s axon connects to its own dendrites.
The human brain has some 8.6 x 1010 (eighty six billion) neurons.[25] Each neuron has on average 7,000 synaptic connections to other neurons. It has been estimated that the brain of a three-year-old child has about 1015 synapses (1 quadrillion). This number declines with age, stabilizing by adulthood. Estimates vary for an adult, ranging from 1014 to 5 x 1014 synapses (100 to 500 trillion).[26]
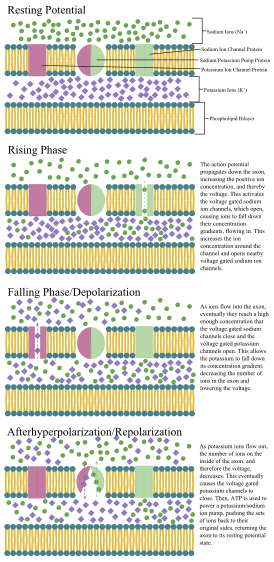
An annotated diagram of the stages of an action potential propagating down an axon including the role of ion concentration and pump and channel proteins
Nonelectrochemical signaling[edit]
Beyond electrical and chemical signaling, studies suggest neurons in healthy human brains can also communicate through:
- force generated by the enlargement of dendritic spines[27]
- the transfer of proteins – transneuronally transported proteins (TNTPs)[28][29]
They can also get modulated by input from the environment and hormones released from other parts of the organism,[30] which could be influenced more or less directly by neurons. This also applies to neurotrophins such as BDNF. The gut microbiome is also connected with the brain.[31]
Neurons also communicate with microglia, the brain’s main immune cells via specialised contact sites, called «somatic junctions». These connections enable microglia to constantly monitor and regulate neuronal functions, and exert neuroprotection, when needed.[32]
Mechanisms for propagating action potentials[edit]
In 1937 John Zachary Young suggested that the squid giant axon could be used to study neuronal electrical properties.[33] It is larger than but similar to human neurons, making it easier to study. By inserting electrodes into the squid giant axons, accurate measurements were made of the membrane potential.
The cell membrane of the axon and soma contain voltage-gated ion channels that allow the neuron to generate and propagate an electrical signal (an action potential). Some neurons also generate subthreshold membrane potential oscillations. These signals are generated and propagated by charge-carrying ions including sodium (Na+), potassium (K+), chloride (Cl−), and calcium (Ca2+).
Several stimuli can activate a neuron leading to electrical activity, including pressure, stretch, chemical transmitters, and changes of the electric potential across the cell membrane.[34] Stimuli cause specific ion-channels within the cell membrane to open, leading to a flow of ions through the cell membrane, changing the membrane potential. Neurons must maintain the specific electrical properties that define their neuron type.[35]
Thin neurons and axons require less metabolic expense to produce and carry action potentials, but thicker axons convey impulses more rapidly. To minimize metabolic expense while maintaining rapid conduction, many neurons have insulating sheaths of myelin around their axons. The sheaths are formed by glial cells: oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system. The sheath enables action potentials to travel faster than in unmyelinated axons of the same diameter, whilst using less energy. The myelin sheath in peripheral nerves normally runs along the axon in sections about 1 mm long, punctuated by unsheathed nodes of Ranvier, which contain a high density of voltage-gated ion channels. Multiple sclerosis is a neurological disorder that results from demyelination of axons in the central nervous system.
Some neurons do not generate action potentials, but instead generate a graded electrical signal, which in turn causes graded neurotransmitter release. Such non-spiking neurons tend to be sensory neurons or interneurons, because they cannot carry signals long distances.
Neural coding[edit]
Neural coding is concerned with how sensory and other information is represented in the brain by neurons. The main goal of studying neural coding is to characterize the relationship between the stimulus and the individual or ensemble neuronal responses, and the relationships among the electrical activities of the neurons within the ensemble.[36] It is thought that neurons can encode both digital and analog information.[37]
All-or-none principle[edit]
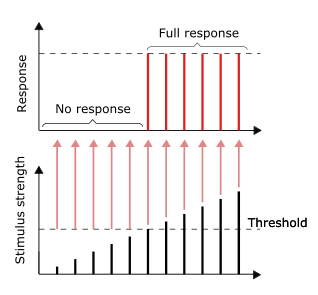
As long as the stimulus reaches the threshold, the full response would be given. Larger stimulus does not result in a larger response, vice versa.[38]: 31
The conduction of nerve impulses is an example of an all-or-none response. In other words, if a neuron responds at all, then it must respond completely. Greater intensity of stimulation, like brighter image/louder sound, does not produce a stronger signal, but can increase firing frequency.[38]: 31 Receptors respond in different ways to stimuli. Slowly adapting or tonic receptors respond to steady stimulus and produce a steady rate of firing. Tonic receptors most often respond to increased intensity of stimulus by increasing their firing frequency, usually as a power function of stimulus plotted against impulses per second. This can be likened to an intrinsic property of light where greater intensity of a specific frequency (color) requires more photons, as the photons can’t become «stronger» for a specific frequency.
Other receptor types include quickly adapting or phasic receptors, where firing decreases or stops with steady stimulus; examples include skin which, when touched causes neurons to fire, but if the object maintains even pressure, the neurons stop firing. The neurons of the skin and muscles that are responsive to pressure and vibration have filtering accessory structures that aid their function.
The pacinian corpuscle is one such structure. It has concentric layers like an onion, which form around the axon terminal. When pressure is applied and the corpuscle is deformed, mechanical stimulus is transferred to the axon, which fires. If the pressure is steady, stimulus ends; thus, typically these neurons respond with a transient depolarization during the initial deformation and again when the pressure is removed, which causes the corpuscle to change shape again. Other types of adaptation are important in extending the function of a number of other neurons.[39]
Etymology and spelling[edit]
The German anatomist Heinrich Wilhelm Waldeyer introduced the term neuron in 1891,[40] based on the ancient Greek νεῦρον neuron ‘sinew, cord, nerve’.[41]
The word was adopted in French with the spelling neurone. That spelling was also used by many writers in English,[42] but has now become rare in American usage and uncommon in British usage.[43][41]
History[edit]

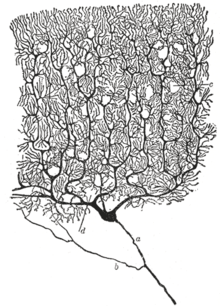
The neuron’s place as the primary functional unit of the nervous system was first recognized in the late 19th century through the work of the Spanish anatomist Santiago Ramón y Cajal.[44]
To make the structure of individual neurons visible, Ramón y Cajal improved a silver staining process that had been developed by Camillo Golgi.[44] The improved process involves a technique called «double impregnation» and is still in use.
In 1888 Ramón y Cajal published a paper about the bird cerebellum. In this paper, he stated that he could not find evidence for anastomosis between axons and dendrites and called each nervous element «an absolutely autonomous canton.»[44][40] This became known as the neuron doctrine, one of the central tenets of modern neuroscience.[44]
In 1891, the German anatomist Heinrich Wilhelm Waldeyer wrote a highly influential review of the neuron doctrine in which he introduced the term neuron to describe the anatomical and physiological unit of the nervous system.[45][46]
The silver impregnation stains are a useful method for neuroanatomical investigations because, for reasons unknown, it stains only a small percentage of cells in a tissue, exposing the complete micro structure of individual neurons without much overlap from other cells.[47]
Neuron doctrine[edit]

The neuron doctrine is the now fundamental idea that neurons are the basic structural and functional units of the nervous system. The theory was put forward by Santiago Ramón y Cajal in the late 19th century. It held that neurons are discrete cells (not connected in a meshwork), acting as metabolically distinct units.
Later discoveries yielded refinements to the doctrine. For example, glial cells, which are non-neuronal, play an essential role in information processing.[48] Also, electrical synapses are more common than previously thought,[49] comprising direct, cytoplasmic connections between neurons. In fact, neurons can form even tighter couplings: the squid giant axon arises from the fusion of multiple axons.[50]
Ramón y Cajal also postulated the Law of Dynamic Polarization, which states that a neuron receives signals at its dendrites and cell body and transmits them, as action potentials, along the axon in one direction: away from the cell body.[51] The Law of Dynamic Polarization has important exceptions; dendrites can serve as synaptic output sites of neurons[52] and axons can receive synaptic inputs.[53]
Compartmental modelling of neurons[edit]
Although neurons are often described of as «fundamental units» of the brain, they perform internal computations. Neurons integrate input within dendrites, and this complexity is lost in models that assume neurons to be a fundamental unit. Dendritic branches can be modeled as spatial compartments, whose activity is related due to passive membrane properties, but may also be different depending on input from synapses. Compartmental modelling of dendrites is especially helpful for understanding the behavior of neurons that are too small to record with electrodes, as is the case for Drosophila melanogaster.[54]
Neurons in the brain[edit]
The number of neurons in the brain varies dramatically from species to species.[55] In a human, there are an estimated 10–20 billion neurons in the cerebral cortex and 55–70 billion neurons in the cerebellum.[56] By contrast, the nematode worm Caenorhabditis elegans has just 302 neurons, making it an ideal model organism as scientists have been able to map all of its neurons. The fruit fly Drosophila melanogaster, a common subject in biological experiments, has around 100,000 neurons and exhibits many complex behaviors. Many properties of neurons, from the type of neurotransmitters used to ion channel composition, are maintained across species, allowing scientists to study processes occurring in more complex organisms in much simpler experimental systems.
Neurological disorders[edit]
Charcot–Marie–Tooth disease (CMT) is a heterogeneous inherited disorder of nerves (neuropathy) that is characterized by loss of muscle tissue and touch sensation, predominantly in the feet and legs extending to the hands and arms in advanced stages. Presently incurable, this disease is one of the most common inherited neurological disorders, affecting 36 in 100,000 people.[57]
Alzheimer’s disease (AD), also known simply as Alzheimer’s, is a neurodegenerative disease characterized by progressive cognitive deterioration, together with declining activities of daily living and neuropsychiatric symptoms or behavioral changes.[58] The most striking early symptom is loss of short-term memory (amnesia), which usually manifests as minor forgetfulness that becomes steadily more pronounced with illness progression, with relative preservation of older memories. As the disorder progresses, cognitive (intellectual) impairment extends to the domains of language (aphasia), skilled movements (apraxia), and recognition (agnosia), and functions such as decision-making and planning become impaired.[59][60]
Parkinson’s disease (PD), also known as Parkinsons, is a degenerative disorder of the central nervous system that often impairs motor skills and speech.[61] Parkinson’s disease belongs to a group of conditions called movement disorders.[62] It is characterized by muscle rigidity, tremor, a slowing of physical movement (bradykinesia), and in extreme cases, a loss of physical movement (akinesia). The primary symptoms are the results of decreased stimulation of the motor cortex by the basal ganglia, normally caused by the insufficient formation and action of dopamine, which is produced in the dopaminergic neurons of the brain. Secondary symptoms may include high level cognitive dysfunction and subtle language problems. PD is both chronic and progressive.
Myasthenia gravis is a neuromuscular disease leading to fluctuating muscle weakness and fatigability during simple activities. Weakness is typically caused by circulating antibodies that block acetylcholine receptors at the post-synaptic neuromuscular junction, inhibiting the stimulative effect of the neurotransmitter acetylcholine. Myasthenia is treated with immunosuppressants, cholinesterase inhibitors and, in selected cases, thymectomy.
Demyelination[edit]

