Ответ на раздражение возбудимой клетки (в нашем случае – аксона) можно представить в виде трех последовательных фаз: формирование потенциала действия (проведение импульса), период абсолютной рефрактерности, период относительной рефрактерности.
После возникновения потенциала действия в аксоне наблюдается период рефрактерности, т.е. период, когда возбудимость клетки снижается с последующим ее восстановлением.
Абсолютная рефрактерность – период, когда клетка не в состоянии генерировать потенциал действия, несмотря на силу импульса.
Относительная рефрактерность – период, когда клетка в состоянии генерировать потенциал действия, однако сила стимулирующего импульса должна быть больше пороговой, т.к. восстановление возбудимости клетки произошло не полностью.
Максимальная частота проведения
Проводить импульс аксон не в состоянии только в период абсолютной рефрактерности, который, по условию задачи, длится 1 мс. В период относительной рефрактерности проведение импульса возможно, однако сила стимулирующего импульса должна быть больше предыдущего.
Тогда время одного цикла работы в максимальном режиме будет состоять из:
- времени проведения (1 мс);
- времени абсолютной рефрактерности (1 мс).
Итого:
1мс + 1мс = 2мс;
Чтобы найти частоту за 1 с, разделим общее время на время одного цикла:
1с/2мс = 1000с/2 мс = 500 импульсов/с – максимальная частота проведения.
Оптимальная частота проведения
Т.к. в предыдущем случае полное восстановление клетки не происходило, то, работая в таком режиме, аксон очень быстро истощится. Чтобы этого не произошло, необходимо проводить импульсы с оптимальной частотой, т.е. учитывать время относительной рефрактерности.
Тогда время одного цикла работы в оптимальном режиме будет состоять из:
- времени проведения (1 мс);
- времени абсолютной рефрактерности (1 мс);
- времени относительной рефрактерности (2 мс).
Итого:
1мс + 1мс + 2мс = 4мс;
Чтобы найти частоту за 1 с, разделим общее время на время одного цикла:
1с/4мс = 1000с/4мс = 250 импульсов/с – оптимальная частота проведения.
Нейрон-детектор частоты следования импульсов. Нейрон такого типа содержит дендриты (узел ветвления), реализующие операцию выделения определенного интервала (частоты) следования импульсов (гл. II, 4). [c.57]
При длительном воздействии сильного раздражителя больщинство рецепторов вначале возбуждают в сенсорном нейроне импульсы с большой частотой, но постепенно частота их снижается, и это ослабление ответа во времени называют адаптацией. Например, войдя в комна- [c.318]
Во-первых, она повышает чувствительность рецептора, обеспечивая немедленный ответ на стимуляцию, которая в противном случае была бы слишком мала, чтобы вызвать реакцию сенсорного нейрона любое, даже очень незначительное, изменение интенсивности сигнала будет вызывать изменение частоты импульсов в этом нейроне. [c.319]
Первая стадия памяти связана с получением информации, которая передается в центр. Имеется основание предполагать, что в восприятии всех сенсорных раздражений лежит один и тот же механизм. Этот вывод исходит из того, что афферентные импульсы идут от рецепторной клетки в ЦНС одним и тем ке способом, путем биотоков возбуждения. Одна афферентная импульсация в зависимости от вида раздражения отличается от другой в основном по частоте и амплитуде. Отсюда можно заключить, что информация в ЦНС воспринимается одним и тем же кодом. Другой особенностью восприятия сенсорных раздражений является то, что один и тот н е импульс воспринимается разными нейронами по-разному. Причину этого явления нужно искать в различии обменных процессов в разных нейронах. [c.7]
Благодаря временной и пространственной суммации мембранный потенциал тела одного постсинаптического нейрона регулируется частотой разрядов множества пресинаптических нейронов. В результате интеграции всех входных сигналов постсинаптическая клетка формирует определенный ответ, обычно в виде импульсов для передачи сигналов другим клеткам, нередко находящимся в отдаленных частях организма. Этот ответный сигнал отражает величину суммарного ПСП в теле клетки. Однако, хотя суммарный ПСП все время плавно изменяется, потенциалы действия имеют постоянную амплитуду и подчиняются закону всё или ничего . Единственной переменной величиной при передаче сигна- [c.321]