Demyelination is the act of demyelinating, or the loss of the myelin sheath insulating the nerves. When myelin degrades, conduction of signals along the nerve can be impaired or lost, and the nerve eventually withers. This leads to certain neurodegenerative disorders like multiple sclerosis and chronic inflammatory demyelinating polyneuropathy.
Axonal degeneration[edit]
Although most injury responses include a calcium influx signaling to promote resealing of severed parts, axonal injuries initially lead to acute axonal degeneration, which is the rapid separation of the proximal and distal ends, occurring within 30 minutes of injury.[63] Degeneration follows with swelling of the axolemma, and eventually leads to bead-like formation. Granular disintegration of the axonal cytoskeleton and inner organelles occurs after axolemma degradation. Early changes include accumulation of mitochondria in the paranodal regions at the site of injury. Endoplasmic reticulum degrades and mitochondria swell up and eventually disintegrate. The disintegration is dependent on ubiquitin and calpain proteases (caused by the influx of calcium ion), suggesting that axonal degeneration is an active process that produces complete fragmentation. The process takes about roughly 24 hours in the PNS and longer in the CNS. The signaling pathways leading to axolemma degeneration are unknown.
Neurogenesis[edit]
Neurons are born through the process of neurogenesis, in which neural stem cells divide to produce differentiated neurons. Once fully differentiated neurons are formed, they are no longer capable of undergoing mitosis. Neurogenesis primarily occurs in the embryo of most organisms.
Adult neurogenesis can occur and studies of the age of human neurons suggest that this process occurs only for a minority of cells, and that the vast majority of neurons in the neocortex forms before birth and persists without replacement. The extent to which adult neurogenesis exists in humans, and its contribution to cognition are controversial, with conflicting reports published in 2018.[64]
The body contains a variety of stem cell types that have the capacity to differentiate into neurons. Researchers found a way to transform human skin cells into nerve cells using transdifferentiation, in which «cells are forced to adopt new identities».[65]
During neurogenesis in the mammalian brain, progenitor and stem cells progress from proliferative divisions to differentiative divisions. This progression leads to the neurons and glia that populate cortical layers. Epigenetic modifications play a key role in regulating gene expression in differentiating neural stem cells, and are critical for cell fate determination in the developing and adult mammalian brain. Epigenetic modifications include DNA cytosine methylation to form 5-methylcytosine and 5-methylcytosine demethylation.[66] These modifications are critical for cell fate determination in the developing and adult mammalian brain. DNA cytosine methylation is catalyzed by DNA methyltransferases (DNMTs). Methylcytosine demethylation is catalyzed in several stages by TET enzymes that carry out oxidative reactions (e.g. 5-methylcytosine to 5-hydroxymethylcytosine) and enzymes of the DNA base excision repair (BER) pathway.[66]
At different stages of mammalian nervous system development two DNA repair processes are employed in the repair of DNA double-strand breaks. These pathways are homologous recombinational repair used in proliferating neural precursor cells, and non-homologous end joining used mainly at later developmental stages[67]
Intercellular communication between developing neurons and microglia is also indispensable for proper neurogenesis and brain development.[68]
Nerve regeneration[edit]
Peripheral axons can regrow if they are severed,[69] but one neuron cannot be functionally replaced by one of another type (Llinás’ law).[18]
See also[edit]
- Artificial neuron
- Bidirectional cell
- Biological neuron model
- Compartmental neuron models
- Connectome
- Dogiel cell
- List of animals by number of neurons
- List of neuroscience databases
- Neuronal galvanotropism
- Neuroplasticity
- Growth cone
- Sholl analysis
References[edit]
- ^ >Moore, Keith; Dalley, Arthur (2005). Clinically Oriented Anatomy (5th ed.). LWW. pp. 47. ISBN 0-7817-3639-0.
A bundle of nerve fibers (axons) connecting neighboring or distant nuclei of the CNS is a tract.
- ^ «What are the parts of the nervous system?». Retrieved 2022-07-08.
- ^ Davies, Melissa (2002-04-09). «The Neuron: size comparison». Neuroscience: A journey through the brain. Retrieved 2009-06-20.
- ^ Chudler EH. «Brain Facts and Figures». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ «16.7: Nervous System». Biology LibreTexts. 2021-01-14. Retrieved 2022-02-28.
- ^ Herrup K, Yang Y (May 2007). «Cell cycle regulation in the postmitotic neuron: oxymoron or new biology?». Nature Reviews. Neuroscience. 8 (5): 368–78. doi:10.1038/nrn2124. PMID 17453017. S2CID 12908713.
- ^ Giménez, C. (February 1998). «[Composition and structure of the neuronal membrane: molecular basis of its physiology and pathology]». Revista de Neurologia. 26 (150): 232–239. ISSN 0210-0010. PMID 9563093.
- ^ State Hospitals Bulletin. State Commission in Lunacy. 1897. p. 378.
- ^ «Medical Definition of Neurotubules». www.merriam-webster.com.
- ^ Zecca L, Gallorini M, Schünemann V, Trautwein AX, Gerlach M, Riederer P, Vezzoni P, Tampellini D (March 2001). «Iron, neuromelanin and ferritin content in the substantia nigra of normal subjects at different ages: consequences for iron storage and neurodegenerative processes». Journal of Neurochemistry. 76 (6): 1766–73. doi:10.1046/j.1471-4159.2001.00186.x. PMID 11259494. S2CID 31301135.
- ^ Herrero MT, Hirsch EC, Kastner A, Luquin MR, Javoy-Agid F, Gonzalo LM, Obeso JA, Agid Y (1993). «Neuromelanin accumulation with age in catecholaminergic neurons from Macaca fascicularis brainstem». Developmental Neuroscience. 15 (1): 37–48. doi:10.1159/000111315. PMID 7505739.
- ^ Brunk UT, Terman A (September 2002). «Lipofuscin: mechanisms of age-related accumulation and influence on cell function». Free Radical Biology & Medicine. 33 (5): 611–9. doi:10.1016/s0891-5849(02)00959-0. PMID 12208347.
- ^ Zhao B, Meka DP, Scharrenberg R, König T, Schwanke B, Kobler O, Windhorst S, Kreutz MR, Mikhaylova M, Calderon de Anda F (August 2017). «Microtubules Modulate F-actin Dynamics during Neuronal Polarization». Scientific Reports. 7 (1): 9583. Bibcode:2017NatSR…7.9583Z. doi:10.1038/s41598-017-09832-8. PMC 5575062. PMID 28851982.
- ^ Lee WC, Huang H, Feng G, Sanes JR, Brown EN, So PT, Nedivi E (February 2006). «Dynamic remodeling of dendritic arbors in GABAergic interneurons of adult visual cortex». PLOS Biology. 4 (2): e29. doi:10.1371/journal.pbio.0040029. PMC 1318477. PMID 16366735.
- ^ Al, Martini, Frederic Et (2005). Anatomy and Physiology’ 2007 Ed.2007 Edition. Rex Bookstore, Inc. p. 288. ISBN 978-971-23-4807-5.
- ^ Gerber U (January 2003). «Metabotropic glutamate receptors in vertebrate retina». Documenta Ophthalmologica. Advances in Ophthalmology. 106 (1): 83–7. doi:10.1023/A:1022477203420. PMID 12675489. S2CID 22296630.
- ^ Wilson NR, Runyan CA, Wang FL, Sur M (August 2012). «Division and subtraction by distinct cortical inhibitory networks in vivo». Nature. 488 (7411): 343–8. Bibcode:2012Natur.488..343W. doi:10.1038/nature11347. hdl:1721.1/92709. PMC 3653570. PMID 22878717.
- ^ a b Llinás RR (2014-01-01). «Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective». Frontiers in Cellular Neuroscience. 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ Kolodin YO, Veselovskaia NN, Veselovsky NS, Fedulova SA. Ion conductances related to shaping the repetitive firing in rat retinal ganglion cells. Acta Physiologica Congress. Archived from the original on 2012-10-07. Retrieved 2009-06-20.
- ^ «Ionic conductances underlying excitability in tonically firing retinal ganglion cells of adult rat». Ykolodin.50webs.com. 2008-04-27. Retrieved 2013-02-16.
- ^ Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y (January 2019). «Histamine: neural circuits and new medications». Sleep. 42 (1). doi:10.1093/sleep/zsy183. PMC 6335869. PMID 30239935.
- ^ «Patch-seq technique helps depict the variation of neural cells in the brain». News-medical.net. 3 December 2020. Retrieved 26 August 2021.
{{cite web}}: CS1 maint: url-status (link) - ^ Macpherson, Gordon (2002). Black’s Medical Dictionary (40 ed.). Lanham, MD: Scarecrow Press. pp. 431–434. ISBN 0810849844.
- ^ Ivannikov MV, Macleod GT (June 2013). «Mitochondrial free Ca²⁺ levels and their effects on energy metabolism in Drosophila motor nerve terminals». Biophysical Journal. 104 (11): 2353–61. Bibcode:2013BpJ…104.2353I. doi:10.1016/j.bpj.2013.03.064. PMC 3672877. PMID 23746507.
- ^ Herculano-Houzel S (November 2009). «The human brain in numbers: a linearly scaled-up primate brain». Frontiers in Human Neuroscience. 3: 31. doi:10.3389/neuro.09.031.2009. PMC 2776484. PMID 19915731.
- ^ Drachman DA (June 2005). «Do we have brain to spare?». Neurology. 64 (12): 2004–5. doi:10.1212/01.WNL.0000166914.38327.BB. PMID 15985565. S2CID 38482114.
- ^ Ucar, Hasan; Watanabe, Satoshi; Noguchi, Jun; Morimoto, Yuichi; Iino, Yusuke; Yagishita, Sho; Takahashi, Noriko; Kasai, Haruo (December 2021). «Mechanical actions of dendritic-spine enlargement on presynaptic exocytosis». Nature. 600 (7890): 686–689. Bibcode:2021Natur.600..686U. doi:10.1038/s41586-021-04125-7. ISSN 1476-4687. PMID 34819666. S2CID 244648506.
Lay summary:
«Forceful synapses reveal mechanical interactions in the brain». Nature. 24 November 2021. doi:10.1038/d41586-021-03516-0. Retrieved 21 February 2022. - ^ «Researchers discover new type of cellular communication in the brain». The Scripps Research Institute. Retrieved 12 February 2022.
- ^ Schiapparelli, Lucio M.; Sharma, Pranav; He, Hai-Yan; Li, Jianli; Shah, Sahil H.; McClatchy, Daniel B.; Ma, Yuanhui; Liu, Han-Hsuan; Goldberg, Jeffrey L.; Yates, John R.; Cline, Hollis T. (25 January 2022). «Proteomic screen reveals diverse protein transport between connected neurons in the visual system». Cell Reports. 38 (4): 110287. doi:10.1016/j.celrep.2021.110287. ISSN 2211-1247. PMC 8906846. PMID 35081342.
- ^ Levitan, Irwin B.; Kaczmarek, Leonard K. (2015). «Electrical Signaling in Neurons». The Neuron. Oxford University Press. pp. 41–62. doi:10.1093/med/9780199773893.003.0003. ISBN 978-0-19-977389-3.
- ^ O’Leary, Olivia F.; Ogbonnaya, Ebere S.; Felice, Daniela; Levone, Brunno R.; C. Conroy, Lorraine; Fitzgerald, Patrick; Bravo, Javier A.; Forsythe, Paul; Bienenstock, John; Dinan, Timothy G.; Cryan, John F. (1 February 2018). «The vagus nerve modulates BDNF expression and neurogenesis in the hippocampus». European Neuropsychopharmacology. 28 (2): 307–316. doi:10.1016/j.euroneuro.2017.12.004. ISSN 0924-977X. PMID 29426666. S2CID 46819013.
- ^ Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z (January 2020). «Microglia monitor and protect neuronal function through specialized somatic purinergic junctions». Science. 367 (6477): 528–537. Bibcode:2020Sci…367..528C. doi:10.1126/science.aax6752. PMID 31831638. S2CID 209343260.
- ^ Chudler EH. «Milestones in Neuroscience Research». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ Patlak J, Gibbons R (2000-11-01). «Electrical Activity of Nerves». Action Potentials in Nerve Cells. Archived from the original on August 27, 2009. Retrieved 2009-06-20.
- ^ Harris-Warrick, RM (October 2011). «Neuromodulation and flexibility in Central Pattern Generator networks». Current Opinion in Neurobiology. 21 (5): 685–92. doi:10.1016/j.conb.2011.05.011. PMC 3171584. PMID 21646013.
- ^ Brown EN, Kass RE, Mitra PP (May 2004). «Multiple neural spike train data analysis: state-of-the-art and future challenges». Nature Neuroscience. 7 (5): 456–61. doi:10.1038/nn1228. PMID 15114358. S2CID 562815.
- ^ Thorpe SJ (1990). «Spike arrival times: A highly efficient coding scheme for neural networks» (PDF). In Eckmiller R, Hartmann G, Hauske G (eds.). Parallel processing in neural systems and computers. North-Holland. pp. 91–94. ISBN 9780444883902. Archived from the original (PDF) on 2012-02-15.
- ^ a b Kalat, James W (2016). Biological psychology (12 ed.). Australia. ISBN 9781305105409. OCLC 898154491.
- ^ Eckert R, Randall D (1983). Animal physiology: mechanisms and adaptations. San Francisco: W.H. Freeman. p. 239. ISBN 978-0-7167-1423-1.
- ^ a b Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
Ramon y Cajal’s first paper on the Golgi stain was on the bird cerebellum, and it appeared in the Revista in 1888. He acknowledged that he found the nerve fibers to be very intricate, but stated that he could find no evidence for either axons or dendrites undergoing anastomosis and forming nets. He called each nervous element ‘an absolutely autonomous canton.’
- ^ a b Oxford English Dictionary, 3rd edition, 2003, s.v.
- ^ Mehta AR, Mehta PR, Anderson SP, MacKinnon BL, Compston A (January 2020). «Grey Matter Etymology and the neuron(e)». Brain. 143 (1): 374–379. doi:10.1093/brain/awz367. PMC 6935745. PMID 31844876.
- ^ «Google Books Ngram Viewer». books.google.com. Retrieved 19 December 2020.
- ^ a b c d López-Muñoz F, Boya J, Alamo C (October 2006). «Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal». Brain Research Bulletin. 70 (4–6): 391–405. doi:10.1016/j.brainresbull.2006.07.010. PMID 17027775. S2CID 11273256.
- ^ Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
… a man who would write a highly influential review of the evidence in favor of the neuron doctrine two years later. In his paper, Waldeyer (1891), … , wrote that nerve cells terminate freely with end arborizations and that the ‘neuron’ is the anatomical and physiological unit of the nervous system. The word ‘neuron’ was born this way.
- ^ «Whonamedit — dictionary of medical eponyms». www.whonamedit.com.
Today, Wilhelm von Waldeyer-Hartz is remembered as the founder of the neurone theory, coining the term «neurone» to describe the cellular function unit of the nervous system and enunciating and clarifying that concept in 1891.
- ^ Grant G (October 2007). «How the 1906 Nobel Prize in Physiology or Medicine was shared between Golgi and Cajal». Brain Research Reviews. 55 (2): 490–8. doi:10.1016/j.brainresrev.2006.11.004. PMID 17306375. S2CID 24331507.
- ^ Witcher MR, Kirov SA, Harris KM (January 2007). «Plasticity of perisynaptic astroglia during synaptogenesis in the mature rat hippocampus». Glia. 55 (1): 13–23. CiteSeerX 10.1.1.598.7002. doi:10.1002/glia.20415. PMID 17001633. S2CID 10664003.
- ^ Connors BW, Long MA (2004). «Electrical synapses in the mammalian brain». Annual Review of Neuroscience. 27 (1): 393–418. doi:10.1146/annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Guillery RW (June 2005). «Observations of synaptic structures: origins of the neuron doctrine and its current status». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1458): 1281–307. doi:10.1098/rstb.2003.1459. PMC 1569502. PMID 16147523.