Из электрофизиологических экспериментов следует, что зависимость средней частоты генерации нейроном импульсов от суммы токов, деполяризующих сому, имеет участок, близкий к линейному [27, 28]. Это дает основание рассматривать сому нейрона как сумматор возбуждений приходящих по соматическим синапсам. [c.20]
Активация одного командного нейрона вызывает определенный двигательный акт. Временные характеристики импульсации обычно не играют решающей роли в осуществлении этого эффекта. В то же время частота импульсов может иметь существенное значение ее повышение может изменить характер конечного эффекта. [c.92]
Еще одно общее свойство нейронов, имеющее важнейшее значение для работы мозга, состоит в том, что частота импульсов в нейроне зависит от силы и длительности стимула. Чем сильнее стимул, тем быстрее залп Спайков (пиков), проходящих по аксону. Таким образом, функция мозга человека в значительной степени сводится к расшифровке потока импульсов. Частота импульсов, проходящих по нейронам, колеблется от нескольких импульсов в секунду до максимальной частоты для большинства нервных волокон — 200 имп./с (в клетках Реншоу в спинном мозге частота импульсации может достигать 1600 импульсов в секунду). Максимум частоты импульсации определяется величиной рефрактерного периода, составляющего мс (гл. 5, разд. Б, 3). [c.327]
До недавнего времени полагали, что единственная функция нервного импульса состоит в быстром проведении сигналов по аксонам на большие расстояния. Важная роль этой функции состоит в том, что сила раздражения кодируется в нейронах путем изменения частоты импульсации. Информация, закодированная в нервных импульсах, передается на другие нейроны через синапсы, образуемые нервными окончаниями. Ни в коей мере не умаляя важности этой функции потенциала действия, мы хотели бы подчеркнуть, что возбудимость может влиять и на другие . процессы жизнедеятельности нервных клеток. [c.172]
Когда нервный импульс приходит к синаптическому окончанию, происходит освобождение медиатора, который частично взаимодействует с рецептором постсинаптической мембраны. Остальная его часть разрушается специальным ферментом или захватывается обратно в пресинаптическую терминаль. Следствием реакции медиатора с постсинаптическим аппаратом является изменение ионных потоков, протекающих через поверхность клетки. В результате происходит сдвиг мембранного потенциала и повышение концентрации ионов калия вне клетки и ионов кальция внутри нее. Сами по себе процессы, вызванные одним импульсом, чрезвычайно кратковременны (не более О, I с), но если импульсы поступают регулярно и с достаточно высокой частотой, возникает процесс суммации, при котором определенные сдвиги в концентрации ионов могут сохраняться достаточно долго. В частности, при прохождении залпа импульсов вьщеляющиеся ионы калия могут в значительных количествах диффундировать к окружающим нейрон клеткам глии и влиять на их деятельность, что в некоторых теориях рассматривается как один из факторов, участвующих в процессах памяти. [c.381]
При слабых сигналах деполяризации будут достигаться не сразу импульсы, возникающие у основания аксона, будут отделены друг от друга более продолжительными интервалами, т. е. слабые раздражения возбудят импульсы низкой частоты, а сильные — импульсы высокой частоты. В организмах любые раздражения передаются по путям, состоящим из множества нейронов, что делает связь более надежной и открывает возможности регулирования работы органов при варьировании силы возбуждающих сигналов. Известны типы нейронов, которые ослабляют сигналы, проходящие от других нейронов (тормозные нейроны), и данный нейрон фактически получает и возбуждающие и тормозные сигналы. Нейрон фактиви-руется, если сумма тех и других превосходит по величине порог его возбуждения. До сих пор речь шла о вставочных нейронах, которые играют роль передатчиков нервного возбуждения. Очень интересны и нейроны, находящиеся на воспринимающих концах нервной цепи, — рецепторные и эффекторные или двигательные. Рецепторы принимают раздражения различных типов это может быть химическое раздражение, механическое, например, давление, прикосновение, температурное, электрическое и др. Все виды раздражений передаются центральной нервной системе в виде электрических импульсов — классический пример биологического кодирования, четко показывающий сходство между функциями вычислительных машин, и теми функциями, которые сами собой возникли в итоге длительной эволюции динамических систем. [c.229]
В полипептидной цепи эта группа, как предполагалось в модели Лаки и Коулсона, отцает четыре электрона для образования общей я-орбитали. Согласно этой модели белок является полупроводником, причем л-электронные орбитали располагаются перпендикулярно оси полипептидной цепи. Позже Эванс и Герей, рассматривая пептидную группу как элементарную ячейку, пришли к выводу о наличии в молекуле белка трех энергетических зон, из которых одна свободна. Более точные расчеты показали, что ширина запрещенной зоны в белках довольно велика и равна 5 эВ. Бриллюэн предложил модель, в которой зоны проводимости белка получаются за счет перекрытия ст-связей. В этой модели ширина запрещенных зон еще больше (8—10 эВ). Проблема полупроводи-мости белковых систем пока ждет решения. Эксперимент показывает, что энергия фотовозбуждения отдельных групп, связанных с белковой цепью, может мигрировать на значительные расстояния и вызывать флуоресценцию других групп. Комплекс миоглобина с оксидом углерода (II) отщепляет СО при действии излучения, которое не поглощается гемином (т. е. группой, непосредственно связанной с СО), но поглощается триптофаном и тирозином — аминокислотами, остатки которых входят в состав белка миоглобина. Здесь энергия мигрирует от белка к геминовой группе. Эти важные свойства белков показывают, что белки в некоторых случаях способны передавать энергию возбуждения, т. е., в общем случае, сигналы . В ходе эволюции функции передачи сигналов в форме серии дискретных импульсов, частота которых зависит от силы раздражения, перешли к более совершенной системе — нейронам нервной сети. [c.348]
| Рис. 18-37. Перекодирование суммарного ПСП в частоту импульсного разряда на выходе нейрона (в аксонеХ Из графиков А и Б видно, как частота импульсов в аксоне возрастает при увеличении суммарного ПСП ва графике В представлена общая зависимость. Г-способ измерения суммарного ПСП.. 4 и на верхних графиках-суммарная интенсив-иость синаптической стимуляции, воспринимаемой телом клетки на нижних графиках-соответствующие разряды импульсов, посылаемых по аксону. На верхних графиках показано, как выглядели бы ПСП, если бы импульсный разряд каким-то образом блокировался. |  |
Однако непрерывности разряда для функции нейрона недостаточно-нужно еще, чтобы частота этого разряда отражала интенсивность стимуляции. Детальные расчеты показывают, что простая система натриевых н медленных калиевых каналов не отвечает поставленным требованиям. Если сила непрерывной стимуляции клеткн ннже определенного порогового уровня, по-теациал действия не возникнет совсем если же она превысит порог, то сразу начнется частая импульсация. Проблему решают быстрые калиевые каналы. В открытом состоянии эти каналы препятствуют действию деполяризующих стимулов и тормозят возникновение импульсов, а открывание этих каналов регулируется таким образом, что онн снижают частоту разряда прн уровнях стимуляции, которые лишь немного выше порогового уровня. Таким путем быстрые калиевые каналы способствуют более плавной зависимости между нитеисивиостью стимуляции и частотой разряда. [c.109]