- ^ Sabbatini RM (April–July 2003). «Neurons and Synapses: The History of Its Discovery». Brain & Mind Magazine: 17.
- ^ Djurisic M, Antic S, Chen WR, Zecevic D (July 2004). «Voltage imaging from dendrites of mitral cells: EPSP attenuation and spike trigger zones». The Journal of Neuroscience. 24 (30): 6703–14. doi:10.1523/JNEUROSCI.0307-04.2004. hdl:1912/2958. PMC 6729725. PMID 15282273.
- ^ Cochilla AJ, Alford S (March 1997). «Glutamate receptor-mediated synaptic excitation in axons of the lamprey». The Journal of Physiology. 499 (Pt 2): 443–57. doi:10.1113/jphysiol.1997.sp021940. PMC 1159318. PMID 9080373.
- ^ Gouwens NW, Wilson RI (2009). «Signal propagation in Drosophila central neurons». Journal of Neuroscience. 29 (19): 6239–6249. doi:10.1523/jneurosci.0764-09.2009. PMC 2709801. PMID 19439602.
- ^ Williams RW, Herrup K (1988). «The control of neuron number». Annual Review of Neuroscience. 11 (1): 423–53. doi:10.1146/annurev.ne.11.030188.002231. PMID 3284447.
- ^ von Bartheld CS, Bahney J, Herculano-Houzel S (December 2016). «The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting». The Journal of Comparative Neurology. 524 (18): 3865–3895. doi:10.1002/cne.24040. PMC 5063692. PMID 27187682.
- ^ Krajewski KM, Lewis RA, Fuerst DR, Turansky C, Hinderer SR, Garbern J, Kamholz J, Shy ME (July 2000). «Neurological dysfunction and axonal degeneration in Charcot-Marie-Tooth disease type 1A». Brain. 123 (7): 1516–27. doi:10.1093/brain/123.7.1516. PMID 10869062.
- ^ «About Alzheimer’s Disease: Symptoms». National Institute on Aging. Archived from the original on 15 January 2012. Retrieved 28 December 2011.
- ^ Burns A, Iliffe S (February 2009). «Alzheimer’s disease». BMJ. 338: b158. doi:10.1136/bmj.b158. PMID 19196745. S2CID 8570146.
- ^ Querfurth HW, LaFerla FM (January 2010). «Alzheimer’s disease». The New England Journal of Medicine. 362 (4): 329–44. doi:10.1056/NEJMra0909142. PMID 20107219. S2CID 205115756.
- ^ «Parkinson’s Disease Information Page». NINDS. 30 June 2016. Archived from the original on 4 January 2017. Retrieved 18 July 2016.
- ^ «Movement Disorders». The International Neuromodulation Society.
- ^ Kerschensteiner M, Schwab ME, Lichtman JW, Misgeld T (May 2005). «In vivo imaging of axonal degeneration and regeneration in the injured spinal cord». Nature Medicine. 11 (5): 572–7. doi:10.1038/nm1229. PMID 15821747. S2CID 25287010.
- ^ Kempermann G, Gage FH, Aigner L, Song H, Curtis MA, Thuret S, Kuhn HG, Jessberger S, Frankland PW, Cameron HA, Gould E, Hen R, Abrous DN, Toni N, Schinder AF, Zhao X, Lucassen PJ, Frisén J (July 2018). «Human Adult Neurogenesis: Evidence and Remaining Questions». Cell Stem Cell. 23 (1): 25–30. doi:10.1016/j.stem.2018.04.004. PMC 6035081. PMID 29681514.
- ^ Callaway, Ewen (26 May 2011). «How to make a human neuron». Nature. doi:10.1038/news.2011.328.
By transforming cells from human skin into working nerve cells, researchers may have come up with a model for nervous-system diseases and perhaps even regenerative therapies based on cell transplants. The achievement, reported online today in Nature, is the latest in a fast-moving field called transdifferentiation, in which cells are forced to adopt new identities. In the past year, researchers have converted connective tissue cells found in skin into heart cells, blood cells, and liver cells.
- ^ a b Wang Z, Tang B, He Y, Jin P (March 2016). «DNA methylation dynamics in neurogenesis». Epigenomics. 8 (3): 401–14. doi:10.2217/epi.15.119. PMC 4864063. PMID 26950681.
- ^ Orii KE, Lee Y, Kondo N, McKinnon PJ (June 2006). «Selective utilization of nonhomologous end-joining and homologous recombination DNA repair pathways during nervous system development». Proceedings of the National Academy of Sciences of the United States of America. 103 (26): 10017–22. Bibcode:2006PNAS..10310017O. doi:10.1073/pnas.0602436103. PMC 1502498. PMID 16777961.
- ^ Cserép, Csaba; Schwarcz, Anett D.; Pósfai, Balázs; László, Zsófia I.; Kellermayer, Anna; Környei, Zsuzsanna; Kisfali, Máté; Nyerges, Miklós; Lele, Zsolt; Katona, István (September 2022). «Microglial control of neuronal development via somatic purinergic junctions». Cell Reports. 40 (12): 111369. doi:10.1016/j.celrep.2022.111369. PMC 9513806. PMID 36130488. S2CID 252416407.
- ^ Yiu G, He Z (August 2006). «Glial inhibition of CNS axon regeneration». Nature Reviews. Neuroscience. 7 (8): 617–27. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
Further reading[edit]
- Bullock TH, Bennett MV, Johnston D, Josephson R, Marder E, Fields RD (November 2005). «Neuroscience. The neuron doctrine, redux». Science. 310 (5749): 791–3. doi:10.1126/science.1114394. PMID 16272104. S2CID 170670241.
- Kandel ER, Schwartz JH, Jessell TM (2000). Principles of Neural Science (4th ed.). New York: McGraw-Hill. ISBN 0-8385-7701-6.
- Peters A, Palay SL, Webster HS (1991). The Fine Structure of the Nervous System (3rd ed.). New York: Oxford University Press. ISBN 0-19-506571-9.
- Ramón y Cajal S (1933). Histology (10th ed.). Baltimore: Wood.
- Roberts A, Bush BM (1981). Neurones without Impulses. Cambridge: Cambridge University Press. ISBN 0-521-29935-7.
- Snell RS (2010). Clinical Neuroanatomy. Lippincott Williams & Wilkins. ISBN 978-0-7817-9427-5.
External links[edit]
- Neurobiology at Curlie
- IBRO (International Brain Research Organization). Fostering neuroscience research especially in less well-funded countries.
- NeuronBank an online neuromics tool for cataloging neuronal types and synaptic connectivity.
- High Resolution Neuroanatomical Images of Primate and Non-Primate Brains.
- The Department of Neuroscience at Wikiversity, which presently offers two courses: Fundamentals of Neuroscience and Comparative Neuroscience.
- NIF Search – Neuron via the Neuroscience Information Framework
- Cell Centered Database – Neuron
- Complete list of neuron types according to the Petilla convention, at NeuroLex.
- NeuroMorpho.Org an online database of digital reconstructions of neuronal morphology.
- Immunohistochemistry Image Gallery: Neuron
- Khan Academy: Anatomy of a neuron
- Neuron images
[править]
Почему у нейрона один аксон?
<- назад
дальше ->
Действительно, а в чем причина этого? Если природа обошлась единственным каналом вывода нервных сигналов из нейрона, значит его вполне достаточно для совершаемых нервной системой актов. Любой нейрон в нервной системе почему-то всегда имеет только один аксон. Если уж и есть нейроны с несколькими аксонами, то только за счет «склеивания» в процессе развития организма нескольких одноаксонных нейронов в один большой нейрон. На первый взгляд получается, что выходная «шина данных» обычного нейрона состоит из единственного «провода», но поскольку хотя бы даже на малых участках нервной системы все-таки предполагается быть транспортировке существенно емких нервных сигналов, вероятно включающих произвольные «порции» потребных сведений, то в свете алгоритмической интерпретации совершаемых нервной системой актов выглядит неясным вопрос о том, как природой решена проблема «разрядности», «структурности» портируемых на выход нейрона данных. В связи с этим в среде лиц, причастных к проектированию самостоятельных «аналогов» нейронов, обычно возникают два противоборствующих объяснения: аксон или является одноканальной шиной для последовательной передачи ненормированной поточной информации, либо алгоритм работы ядра нейрона основан на строгом акцентировании «типоразмера» обслуживаемых данных, четком структурировании выходящего информационного потока.
Попробуем посильно разобрать каждое объяснение с учетом искусственной аппаратной или же программной реализации природных особенностей работы выходного канала нейрона. Той самой особенности, которую приходится поневоле воображать, моделировать в уме с помощью плохо обоснованных допущений, на основе банальных представлений о передаче информации в компьютерных системах, доверяясь мнимой некоторой общности таких представлений со спецификой передачи оперативной информации в нервных сигналах. Начнем со второго объяснения как знакомого всякому, кто хоть мельком касался темы способов представления данных в компьютерах.
Структурирование данных в принципе и есть предварительное описание содержимого информационного потока, коим может выступать тот гипотетически емкий нервный сигнал, только в обычном случае поля структуры (контейнеры «порций» соответствующих сведений) строго фиксированы по емкости несомой в них информации. Структурированные данные несложно передавать одноканальным потоком, и значительный плюс здесь заключен вот в чем: нет нужды вносить в поток дополнительные информационные элементы, отделяющие поля структуры друг от друга, ведь отправляющий и принимающий связаны единым соглашением о порядке и размере следуемых в потоке «порций» данных. Касаемо внутреннего устройства нейрона это означает, что последний в таком случае избавлен от необходимости иметь в своем составе аппаратные или программные компоненты, следящие за разделительными признаками во входном потоке или же, наоборот, формирующие такие признаки в выходном потоке нейрона.
Но при подобных условиях искусственный нейрон не в состоянии передавать или принимать за один «такт» деятельности информации больше или меньше, чем ограничено размером выбранной структуры. Поскольку нейроны в организме конструируются на одной и той же «элементной» базе, а также существует великое разнообразие транспортируемых через них типов информации, то предшествующая проектировка полей структуры информационного потока должна удовлетворять требованиям всех мыслимых нервных узлов и центров, что кажется сложным осуществить одной структурой с запрограммированным порядком и размером полей.
Более выгодной видится поддержка потоков с ненормированным порядком и размером полей, то есть безтиповое структурирование внедрением в поток специальных отметок, идентификаторов, определяющих не только концы «порций» сведений, но также их именные, классификационные приметы. Возможно и даже наиболее вероятно, что именно так или по схожей идее организована методика передачи вместительных нервных сигналов в настоящих нейронах. Подобный поток легко обрабатывать порциями, легко пропускать при необходимости его участки, легко вообще отказаться от обработки в некоторой точке потоке. Заманчивая идея: обрабатывать поток, ориентируясь на какие-то признаки внутри него. Вот только непонятно, как организовать обратную связь с отправителем потока. Как ему сообщить, что нейрон решил прервать обработку потока и отправитель должен прекратить передачу?
Конечно, первое из противоборствующих объяснений привлекает широтой возможностей, в то же время оно вносит в имитацию нейронов дополнительные трудности, обусловленные спецификой обслуживания ненормированных потоков: это и аппаратные (программные) устройства обратной связи; это и устройства отслеживания начала и конца передачи, внедренных идентификаторов и тому подобных вещей. В случае поддержки обратной связи в поток придется внедрить еще и код получателя, ибо у нейрона аксон один, а сообщить получателю свою резолюцию по обработке идущего от него потока нейрон может только с помощью своего выхода. К нему подключены и другие нейроны, ждущие своего потока данных, и в этот момент по выходной шине начинают идти сигналы какому-то неведомому им отправителю. Следовательно, у нейронов должны быть еще и устройства, выделяющие из нервного сигнала код получателя, чтобы по ошибке не отзываться на чужой сигнал.
Вообще кажется странным, почему природа создала один аксон. Что это — экономия, необходимость или неведомая технология обмена нервными сигналами? Экономя на одном аксоне как на последовательной шине ненормированных поточных данных, природа неизбежно вышла бы на весомые затраты в дендритах. Сводя предположительно многоканальную внутреннюю шину данных нейрона в одноканальный выходной сигнал, природа обязана была бы позаботиться, чтобы каждый дендрит нейрона имел аппаратные устройства по «расканаливанию» и введению поточных данных входного сигнала во внутреннюю шину данных. Крупные нейроны головного мозга имеют десятки тысяч дендритов. Выходят неимоверные аппаратные затраты, если необходимо каждый дендрит снабдить таким вот «разводящим» устройством, так сказать, демультиплексором (мультиплексор — микросхема с несколькими входами и одним выходом; управляющими сигналами осуществляется подача на выход данных одного из входных каналов). В этих аналогиях работа аксона напоминает мультиплексирование внутренней шины данных с отсроченным попаданием всех предложенных данных в выходной поток, работа дендрита — демультиплексирование.
Понятно, насколько выгодным в транспортном смысле оказывается одноканальный выход с ненормированным информационным потоком. Да только в программной имитации, в соответствии с доступными сегодня пропускными и вычислительными мощностями, целесообразным пока оказывается склонение к фиксированной размерной структуризации выходного потока. Это прямо повышает производительность запрограммированной нейронной схемы за счет подведения под работу ядра нейрона требования выводить на выход исключительно точные и краткие данные, укладывающиеся в заблаговременно описанную и единую для всех искусственных нейронов структуру информационного потока. Во-первых, это искусственно ограничивает использование памяти эмулирующего устройства (например компьютера), во-вторых, освобождает программные модели нейронов от затратных по времени функций мультиплексирования и демультиплексирования сигналов в аксонах и дендритах. Минус заключен в том, что помимо потери некоторых алгоритмических путей обработки нет никакой гарантии, что избранная статическая структура информационного потока окажется подходящей для большинства целевых нейронных схем.
Дмитрий Сахань, 7 декабря 2002 года
[править]
Справка по разделу -Теория по нейронным микросхемам
Этот неподвластный интеллект
Нейронные микросхемы
Нейронные микросхемы с обратными функциями
Заметки для экспериментов с нейронными микросхемами
Почему у нейрона один аксон?
Каков «алгоритм» работы рецептора?
Следствия в нейронах вместо данных
Каковы настоящие функции нейрона?
Препарируя нейрон
К вопросу о скорости работы нейронов
Рободелам на заметку
Рободелам на заметку 2
Как нейрон фильтрует сигналы
Идея: ядро нейрона помнит правильно выполненное следствие
А не работает ли нейрон по-другому?
К вопросу о механизме обучения нейронов
[править]
Искусственный интеллект в домашних условиях Дмитрий Сахань
Нервная система выполняет ряд важных функций:
- обеспечивает связь организма с окружающим миром;
- управляет работой всех органов;
- координирует функционирование всех систем органов, обеспечивая их согласованную работу.
Нервная ткань
Нервная ткань отличается от других тканей нашего организма тем, что обладает особыми свойствами — возбудимостью и проводимостью. Эти свойства нервной ткани обусловлены особенностями её строения.
В состав нервной ткани входят клетки двух видов. Основные функции выполняют нейроны, а клетки-спутники (клетки нейроглии) служат опорой и обеспечивают обмен веществ.