Почти в точности на тех же принципах основано преобразование сигналов в органах чувств. Это можно хорошо проиллюстрировать иа примере мышечных рецепторов растяжения, где первоначальный стимул, вызывающий изменение проницаемости мембраны, имеет механическую, а не химическую природу. Рецепторы растяжения доставляют нервной системе информацию о длине мышцы и скорости ее изменения. Эта сенсорная обратная связь (наряду с сигналами от головного мозга и некоторых частей спинного мозга) помогает регулировать импульсацию двигательных нейронов, как это объяснено в подписи к рис. 18-45. Каждая мышца содержит группы видоизмененных мышечных волокон, образующих так называемые мышечные веретена. Каждое отдельное волокно в веретене обвито окончаниями сенсорных нейронов (рис. 18-45). При растяжении волокон веретена в этих нейронах возникают импульсы (потенциалы действия), которые передаются в спинной мозг. Электрическое поведение одного сенсорного нейрона можно исследовать с помошью внутриклеточного электрода, помещенного около того места, где нейрон прилегает к волокну. Частота импульсного разряда градуально [c.119]
Специальные преобразователи переводят сенсорные стимулы в форму нервных сигналов. Например, в рецепторе растяжения мышцы окончание сенсорного нерва деполяризуется при растяжении и величина деполяризации-рецепторный потенциал-для дальнейшей передачи перекодируется в частоту импульсного разряда. Слуховые волосковые клетки, избирательно реагирующие на звуки определенной частоты, сами не посылают импульсов, а передают сигналы о величине рецетпорного потенциала соседним нейронам через химические синапсы. Таким же образом действуют фоторецепторы глаза. В фоторецепторах свет вызывает конформационное изменение молекул родопсина, и это благодаря участию внутриклеточного второго посредника ведет к закрытию натриевых каналов в плазматической мембране, к ее гиперполяризации и в результате-к уменьшению количества высвобождаемого медиатора. Далее вставочные нейроны передают сигнал ганглиозным клеткам сетчатки, которые пересылают его в мозг в виде потенциалов действия. Проходя череъ нейронную сеть с конвергентными, дивергентными и тормозными латеральными связями, информация подвергается обработке, благодаря которой клетки высших уровней зрительной системы могут выявлять более сложные особенности пространственного распределения световых стимулов. [c.130]
Получается, что нейрон при передаче сигналов <обед-няет информацию. Правда, оказывается, что хотя нейрон ничего не может сообш ить об источниках входных сигналов, он может сообщить об их силе. Как это делается Амплитуда входного сигнала — зто сумма (алгебраическая, конечно) всех синаптических потенциалов, создаваемых активными синапсами. Пусть во время действия сигнала амплитуда его не меняется (это значит, что постоянна разность между числом активных возбуждающих и числом тормозных синапсов). Тогда через мембрану нейрона все время будет течь примерно постоянный синаптический ток. Легко сообразить, что чем больше ток, идущий через мембрану, тем с большей частотой будет работать нейрон (рис. 55). Действительно, после каждого очередного импульса мембрана нейрона гиперполяризуется, так как открываются калиевые каналы. Чем больше ток, текущий. через мембрану нейрона тем быстрее он возвратит его [c.215]
Из приведенных экспериментальных данных следует, что информация о частоте заключена не только в пространственной кар-типе распределения возбуждения нейронов, но также и во временной организации последовательпости импульсов у каждого нейрона. Можно ожидать, что носледпее обстоятельство используется на последующих уровнях слуховой системы для более точного [c.79]
В нервной системе обнаруживают нейроны, реакция кс1торых на входные сигналы проявляется в градуальном изменении пара-метров выходного сигнала. Выходной сигнал может быть закодирован частотой генерации импульсов (спайков) или длиной п чки [c.43]
Результаты анализа привели к выводу, что среди нервных клеток существует тип нейронов-детекторов, которые срабатывают только при приходе на и входы ансамбля импульсов с определенными временными интервалами друг относительно друга. Или вероятность срабатывания тем выше, чем ближе ансамбль импульсов к тому, па который настроен данный нейрон. Нейроны-детекторы выполняют такие функции, как детекция момента совпадения двух сигналов, детекция определепной задержки между двумя импульсами, детекция скорости и направления перемещения возбуждения по контролируемой нейроном совокупности нервных элементов (других нейронов, рецепторов), детекция кодовой последовательности (т. е. определенным образом расставленной во времени последовательности импульсов по одному аксону), детекция определенной частоты следования импульсов. [c.62]