Рис. (1). Нервная ткань
Функции нейронов: генерирование и передача нервных импульсов; обработка и хранение поступающей информации.
Нервный импульс — это волна возбуждения (биоэлектрическая волна), распространяющаяся по нервным клеткам.
Нейрон — основная клетка нервной ткани. Он имеет тело и отростки двух типов. В теле нейрона располагается ядро и органоиды, а по отросткам передаются нервные импульсы.
Дендриты — это отростки, по которым нервные импульсы передаются к телу нейрона. Эти отростки сильно ветвятся. У нейрона может быть несколько дендритов.
Аксон — это отросток, по которому импульсы передаются от тела клетки. Аксон обычно ветвится только на конце. У каждого нейрона всего один аксон.

Рис. (2). Строение нейрона
Аксоны часто окружены оболочкой из жироподобного вещества миелина. Это вещество имеет белый цвет. Скопления миелинизированных аксонов образуют белое вещество головного и спинного мозга. Тела нервных клеток и дендриты не покрыты миелином. Они серого цвета, а их группы составляют серое вещество центральной нервной системы.
Передача нервных импульсов с одной клетки на другую происходит в синапсах.
Синапс — это место контакта между двумя нейронами или между нейроном и клеткой рабочего органа.
Главными элементами синапса являются мембраны двух клеток (пресинаптическая и постсинаптическая мембраны) и пространство между ними (синаптическая щель).

Рис. (3). Строение синапса
В аксоне пресинаптического нейрона вырабатывается медиатор — особое вещество, с помощью которого происходит передача нервного импульса.
Под действием нервного импульса медиатор выделяется в синаптическую щель. Рецепторы постсинаптической мембраны реагируют на его появление и генерируют возникновение нервного импульса в следующем нейроне. Так в синапсе происходит химическая передача возбуждения с одной клетки на другую.
Нейроны различаются по своему строению и выполняемым функциям.

Рис. (4). Виды нейронов
По выполняемым функциям выделяют три типа нейронов.
Чувствительные (сенсорные) нейроны проводят информацию от органов в мозг. Тела таких нейронов находятся в нервных узлах вне центральной нервной системы.
Другая группа нейронов передаёт информацию от головного и спинного мозга к органам. Это двигательные (моторные) нейроны. Их тела находятся в сером веществе центральной нервной системы, а аксоны находятся за пределами ЦНС.
Третий вид нейронов осуществляет связь между чувствительными и двигательными нейронами. Это вставочные нейроны, они находятся в головном и спинном мозге.
Скопление нейронов в головном или спинном мозге называют ядром.

Рис. (5). Типы нейронов и синапсы
Связь между органами и центральной нервной системой осуществляется через нервы.
Нерв — это орган, в состав которого входят пучки нервных волокон, покрытые соединительнотканной оболочкой.

Рис. (6). Нерв
Нервы выполняют проводниковую функцию. Они связывают головной и спинной мозг с кожей, органами чувств и с внутренними органами.
Нервы бывают чувствительные, двигательные и смешанные.
Чувствительные нервы проводят нервные импульсы от рецепторов в мозг. В их состав входят дендриты чувствительных нейронов.
Двигательные нервы состоят из аксонов двигательных нейронов. Их функция — проведение импульсов от мозга к рабочим органам.
Смешанные нервы образованы чувствительными и двигательными волокнами и способные проводить импульсы как к ЦНС, так и от ЦНС.
Нервные сплетения представлены сетчатыми скоплениями нервных волокон разных нервов, связывающих ЦНС с внутренними органами, скелетными мышцами и кожей.
Наиболее известное солнечное сплетение находится в брюшной полости.
Источники:
Рис. 1. Нервная ткань https://image.shutterstock.com/image-photo/mammalian-nervous-tissue-under-microscope-600w-74170234.jpg
Рис. 2. Строение нейрона https://image.shutterstock.com/image-vector/education-chart-biology-nerve-cell-600w-661087429.jpg
Рис. 3. Строение синапса https://image.shutterstock.com/image-illustration/gap-between-two-nerve-cells-600w-1284912691.jpg
Рис. 4. Виды нейронов https://image.shutterstock.com/image-illustration/different-kinds-neurons-scheme-structure-600w-138356969.jpg
Рис. 5. Типы нейронов и синапсы © ЯКласс
Рис. 6. Нерв https://image.shutterstock.com/image-illustration/nerve-structure-anatomy-600w-1041115012.jpg
Строение нейронов
Нейроны
отличаются большой сложностью строения.
Размеры клеток чрезвычайно разнообразны
(от 4-6 мкм до 130 мкм). Форма нейрона также
очень вариабильна, но всем нервным
клеткам свойственны отростки (один или
несколько), отходящие от тела. У человека
содержится более триллиона (10 ) нервных
клеток.
На строго
определенных этапах онтогенеза
запрограммирована массовая гибель
нейроновцентральной и периферической
нервной системы. За 1 год жизни погибает
около 10 млн. нейронов, а в течение жизни
мозг теряет около 0,1 % всех нейронов.
Гибель определяет ряд факторов:
-
выживают наиболее
активно участвующие в межклеточных
взаимодействиях нейрона ( быстрее
растут, имеют больше отростков, больше
контактов с клетками – мишенями). -
имеются гены,
ответственные за выход между жизнью
или смертью. -
сбои в кровоснабжении.
По количеству
отростков нейроны делятся на:
-
униполярные –
одноотростчатые, -
биполярные –
двуотростчатые, -
мультиполярные
– многоотростчатые.
Среди
униполярных нейронов различают истинные
униполяры,
лежащие в сетчатке
глаза, и ложные униполяры, расположенные
в спинномозговых узлах. Ложные униполяры
в процессе развития были биполярными
клетками, но затем произошло вытягивание
части клетки в длинный отросток, который
часто делает несколько оборотов вокруг
тела и затем Т- образно ветвится.
Отростки
нервных клеток отличаются по строению,
у каждой нервной клетки есть аксон или
нейрит, который идет от тела клетки в
виде тяжа, имеющего одинаковую по всей
длине толщину. Часто аксоны идут на
большие расстояния. По ходу нейрита
отходят тонкие веточки – коллатерали.
Аксон, передающий отросток и импульс
в нем, идет от клетки на периферию.
Заканчивается аксон эффектором или
двигательным окончанием в мышечной или
железистой ткани. Длина аксона может
быть более 100 см. В аксоне нет
эндоплазматической сети и свободных
рибосом, поэтому все белки секретируются
в теле, а затем транспортируются по
аксону.
Другие
отростки начинаются от тела клетки
широким основанием и сильно ветвятся.
Они называются древовидными отростками
или дендритами и являются воспринимающими
отростками, в которых импульс
распространяется к телу клетки. Дендриты
заканчиваются чувствительными нервными
окончаниями или рецепторами, специфически
воспринимающими раздражения.
Истинные
униполярные нейроны имеют только один
аксон, а восприятие импульсов осуществляется
всей поверхностью клетки. Единственным
примером унипотентных клеток у человека
являются амокриновые клетки сетчатки.
Биполярные
нейроны лежат в сетчатке глаза и имеют
аксон и один ветвящийся отросток –
дендрит
Многоотросчатые
мультиполярные нейроны широко
распространены и лежат в спинном и
головном мозге, вегетативных нервных
узлах и т.д. Эти клетки имеют один аксон
и многочисленные ветвящиеся дендриты.
В зависимости
от расположения нейроны делятся на
центральные, лежащие в головном и спинном
мозге, и периферические – это невроны
вегетативных ганглий, органных нервных
сплетений и спинномозговых узлов.
Нервные
клетки тесно взаимодействуют с сосудами.
Различают 3 варианта взаимодействия:
Нервные
клетки в организме лежат в виде цепей,
т.е. одна клетка контактирует с другой
и передает на нее свой импульс. Такие
цепи клеток называются рефлекторными
дугами. В зависимости от положения
нейронов в рефлекторной дуге они имеют
различную функцию. По функции невроны
могут быть чувствительными, двигательными,
ассоциативными и вставочными. Между
собой или с органом – мишенью нервные
клетки взаимодействуют с помощью
химических веществ – нейромидиаторов.
Активность
нейрона может быть индуцирована импульсом
от другого нейрона или быть спонтанной.
В этом случае нейрон играет роль
пейсмекера (водителя ритма). Такие
нейроны имеются в ряде центров, в том
числе дыхательном.
Первым воспринимающим
нейроном в рефлекторной дуге является
чувствительная клетка. Раздражение
воспринимается рецептором – чувствительным
окончанием, по дендриту импульс достигает
тела клетки, а затем передается по аксону
на другой нейрон. Команда к действию на
рабочий орган передается двигательным
или эффекторным нейроном. Эффекторный
нейрон может получить импульс
непосредственно от чувствительной
клетки, тогда рефлекторная дуга будет
состоять из двух нейронов.
В более сложных
рефлекторных дугах есть среднее звено
– вставочный нейрон. Он воспринимает
импульс от чувствительной клетки и
передает на двигательную.
Иногда несколько
клеток с одинаковой функцией (чувствительные
или двигательные) объединяются одним
нейроном, который концентрирует в себе
импульсы с нескольких клеток – это
ассоциативные невроны. Эти нейроны
передают импульс дальше на вставочные
или на эффекторные нейроны.
В теле нейрона у
большинства нервных клеток содержится
одно ядро. Многоядерные нервные клетки
свойственны некоторым периферическим
ганглиям вегетативной нервной системы.
На гистологических препаратах ядро
нервной клетки имеет вид светлого
пузырька с четко различимым ядрышком
и немногочисленными глыбками хроматина.
При электронной микроскопии обнаруживаются
те же субмикроскопические компоненты,
что и в ядрах других клеток. Ядерная
оболочка имеет многочисленные поры.
Хроматин распылен. Такая структура ядра
характерна для активных в метаболическом
отношении ядерных аппаратов.
Ядерная оболочка
в процессе эмбриогенеза образует
глубокие складки, заходящие в кариоплазму.
К моменту рождения складчатость
становится значительно меньше. У
новорожденного наблюдается уже
преобладание объема цитоплазмы над
ядром, так как в период эмбриогенеза
эти отношения обратные.
Цитоплазма нервной
клетки носит название нейроплазмы. В
ней располагаются органоиды и включения.
Аппарат Гольджи
был впервые обнаружен в нервных клетках.
Он имеет вид сложной корзинки, окружающей
ядро со всех сторон. Это своеобразный
диффузный тип аппарата Гольджи. При
электронной микроскопии он состоит из
крупных вакуолей, мелких пузырьков и
пакетов двойных мембран, образующих
анастомозирующую сеть вокруг ядерного
аппарата нервной клетки. Однако чаще
всего аппарат Гольджи располагается
между ядром и местом отхождения аксона
– аксонный холмик. Аппарат Гольджи
является местом генерации потенциала
действия.
Митохондрии имеют
вид очень коротких палочек. Они
обнаруживаются в теле клетки и во всех
отростках. В концевых разветвлениях
нервных отростков, т.е. в нервных
окончаниях наблюдается их скопление.
Ультраструктура митохондрий типична,
но их внутренняя мембрана не образует
большого количества крист. Они очень
чувствительны к гипоксии. Впервые
митохондрии описал в мышечных клетках
Келликер более 100 лет назад. В некоторых
нейронах между кристами митохондрий
имеются анастамозы. Количество крист
и их общая поверхность прямо связаны с
интенсивностью их дыхания. Необычным
является накопление митохондрий в
нервных окончаниях. В отростках они
ориентируются своей продольной осью
по ходу отростков.
Клеточный центр
в нервных клетках состоит из 2-ух
центриолей, окруженных светлой сферой,
и выражен в молодых нейронах значительно
лучше. В зрелых нейронах клеточный центр
обнаруживается с трудом и во взрослом
организме центросома претерпевает
дегенеративные изменения.
При окрашивании
нервных клеток толуоидным синим в
цитоплазме обнаруживаются глыбки
различных размеров – базофильное
вещество, или субстанция Ниссля.Это
очень нестойкое вещество: при общей
усталости в следствии длительной работы
или нервного возбуждения глыбки вещества
Ниссля исчезают. Гистохимически в
глыбках была обнаружена РНК и гликоген.
Электронно-микроскопические исследования
показали, что глыбки Ниссля представляют
собой эндоплазматическую сеть. На
мембранах эндоплазматической сети
много рибосом. В нейроплазме так же
много и свободных рибосом, образующих
розеткообразные скопления. Развитая
гранулярная эндоплазматическая сеть
обеспечивает синтез большого количества
белка. Синтез белка наблюдается только
в теле нейрона и в дендритах. Для нервных
клеток характерен высокий уровень
синтетических процессов и в первую
очередь белку и РНК.
В сторону аксона
и по аксону наблюдается постоянный
токполужидкого содержимого нейрона,
движущегося на периферию нейрита со
скоростью 1-10 мм в сутки. Помимо медленного
перемещения нейроплазмы обнаружен и
быстрый ток(от 100 до 2000 мм в сутки), он
имеет универсальный характер. Быстрый
ток зависит от процессов окислительного
фосфорилирования, наличия кальция и
нарушается при разрушении микротрубочек
и нейрофиламентов. Быстрым транспортом
переносятся холинэстераза, аминокислоты,
митохондрии, нуклеотиды. Быстрый
транспорт тесно связан с подачей
кислорода. Через 10 минут после смерти
прекращается движение в периферическом
нерве млекопитающих. Для патологии
существование аксоплазматического
движения имеет значение в том смысле,
что по аксону могут распространяться
различные инфекционные агенты, как из
периферии организма в центральную
нервную систему, так и внутри ее.
Непрерывный аксоплазматический транспорт
является активным процессом, требующим
затрат энергии. Некоторые вещества
обладают способностью перемещаться по
аксону в обратном направлении (ретроградный
транспорт): ацетилхолинэстераза,
вирус полиомиэлита, вирус герпеса,
столбнячный токсин, который вырабатывается
бактериями, попавшими в кожную рану, по
аксону достигает центральной нервной
системы и вызывает судороги.
У новорожденного
нейроплазма бедна глыбками базофильного
вещества. С возрастом наблюдается
увеличение числа и размеров глыбок.
Специфическими
структурами нервных клеток являются
также нейрофибриллы и микротрубочки.
Нейрофибриллыобнаруживаются в
нейронах при фиксации и в теле клетки
имеют беспорядочное расположение в
виде войлока, а в отростках лежат
параллельно друг другу. В живых клетках
они были найдены при помощи фазово-контрольной
киносъёмки.
При электронной
микроскопии в цитоплазме тела и отростков
находят гомогенные нити нейропротофибриллы,
состоящие из нейрофиламентов.
Нейрофиламенты это фибриллярные
структуры диаметром от 40 до 100 А. Они
состоят из спирально закрученных нитей,
представленных белковыми молекулами
весом 80000. Нейрофибриллы возникают при
пучковой агрегации существующих
прижизненно нейропротофибрилл. Одно
время нейрофибриллам приписывали
функцию проведения импульсов, но
оказалось, что после перерезки нервного
волокна проводимость сохраняется даже
тогда, когда нейрофибриллы уже
дегенерируют. Очевидно, основная роль
в процессе проведения импульса принадлежит
межфибриллярной нейроплазмы. Таким
образом, функциональное значение
нейрофибрилл не ясно.
Микротрубочкипредставляют собой цилиндрические
образования. Их сердцевина обладает
низкой электронной плотностью. Стенки
образованы 13 ориентированными продольно
фибриллярными субъединицами. Каждая
фибрилла в свою очередь состоит из
мономеров, которые агрегируют и образуют
вытянутую фибриллу. Большинство
микротрубочек располагается в отростках
продольно. По микротрубочкам осуществляется
транспорт веществ (белков, нейромедиаторов),
органоидов (митохондрий, везикул),
ферменты синтеза медиаторов.
Лизосомы в
нервных клетках мелкие, их мало, и
структуры их не отличаются от других
клеток. Они содержат высоко активную
кислую фосфотазу. Лизосомы лежат в
основном в теле нервных клеток. При
дегенеративных процессах, в нейронах
число лизосом возрастает.
В нейроплазме
нервных клеток обнаруживаются включения
пигмента и гликогена. В нервных клетках
находят два вида пигментов – это
липофусцин, имеющий бледно-жёлтый или
зеленовато-жёлтый цвет, и меланин –
пигмент тёмно-бурого или коричневого
цвета (например, черное вещество
–substantianigraв ножках мозга).
Меланинобнаруживается в клетках очень рано –
к концу первого года жизни.Липофусцин
накапливается
позднее, но к 30 годам он может быть
выявлен почти во всех клетках. Пигменты
типа липофусцина играют важную роль в
обменных процессах. Пигменты относящиеся
к хромотопротеидам, являются катализаторами
в окислительно-восстановительных
процессах. Они являются древней
окислительно-восстановительной системой
нейроплазмы.
Гликоген
накапливается, в нейроне в период
относительного покоя в областях
распространения вещества Ниссля.
Гликоген содержится в телах и проксимальных
отрезках дендритов. Аксоны лишены
полисахаридов. В нервных клетках
содержатся и ферменты: оксидаза, фосфатаза
и холинэстераза. Специфическим белком
аксоплазмы является нейромодулин.
Соседние файлы в папке все лекции по гисте
- #
- #
- #
- #
- #
- #
- #
- #
- #
Основная статья: Тело человека
Строение нейронов
Клетка нейрона содержит ядро, тело клетки и отростки (дендриты и аксоны). Если можно так сказать, то нейрон — это самая «отросчатая» клетка в человеческом организме, так как его дендриты и аксоны растягиваются словно провода, соединяя нейрон с другими нейронами.