Наряду с этим известны физиологические данные о ие дискретной, а градуальной реакции нервных клеток, когда параметры импульсной последовательности в их ответе (число импульсов, частота их следования) зависят от параметров стимула. Проведенный анализ операций в дендритах также указывает на возможность ряда градуальных преобразований управляемая задержка (латентность) появления сигнала на выходе нейрона, управляемая трансформация импульсной носледовательности. Совокупность этих сведений позволяет заключить, что сущесгвует другой тип нервных клеток, ото нейроны-преобразователи, производящее аналоговые преобразования входных сигналов. Нейроны-преобразо- [c.62]
Для нейронов-преобразователей к числу основных операций следует отнести операцию накопления возбуждения или временную суммацию импульсов , широко отмечаемую в работах по физиологии нервной клетки. Как известно, такого рода приемники воспринимают информацию, закодированную в виде числа (за определенное время) или частоты следования импульсов. Выходной сигнал нейрона-нреобразователя, с пашей точки зрения, может быть двух видов. В одних случаях это люжет быть трансформируемая частота следования импульсов (результат суммирования или умножения на постоянный или управляемый коэффициент) тогда, когда информация предназначается другим нейронам-преобразователям. В других случаях выходной сигнал может быть импульсным, временное положение которого изменяется (например, есть функция числа суммируемых входных импульсов), но это указывает на то, что информация адресована нейро-нам-детектора г. [c.63]
Относительно третьего различия известно следующее. В отсутствие стимула все нейроны спирального ганглия проявляют спонтанную активность, частота которой лежит в диапазоне от единиц до сотпи импульсов в секунду [93]. Распределение частот спонтанной активности (рис. 33) имеет два максимума — один в области 5 Гц, второй — в области 30—80 Гц. Следовательно, есть основания считать, что по величине частоты спонтанной активности нейроны спирального ганглия делятся на две группы — низкочастотную (Т сп 5 Гц) и высокочастотную (Т оп 50 Гц). [c.81]
Электрофизиологические данные говорят о том, что нейроны упорядоченно расположены по оптимальным частотам тонотопика),что они являются физическими, содержащими в своем ответе на стимул — звуковой щелчок — 1 — 2 импульса. Вероятность появления импульса зависит от разности моментов At и относительной разности амплитуд АА/А звуковых сигналов слева и справа [183, 184]. [c.111]
Нервные пути, регулирующие деятельность сердца, изображены на рис. 19.9. Симпатическая иннервация сердца осуществляется постганглионарными волокнами, идущими от симпатической системы. При возбуждении из окончаний этих волокон освобождается норадреналин, действующий на Ргздренорецеп-торы клеток сердца. Стимуляция этих рецепторов приводит к активации аденилатциклазы в дальнейшем развертывается цепь событий, которую мы уже рассматривали как в настоящей главе, так и при обсуждении рис. 9.9. В конечном счете увеличивается проницаемость мембраны мышечных клеток главным образом для ионов Са= + это приводит к повышению частоты и силы сердечных сокращений и к ускорению проведения импульсов. Увеличение кальциевой проницаемости сходно с тем, что было обнаружено в некоторых нейронах (см. гл. 9) при модулирующем влиянии медиаторов на возбудимые кальциевые каналы. [c.43]