![]()
Так, нейроны соединяются один с другим, формируя биологические нейронные сети. По сути, циркуляция возбуждения в этих сетях — это и есть наше мышление, наши чувства, это и есть мы.
Нейроны бывают разного размера: от 4 до 100 мкм в ширину.
Чтобы представить их размер, сравните с точкой в этом тексте, её размер примерно 500 микрон, т.е. в одной точке может содержаться около 100 нервных клеток.
Дивергенция — способность одиночного нейрона устанавливать многочисленные связи
Дивергенция — способность одиночного нейрона устанавливать многочисленные связи с различными нервными клетками.
Ниже на видео показано формирование нейронных систем. Рост живой нейронной сети.
Таким образом, каждый нейрон может обеспечивать широкое распространение импульса из одной точки на целый орган или систему. Также, благодаря процессу дивергенции одна и та же клетка может участвовать в организации различных реакций и контролировать большее число нейронов.
В ходе исследований был снят процесс изменения структуры нейронных связей в живом организме:
Синапс — соединение между нейронами
2023: Впервые созданы зрелые нейроны из стволовых клеток
12 января 2023 года американские исследователи из Северо-Западного университета (штат Иллинойс) сообщили о получении первых высокозрелых нейронов из индуцированных плюрипотентных стволовых клеток (ИПСК). Это открывает новые возможности по терапии нейродегенеративных заболеваний.
В ходе предыдущих работ учёные уже дифференцировали стволовые клетки в нейроны, но они были функционально незрелыми и напоминали нейроны на ранних стадиях развития. Ограниченное созревание, которое обеспечивают современные методы культивирования стволовых клеток, снижает потенциал для исследований в области нейродегенерации. Решение проблемы предложили специалисты Северо-Западного университета.

![]()
Впервые созданы зрелые нейроны из стволовых клеток
Команда сначала дифференцировала ИПСК человека в моторные и кортикальные нейроны, а затем поместила их на покрытия из синтетических нановолокон, содержащих быстро движущиеся «танцующие» молекулы. Предложенная технология решает сразу несколько важных задач. Она позволяет получать более зрелые нейроны, которые демонстрируют улучшенную электрическую активность. Кроме того, такие нейроны более расположены к установлению синаптических связей. Вдобавок они меньше слипаются по сравнению с типичными нейронами, происходящими от стволовых клеток.
Зрелые нейроны, по мнению исследователей, открывают совершенно новые возможности для изучения нейродегенеративных заболеваний, таких как боковой амиотрофический склероз, болезнь Паркинсона, болезнь Альцгеймера или рассеянный склероз. Кроме того, нейроны могут быть трансплантированы пациентам с травмами спинного мозга.
|
|
Мы подтвердили, что нейроны, покрытые нашими нановолокнами, достигают большей зрелости по сравнению с другими методами. Зрелые нейроны способны лучше устанавливать синаптические связи, которые имеют основополагающее значение для их функционирования, — отметил профессор Сэмюэль И. Ступп (Samuel I. Stupp), соавтор исследования. [1] |
|
Как происходит передача нервного импульса от одного нейрона к другому

![]()
Микрофотография вскрытого нервного окончания. Можно увидеть везикулы с нейромедиатором (синие, фиолетовые и оранжевые шарики).
Вопреки расхожему мнению, передача нервного импульса вовсе не электрический процесс (в отличие от проведения импульса). Всё это — химический процесс, который осуществляется благодаря медиаторам.
Нейромедиаторы — короткоживущие вещества локального действия; они выделяются в щель между нейронами и передают сигнал соседним клеткам. Всё это соединение двух нейронов называется синапс.

![]()
Так выглядит синапс — соединение между нейронами. В данный момент через синаптическую щель происходит транспорт молекул активного вещества для передачи возбуждения на другой нейрон.
Зачем организму необходимы такие синапсы? Почему нельзя просто непрерывно передавать импульс? На 100 процентов пока ещё нельзя сказать, однако, понятно одно — подобные щели позволяют лучше контролировать и направлять нервные процессы в нашем организме. Вместо простого проведения импульса из точки А в точку Б, наш мозг как бы расставляет «блокпосты», на которых он может затормозить импульс (сделать рецепторы нечувствительными к нейромедиатору) или же, например, ускорить его (путём увеличения количества медиатора в щели).
Весь процесс транспортировки нейромедиатора через синаптическую щель в анимации ниже:

![]()
Кора мозжечка в 3D, где красными дендритными деревьями раскинулись среди прочих нервных клеток ветвистые нейроны Пуркинье, способные образовывать до 100 тысяч соединений (синапсов).
Нервные импульсы перемещаются со скоростью 402 километра в час
Ощущение боли, по сути, наступает мгновенно, когда вы касаетесь чего-то горячего или укалываетесь иголкой. Благодаря нашей высокоразвитой нервной системе человек может реагировать менее чем за миллисекунду на вещи, от которых следует держаться подальше.
Нервные импульсы настолько быстрые, что электрические сигналы, отвечающие за ощущения, путешествуют в мозг и из мозга со средней скоростью 402 километра в час.
Миелин предотвращает рассеивание нервных импульсов
Миелиновая оболочка — электроизолирующая жирная белая оболочка, покрывающая аксоны многих нейронов.

![]()
Миелин нарастает на наиболее часто используемых путях в мозге. Нейроны с миелином передают электрические сигналы в 10 раз быстрее, чем нейроны без
миелина.
Миелин прерывается только в области перехватов Ранвье, которые встречаются через правильные промежутки длиной 0.2 mm – >1 mm. В связи с тем, что ионные токи не могут проходить сквозь миелин, вход и выход ионов осуществляется лишь в области перехватов. Это ведёт к увеличению скорости проведения нервного импульса. Таким образом, по миелинизированным волокнам импульс проводится приблизительно в 5—10 раз быстрее, чем по немиелинизированным.
Миелиновую оболочку образуют глиальные клетки: в периферической нервной системе — Шванновские клетки, в центральной нервной системе — олигодендроциты. Миелиновая оболочка формируется из плоского выроста тела глиальной клетки, многократно оборачивающего аксон подобно изоляционной ленте.
Общая длина волокон миелина человеческого мозга (включая и волокна, что соединяют между собой полушария, формируя так называемое мозолистое тело) оценивается в примерно 150 тысяч километров. Это равно четырем окружностям земного экватора.
Как нейроны анализируют поступающие импульсы
Каждый из нейронов способен принимать сотни сообщений в секунду
Чтобы не оказаться перегруженным информацией, он должен уметь судить о степени ее значимости и делать ее предварительный анализ.
Эта вычислительная деятельность происходит внутри клетки. Там складываются возбуждающие и вычитаются тормозящие импульсы.
И для того чтобы нейрон сгенерировал собственный импульс, необходимо, чтобы сумма предыдущих оказалась больше определенного значения.
Если сложение возбуждающих и тормозящих импульсов не превысит этот предел, нейрон будет «молчать».
Образование новых нейронов
Нейрогенез и синаптический прунинг
Мозг обладает способностью к нейрогенезу, то есть к образованию новых нейронов, только во время внутриутробного развития, до первых месяцев жизни.
Однако в первые три года жизни образует максимальное количество синапсов. Согласно некоторым исследованиям, у малыша трех лет от роду в мозге действует около миллиона миллиардов контактов: каждый нейрон вступает в контакт с другим не менее 15 тысяч раз.
У взрослого сохраняется примерно половина этих соединений. Очень любопытный выбор эволюции: вместо того чтобы накапливать связи, она предпочла создать их избыток, чтобы потом спокойно пожертвовать лишними.
Этот процесс называется синаптический прунинг.
Изучение нейрогенеза (образования новых нервных клеток — нейронов) — относительно новое направление исследований. За последние годы было доказано, что новые нейроны на протяжении всей жизни образуются в мозге многих млекопитающих, однако по вопросу о нейрогенезе у человека консенсуса в научном сообществе к 2019 году до сих пор нет.
Новые методы визуализации (такие, как конфокальная микроскопия), позволили доказать, что по крайней мере до полового созревания новые нейроны образуются в человеческом гиппокампе — области мозга, участвующей в формировании эмоций и памяти.
Исследования показывают, что в зубчатой фасции (части мозга, где происходит нейрогенез) имеются тысячи молодых, не до конца оформившихся нейронов во всех пробах, вне зависимости от возраста людей. Однако чем старше человек, тем меньше в зубчатой фасции клеток, вырабатывающих вещества, которые связаны со способностью мозга к перестройке существующих нейронных связей и образованию новых.
В заключении можно сказать, что всё-таки во взрослом возрасте также появляются новые нейроны, однако они образуют меньше связей друг с другом и другими нейронами, или реже мигрируют в другие отделы мозга, так что мы не можем назвать это полной регенерацией.
Влияние алкоголя на нейроны плода
В умеренных дозах алкоголь не убивает взрослые нейроны, но он может оказывать сильное воздействие на развивающиеся нервные клетки.
Поскольку почти все нейроны формируются и перемещаются на свои места еще до рождения, мозг плода очень восприимчив к алкоголю.
Алкоголь может убить недавно появившиеся на свет нейроны, воспрепятствовать их рождению и помешать их перемещению от места рождения на место конечного пребывания.
Даже кратковременного увеличения уровня алкоголя в крови бывает достаточно, чтобы некоторые нервные клетки плода погибли.
Глиальные клетки — клей для нейронов
Глиальные клетки действительно, как думали ученые раньше, играют роль клея – они окружают нейроны и удерживают их в определенном месте.
Кроме того, они поставляют нейронам топливо – питательные вещества и кислород – и работают электриками, выстраивая миелиновые оболочки, регулирующие передачу потенциала вдоль аксонов.
Освоили глиальные клетки и профессию дворников – они задерживают патогены и устраняют нейроны, прекратившие всякую активность.
Без этих важных функций глиальных клеток человеческий мозг не мог бы функционировать столь эффективно, как сейчас.
Как мы теряем нервные клетки с самого рождения
Сколько нейронов (нервных клеток) в мозге человека? У нас их около 85 миллиардов. Для сравнения, у медузы — всего 800, у таракана — миллион, а у осьминога — 300 млн.
Многие считают, что нервные клетки гибнут лишь в пожилые годы, но большая их часть теряется нами еще в детстве, когда в голове ребенка происходит процесс естественного отбора.
Как в джунглях, среди нейронов выживают наиболее эффективные и приспособленные.
Правда ли, что мозг человека работает как компьютер?
Основная статья: Мозг человека
Это не совсем так, ведь мозг человека гораздо мощнее. Компьютер работает последовательно, а мозг человека параллельно. Это связано с тем, что нейроны выполняют одновременно все функции компьютера — запоминание, воспроизведение, хранение.
Одна ячейка памяти компьютера может иметь только одно из двух значений, а мозг устроен гораздо сложнее в этом плане. У нейронов есть так называемые шипики — отростки, которые и отвечают за соединения и получение связей. Это прямой аналог нуля и единицы в ячейке данных памяти компьютера. Один нейрон может иметь более 20 соединений. Это говорит о том, что наш мозг настолько совершенен, что компьютеры не смогут приблизиться к нему по уровню производительности, скорее всего, никогда.
Роль нейронов в работе памяти
Физиологической основой памяти являются «следы» ранее бывших нервных процессов, сохраняющихся в мозге. Любой вызванный внешним раздражением нервный процесс (например, передача изображения какого-то рисунка в мозг), не проходит для нервной ткани бесследно, а оставляет в ней как бы «след» в виде определенных функциональных изменений. Таким образом, при восприятии определенной информации, между некоторыми группами нейронов образуется связь, которая и кодирует эту инфорамцию. И чем чаще данная информация поступает в мозг, тем чаще нервный импульс проходит по связи и тем больше связь «закрепляется».
Когда мы увидим, например, рисунок еще раз, то нервный импульс пройдет по знакомому пути и связь между определенными нейронами станет еще сильнее и так далее.
Согласно последним исследованиям, материальным носителем информации о разных событиях является не возбуждение разных нейронов, а различные комплексы нейронных сетей, которые и образуются в момент восприятия информации.
Ниже запись эксперимента на эту тему: здесь нейроны образуют между собой новые связи прямо в пробирке.
Нейроны, специализирующиеся на распознавании лиц
В зрительной зоне мозга существуют нейроны, специализирующиеся на распознавании лиц. Они анализируют черты лица, в первую очередь – форму глаз, и относят их к категории «человеческие лица».
После этого они сравнивают эти образы с образами, хранящимися в памяти. В результате, мы можем быстро распознавать лица наших родственников и знакомых.
При поражении этой зоны мозга возникает невозможность распознавать лица, даже самые знакомые, при том, что и зрение и распознавание других объектов может оставаться на нормальном уровне.
Чтение мыслей
Основная статья: Чтение мыслей
Искусственные нейроны
2023: Разработаны искусственные нейроны, способные подключаться к живым в организме человека
В середине января 2023 года появилась информация о том, что исследователи из Университета Линчепинга в Швеции разработали искусственные нейроны, которые демонстрируют 15 из 20 характеристик биологических нейронных клеток и могут общаться с естественными нейронами в организме. Ученые назвали свое устройство «органическим электрохимическим нейроном на основе проводимости» (c-OECN).
В основе c-OECN лежат материалы, способные проводить отрицательный заряд, включая органические электрохимические транзисторы и проводящие полимеры n-типа. Напечатав тысячи таких транзисторов на гибкой подложке, исследователи смогли создать искусственные нейроны. Устройство использует ионы для управления потоком электричества, подобно биологическим нейронам, и на данный момент шведская команда продемонстрировала, что оно может управлять блуждающим нервом у мышей, что позволяет предположить, что оно имеет большой потенциал для применения в медицине.

![]()
Искусственный нейрон c-OECN
Искусственный нейрон c-OECN является примером, поскольку он использует ионы для управления электричеством, проходящим через него, подобно тому, как это делают биологические нейроны, открывая и закрывая ионные каналы. Ученые надеются, что эти искусственные нейроны могут помочь в создании более реалистичного нейронного контроля в различных медицинских технологиях. Искусственные нейроны используют ионы для управления электричеством, проходящим через проводящий полимер внутри них, что приводит к скачкам напряжения, имитирующим то, что происходит в настоящих нейронах. Технология позволяет устройству контролируемо увеличивать и уменьшать ток в виде почти идеальной колоколообразной кривой, что напоминает электрическую активность в нейронах, которая контролируется натриевыми ионными каналами.
На январь 2023 года исследователи протестировали технологию на мышах и имплантировали напечатанные нейроны с целью управления блуждающим нервом. Нейроны успешно взаимодействовали с блуждающим нервом, что привело к снижению частоты сердечных сокращений у мышей на 4,5%.[2]
Исследования
1860: Первое описание нейрона Отто Дейтерса

![]()
Этот рисунок сделал в 1860 году молодой немецкий анатом Отто Дейтерс. На тот момент ему было всего 26 лет.
Это — первое в истории полноценное описание классического нейрона, в котором были выделены дендриты и аксон.
Отто Дейтерс умер от тифа всего в 29 лет, однако успел сделать в нейронанатомии очень много.
Технологии для исследования нейронов
2023: Российские ученые создали технологию клеточной томографии, которая ускорит процесс выявления болезней
Технологию клеточной томографии предложил коллектив исследователей из НИТУ МИСИС, МГУ имени М.В. Ломоносова и ВНИИОФИ. Она позволит преодолеть ограниченность методов фазовой и абсорбционной микроскопии, которые анализируют лишь единичную клетку правильной формы. В будущем с помощью локального томографа планируется изучение субклеточных структур и цитоплазмы при функционировании нейрона, что приблизит ученых к пониманию того, как работает человеческий мозг, сообщили 9 марта 2023 года Zdrav.Expert представители НИТУ МИСиС. Подробнее здесь.
Нейроны
Нейрон (нервная клетка) – это структурно-функциональная единица нервной системы. Эта клетка имеетсложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. Ворганизме человека насчитывается более ста миллиардов нейронов.
Обзор
Сложность и многообразие нервной системы зависит от взаимодействия между нейронами, которые, в своюочередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов сдругими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
Строение
Тело клетки
Нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро (с большим количеством ядерных пор) и другие органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппаратГольджи), и отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» длятранспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). В теленейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно иизвестна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается назаметном расстоянии от начала аксона, что служит гистологическим признаком аксона.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Схема строения нейрона
Аксон обычно — длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образованиявлияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношениедлины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Одиннейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточенымитохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения убольшинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всехнейронов эта зона называется триггерной.
Cинапс
Си́напс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторнойклеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптическойпередачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризациюнейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозящими. Обычнодля возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Классификация
Структурная классификация
На основании числа и расположения дейндритов и аксона нейроны делятся на безаксонные, униполярныенейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночныхганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки уклетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничногонерва в среднем мозге.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные вспециализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом ивестибулярном ганглиях;
Мультиполярные нейроны — Нейроны с одним аксоном и несколькими дендритами. Данный вид нервныхклеток преобладает в центральной нервной системе
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один остросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурнопредставляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурнодендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной являетсяначало этого разветвления (т. е. находится вне тела клетки).
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точноеназвание распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятсяпервичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободныеокончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятсяконечные нейроны — ультиматные и предпоследние – неультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — эта группа нейронов осуществляет связь междуэфферентными и афферентными, их делят на комисуральные и проекционные (головной мозг).
Морфологическая классификация
Нервные клетки бывают звездчатые и веретенообразные, пирамидальные, зернистые, грушевидные и т.д.
Развитие и рост нейрона
Конус роста
Нейрон развивается из небольшой клетки — предшественницы, которая перестаёт делиться ещё до того, каквыпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным. [1](рус.)) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегосяотростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путьчерез окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит изуплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину идлину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса ростапокрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются вконус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать кнему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильнойформы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная массаперепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки инейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновьсинтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью околомиллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время ростаотростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек инейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — этообласть быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потокомбыстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится кконусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняятаким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейронырасселяются и находят себе постоянное место.
Источник: http://kineziolog.su/content/kholinergicheskaya-sistema-regulyatsii
Записаться онлайн или получить расписание ближайших мастер классов можно по ссылке: https://qviz.matomba.ru/1c06710015fbab389df6
Глава 3. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ НЕРВНО-ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ
§ 3.1. НЕЙРОН КАК СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА НЕРВНОЙ СИСТЕМЫ
Структурно-функциональной единицей нервной системы является нервная клетка, или нейрон, или нейроцит. Всю нервную систему можно представить как взаимосвязанную и взаимодействующую сеть из нескольких триллионов нервных клеток. Несмотря на их громадное разнообразие, можно говорить о ряде общих структурных и функциональных признаков, присущих всем нервным клеткам (рис. 5).
В нейроне выделяют следующие основные части: тело, отростки и их окончания.
Тело нейрона, размеры которого колеблются от 4 до 130 мкм, представляет собой скопление клеточной плазмы, в которой располагается ядро — носитель генетической информации, митохондрии — универсальные «генераторы» энергии, необходимой для обеспечения деятельности клетки, и большое количество структур, выполняющих различные специфические функции.
Поверхность нейрона, его оболочка, часто именуемая просто мембраной, не только обеспечивает обмен с окружающей средой, но, обладая свойствами полупроницаемой мембраны, является структурой, где развиваются сложные процессы биоэлектрогенеза, лежащие в основе главных функций нервной клетки.
Отростки нервных клеток являются выростами цитоплазмы. Различают два вида отростков. Дендриты — короткие, древовидно ветвящиеся, постепенно истончаются и заканчиваются в окружающих тканях. Количество их достигает десяти, они многократно увеличивают поверхность клетки.
Помимо дендритов нервная клетка всегда имеет один аксон (или нейрит). Этот отросток всегда более крупный, длинный (до 1 м) и менее ветвистый. Аксон заканчивается синапсом, при помощи которого он функционально взаимодействует с иннервируемыми структурами.

Рис. 5. Строение нервной клетки (упрощенная схема)
По своей функциональной значимости в составе рефлекторной дуги различают три вида нейронов:
рецепторные (чувствительные, афферентные), имеющие чувствительные нервные окончания, которые способны воспринимать раздражения из внешней или внутренней среды;
эффекторные (эфферентные), окончания аксонов которых передают нервный сигнал на рабочий орган;
ассоциативные (вставочные, центральные), являющиеся промежуточными в составе рефлекторной дуги и передающие информацию с чувствительного нейрона на эффекторные.
Следует иметь в виду, что на теле и отростках большинства нервных клеток имеется очень большое количество синапсов, через которые поступает информация с других нейронов.
Несмотря на громадное морфологическое и функциональное разнообразие нейронов, можно выделить ряд ключевых свойств и функций.
К числу наиболее важных свойств относятся:
1. Наличие трансмембранной разности потенциалов, т. е. между наружной и внутренней поверхностями оболочки нейрона в покое регистрируется разность потенциала порядка 90 мВ, наружная поверхность электроположительна по отношению к внутренней. Величина и направление трансмембранного тока меняются в зависимости от состояния нейрона.
2. Очень высокая чувствительность к некоторым химическим веществам (медиаторам) и электрическому току.
3. Способность к нейросекреции, т. е. к синтезу и выделению в окружающую среду или в синаптическую щель биологически активных веществ.
4. Высокий уровень энергетических процессов, что обусловливает необходимость постоянного притока основного источника энергии — глюкозы и кислорода, необходимого для окисления.
Принято различать следующие функции нейрона:
1. Воспринимающая — эта функция представлена двумя механизмами. Во-первых, чувствительные окончания дендритов способны обеспечить рецепцию, т. е. трансформацию специфической энергии раздражителя внешней или внутренней среды в неспецифический процесс нервного возбуждения, нервный импульс, который по отростку распространяется по направлению к телу нервной клетки. Во-вторых, на всех частях нейрона имеются многочисленные (до нескольких десятков тысяч) синапсы, при помощи которых химическим путем возбуждение передается от одного нейрона к другому. Химические вещества, осуществляющие эту передачу, обозначают медиаторы (или нейротрансмиттеры). К их числу, в частности, относятся адреналин, норадреналин, дофамин, серотонин, ацетилхолин, гамма-аминомасляная кислота и многие другие. В результате воздействия медиатора в теле нервной клетки развивается возбуждение и возникновение нервного импульса или снижение возбудимости нейрона — его торможение.
2. Интегративная — обработка одновременно или в течение короткого интервала времени поступающих нервных сигналов по механизму их алгебраической суммации, в результате которой на выходе нейрона формируется сигнал, несущий в себе информацию всех суммированных сигналов.
3. Мнестическая, основанная на существовании тонких молекулярных биофизических процессов, сохраняющих след от всякого предыдущего воздействия и благодаря этому трансформирующих характер ответной реакции на всякое последующее. По существу, это элементарная форма памяти и научения.
4. Проводниковая функция, суть которой состоит в том, что от тела нейрона по аксону к его окончанию в естественных условиях только в одном этом направлении распространяется, не затухая, нервный импульс. Скорость его распространения в зависимости от морфофункциональных особенностей проводника колеблется от нескольких сантиметров до 100–120 метров в секунду.
Психология bookap
5. Передающая, проявляющаяся в том, что нервный импульс, достигнув окончания аксона, который, собственно, уже входит в структуру синапса, обусловливает выделение медиатора — непосредственного передатчика возбуждения к другому нейрону или исполнительному органу.
Часто в бытовых разговорах приходится слышать сожалеющее высказывание, что нервные клетки не восстанавливаются. Да, применительно к телу нейрона, это действительно так, и в ряде случаев это действительно плохо. Но следует также иметь в виду, что количество нейронов у человека значительно превышает его потребности на протяжении всей жизни. И, кроме того, как указывалось выше, нервные клетки на протяжении жизни человека «обучаются», «приобретают опыт», а потому включение в слаженный нейрональный ансамбль «необученного» элемента затруднило бы его работу.
Нервная ткань — основная ткань, формирующая нервную систему и создающая условия для реализации ее многочисленных функций. Нервная ткань имеет эктодермальное происхождение, не принято делить нервную ткань на какие-либо виды тканей. Обладает двумя основными свойствами: возбудимостью и проводимостью.
Нейрон
Структурно-функциональной единицей нервной ткани является нейрон (от др.-греч. νεῦρον — волокно, нерв) — клетка с одним
длинным отростком — аксоном (греч. axis — ось), и одним/несколькими короткими — дендритами (греч. dendros — дерево).
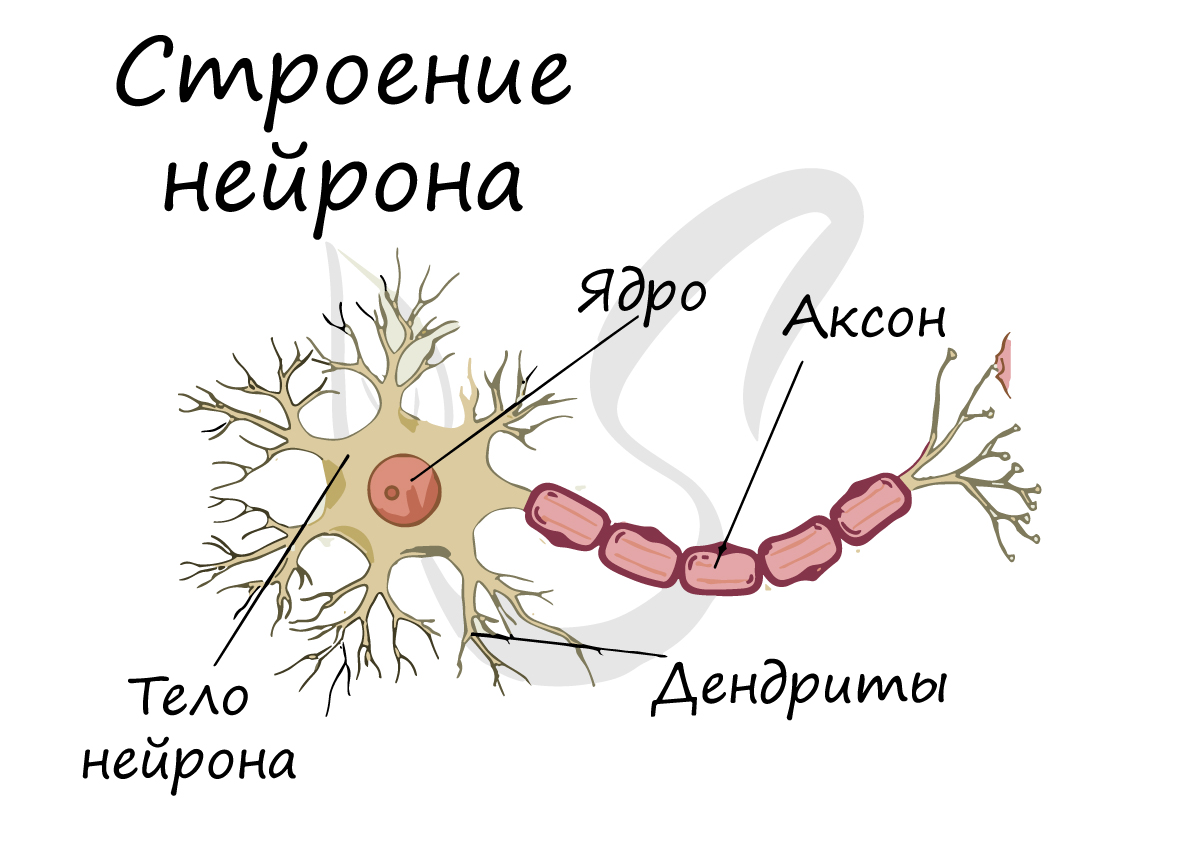
Спешу сообщить, что представление, будто короткий отросток нейрона — всегда дендрит, а длинный — всегда аксон, в корне неверно. С точки
зрения физиологии правильнее дать следующие определения: дендрит — отросток нейрона, по которому нервный импульс перемещается к телу нейрона, аксон — отросток нейрона, по которому импульс перемещается от тела нейрона.
Нейроны обладают 4 свойствами:
- Рецепция (лат. receptio — принятие) — способны воспринимать поступающие сигналы (дендриты)
- В ответ на сигналы способны переходить в состояние возбуждения или торможения
- Проведение возбуждения (от дендрита к телу нейрона, затем — к концу аксона)
- Передача сигнала другим объектам — нейрону или эффекторному органу
В физиологии эффекторным (от лат. efferes — выносящий) органом часто называют исполнительный орган или орган-мишень воздействия (мышцы, железы). Орган-эффектор выполняет те или иные «приказы» ЦНС (центральной нервной системы) или эндокринных желёз
Отростки нейронов проводят нервные импульсы и передают их другим нейронам, эффекторам, благодаря чему
мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
Миелиновая оболочка
Нервные волокна подразделяются на миелиновые и безмиелиновые. Нервное волокно — это один или несколько отростков нейронов (могут быть как аксоны, так и дендриты) с окружающей оболочкой.
Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы (скорость проведения 1-2 м/c). Миелиновые — образуют белое вещество головного и спинного мозга, нервные волокна соматической нервной системы (5-120 м/с).
В миелиновых нервных волокнах отростки нейронов покрыты миелиновой оболочкой (на 70-75% состоит из липидов (жиров)), которая обеспечивает изолированное проведение нервного
импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и,
когда мы хотели сделать движение рукой, то вместе с рукой двигалась бы нога.
Существует болезнь при которой собственные антитела уничтожают миелиновую оболочку нервных волокон головного и спинного мозга (случаются и такие сбои в работе организма). Эта
болезнь — рассеянный склероз, по мере прогрессирования приводит к разрушению не только миелиновой оболочки, но и нервов — а значит,
происходит атрофия мышц и человек постепенно становится обездвиженным.

Миелиновый слой представлен несколькими слоями мембраны глиальной клетки (леммоцит, шванновская клетка), которые закручиваются вокруг осевого цилиндра (отростка нейрона). Это закручивание хорошо видно на картинке, где изображен здоровый нерв, чуть выше 
Миелиновый слой оболочки волокна регулярно прерывается в местах стыка соседних леммоцитов — перехваты Ранвье. Миелиновая оболочка обеспечивает изолированное и более быстрое проведение возбуждения (сальтаторный тип, лат. salto — скачу, прыгаю).

Нейроглия (греч. νεῦρον — волокно, нерв + γλία — клей)
Вы уже убедились, насколько значимы нейроны, их высокая специализация приводит к возникновению особого окружения — нейроглии.
Нейроглия (глиальные клетки, глиоциты) — вспомогательная часть нервной системы, которая выполняет ряд важных функций:
- Опорная — поддерживает нейроны в определенном положении
- Регенераторная (лат. regeneratio — возрождение) — в случае повреждения нервных структур нейроглия способствует регенерации
- Трофическая (греч. trophe — питание) — с помощью нейроглии осуществляется питание нейронов: напрямую с кровью нейроны не контактируют
- Электроизоляционная — леммоциты (шванновские клетки) закручиваются вокруг отростков нейронов и формируют миелиновую оболочку
- Барьерная и защитная — изолируют нейроны от тканей внутренней среды организма
- Некоторые глиоциты секретируют цереброспинальную (спинномозговую) жидкость — ликвор (от лат. liquor — жидкость)
В состав нейроглии входят разные клетки, их в десятки раз больше чем самих нейронов. В периферическом отделе нервной
системы миелиновая оболочка, изученная нами, образуется именно из нейроглии — шванновских клеток (леммоцитов). Между ними хорошо
заметны перехваты Ранвье — участки, лишенные миелиновой оболочки, между двумя смежными шванновскими клетками.
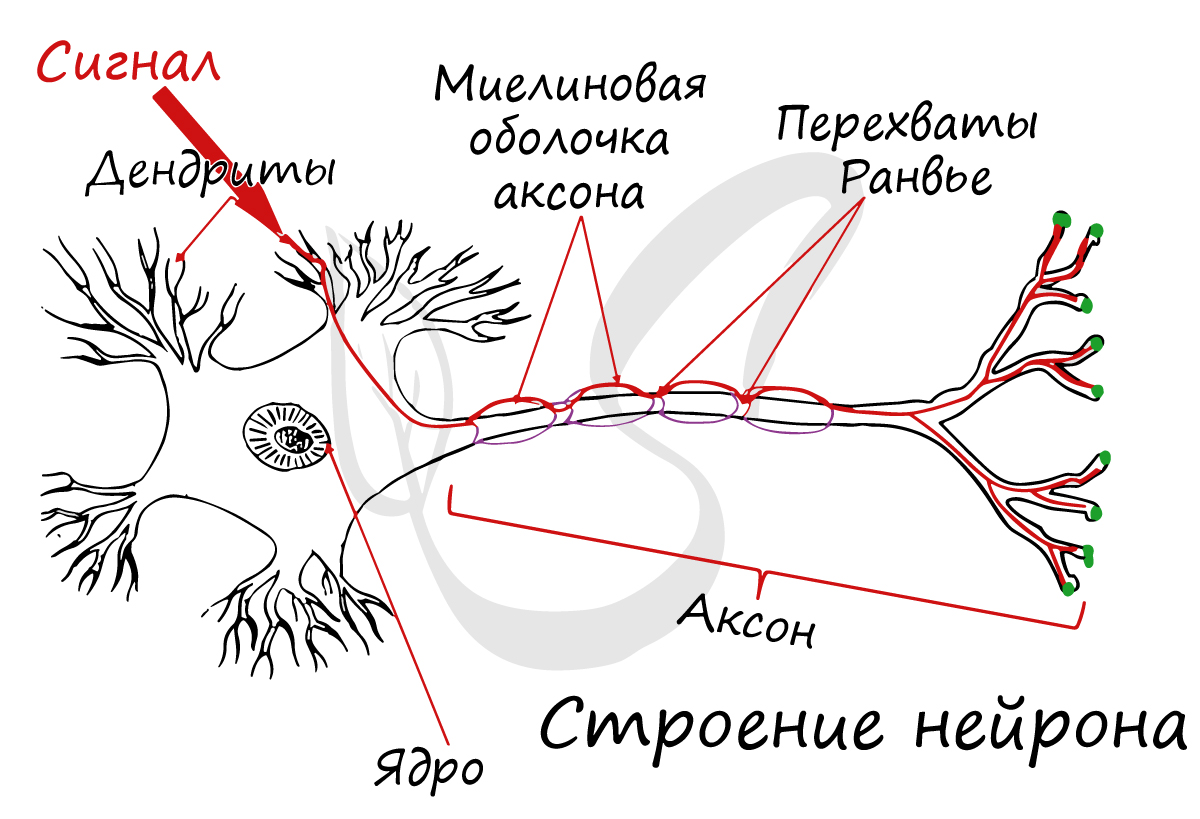
Классификация нейронов
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
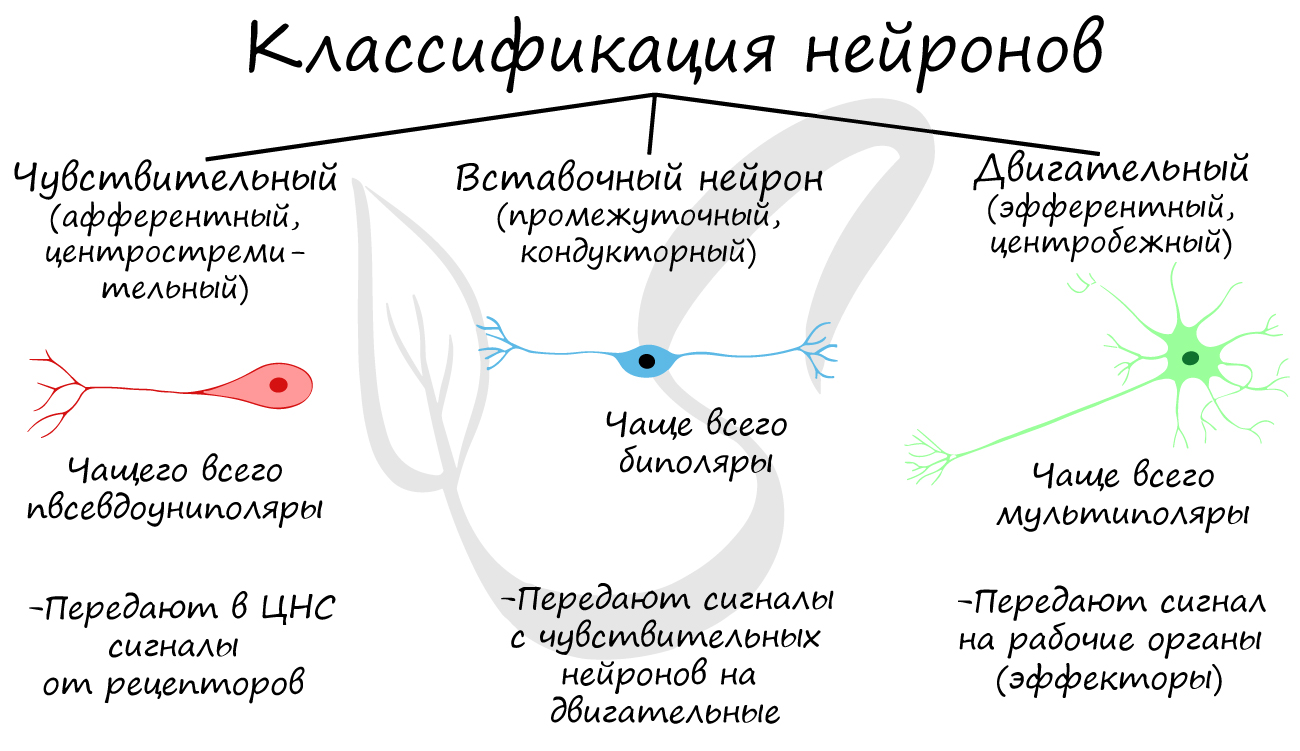
Чувствительные нейроны также называются афферентные, центростремительные, сенсорные, воспринимающие — они воспринимают раздражения, преобразуют их в нервные импульсы и передают в ЦНС. Рецептором называют концевое окончание чувствительных нервных
волокон, воспринимающих раздражитель.
Вставочные нейроны также называются промежуточные, ассоциативные — они обеспечивают связь между чувствительными и двигательными
нейронами, передают возбуждение в различные отделы ЦНС, участвуют в обработке информации и выработке команд.
Двигательные нейроны по-другому называются эфферентные, центробежные, мотонейроны — они передают нервный импульс (возбуждение) на
эффектор (рабочий орган). Наиболее простой пример взаимодействия нейронов — коленный рефлекс (однако вставочного нейрона
на данной схеме нет). Более подробно рефлекторные дуги и их виды мы изучим в разделе, посвященном нервной системе.
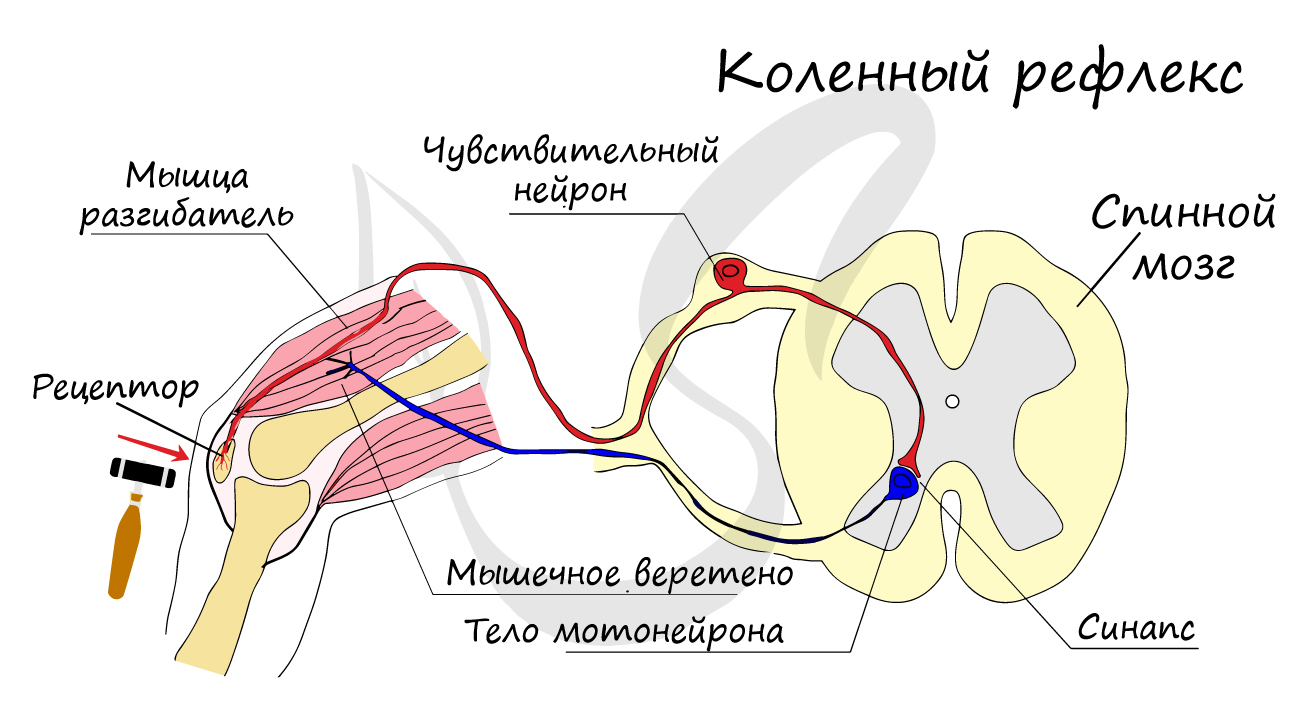
Синапс
На схеме выше вы наверняка заметили новый термин — синапс (греч. sýnapsis — соединение). Синапсом называют место контакта между двумя нейронами или между
нейроном и эффектором (органом-мишенью). В синапсе нервный импульс «преобразуется» в химический: происходит выброс особых
веществ — нейромедиаторов (наиболее известный — ацетилхолин) в синаптическую щель.
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с
нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают
сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
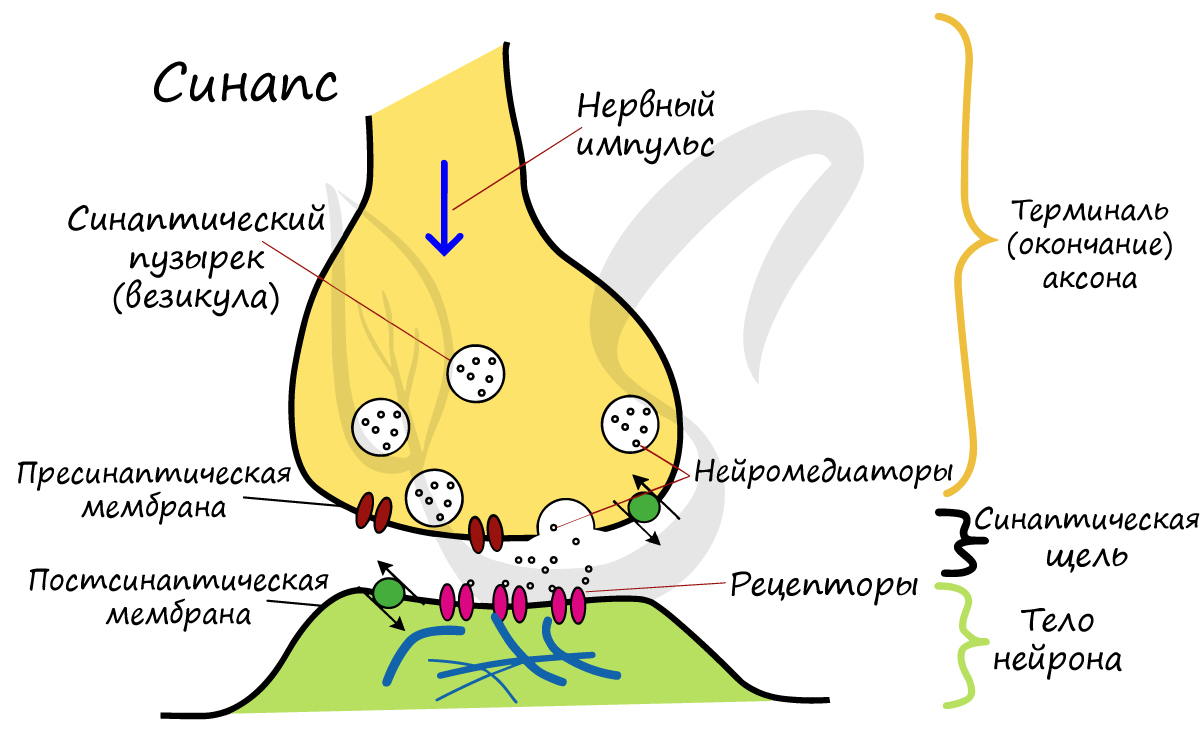
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение (нервный импульс)
передается другому нейрону. Так устроена нервная система: электрический путь передачи сменяется
химическим (в синапсе).
Яд кураре
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими Не могу утаить
историю о яде кураре, который используют индейцы для охоты с древних времен.
Этот яд блокирует ацетилхолиновые рецепторы на постсинаптической мембране, и, как следствие, химическая передача возбуждения с
одного нейрона на другой становится невозможна. Это приводит к тому, что нервные импульсы перестают поступать к эффекторам,
в том числе к дыхательным мышцам (межреберным, диафрагме), вследствие чего дыхание останавливается и наступает смерть животного.

Нервы и нервные узлы
Собираясь вместе, отростки нейронов (нервные волокна) образуют пучки нервных волокон. Нервные пучки объединяются в нервы, которые покрыты соединительнотканной оболочкой.
В случае, если тела нейронов концентрируются в одном месте за пределами центральной нервной системы, их скопления
называют нервным узлом — или ганглием (от др.-греч. γάγγλιον — узел).
В случае сложных соединений между нервными волокнами говорят о нервных сплетениях. Одно из наиболее известных —
плечевое сплетение.
Болезни нервной системы
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения
чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом
движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет
невозможно: возникает паралич, но чувствительность может сохраняться.
Существует тяжелое мышечное заболеванием — миастения (от др.-греч. μῦς — «мышца» и ἀσθένεια — «бессилие, слабость»), при
котором собственные антитела разрушают мотонейроны (двигательные нейроны).

Постепенно любые движения мышцами становятся для пациента все труднее,
становится тяжело долго говорить, повышается утомляемость. Наблюдается характерный симптом — опущение верхнего века.
Болезнь может привести к слабости диафрагмы и дыхательных мышц, вследствие чего дыхание становится невозможным.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.