В лаборатории Циммермана проводились исследования зависимости амплитуд определенных компонент ВМП и ВЭП от интенсивности и характера звукового стимула [280]. Выявлено, что амплитуда ВМП в ответ на сигналы с одинаковым средним значением звукового давления максимальна для чистых тонов, ниже для белого (содержащего все тона) шума и минимальна для щелчков. Это согласуется с результатами внутрикорковых измерений активности нейронных колонок. Поэтому в дальнейшем, как и в рдае других лабораторий [239, 269], в качестве стимула использовались кратковременные импульсы звучания с частотой 1 кГц [283]. Изучался отклик возбуждаемого этими импульсами конкретного источника в слуховой области коры, магнитное поле которого соответствовало полю токового диполя. Поэтому сигнал принимался с помощью недиагонального (восьмеркообразного) градиометра с расстоянием между центрами петель (базой) 4 см. Этот прибор наилучшим образом приспособлен к регистрации магнитного поля токового диполя, когда одна из петель восьмерки находится над одним, а другая — над другим экстремумом поля (см. рис. 21). К недостатку такого градиометра можно отнести то, что он оптимален для приема сигнала только с определенной глубины, так как база его фиксирована. [c.159]
Молекулярная биология клетки — Том 3 — Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Нервная система
Роль ионных каналов в совместной переработке информации нейронами
В центральной нервной системе нейроны обычно принимают сигналы от множества пресинаптических клеток — их число может достигать тысячи или даже нескольких тысяч. Например, на типичном мотонейроне спинного мозга синапсы образуют тысячи нервных окончаний от сотен и, возможно, тысяч различных нейронов; тело нейрона и дендриты почти полностью покрыты синапсами (рис. 19-30). Некоторые из этих синапсов передают сигналы от головного мозга, другие доставляют сенсорную информацию от мышц и кожи, третьи сообщают результаты «вычислений», производимых вставочными нейронами спинного мозга. Мотонейрон должен интегрировать информацию, получаемую из этих многочисленных источников, и либо реагировать, посылая сигналы по аксону, либо оставаться в покое.
Мотонейрон служит типичным примером того, как отдельные нейроны участвуют в фундаментальной задаче вычисления правильного выходного сигнала в ответ на сложную совокупность входных сигналов. Из множества синапсов на мотонейроне одни будут стремиться возбудить его, а другие — затормозить. Хотя все окончания аксона данного мотонейрона выделяют один и тот же медиатор, мотонейрон имеет много различных рецепторных белков, сосредоточенных на разных постсинаптических участках его поверхности. В каждом из таких участков под действием пресинаптических импульсов открывается или закрывается определенная группа каналов, в результате чего в мотонейроне происходит характерное изменение потенциала — возникает постсинаптический потенциал (ПСП). Деполяризация соответствует возбудительному ПСП (возникающему, например, при открытии каналов для ионов натрия), а гиперполяризация — тормозному ПСП (появляющемуся, например, при открытии хлоридных каналов). ПСП, генерируемые разными синапсами на одном и том же нейроне, очень сильно варьируют по величине и по продолжительности. В одном синапсе на мотонейроне пришедший импульс может вызвать деполяризацию менее 0,1 мВ, а в другом — деполяризацию в 5 мВ. Но, как мы увидим, природа системы такова, что даже небольшие ПСП, суммируясь, могут давать большой эффект.

Рис. 19-30. Тело спинномозгового мотонейрона. Показана лишь небольшая часть из многих тысяч нервных окончаний, образующих на клетке синапсы и приносящих сигналы от других частей организма для регулирования ее импульсного разряда. Участки плазматической мембраны мотонейрона, не занятые синаптическими окончаниями, покрыты глиальными клетками (на рисунке не показаны).
19.4.1. Сдвиг мембранного потенциала в теле постсинаптической клетки — это результат пространственной и временной суммации множества постсинаптических потенциалов [25, 26]
Хотя мембрана дендритов и тела большинства нейронов богата рецепторными белками, она содержит очень мало потенциал-зависимых натриевых каналов и поэтому относительно невозбудима. Одиночные ПСП, как правило, не приводят к возникновению потенциала действия. Каждый пришедший сигнал точно отображается величиной градуального ПСП, которая уменьшается по мере удаления от входного синапса. Если сигналы одновременно приходят к синапсам, находящимся на одном и том же участке дендритного дерева, то общий ПСП будет близок к сумме индивидуальных ПСП, причем тормозные ПСП будут учитываться с отрицательным знаком. В то же время суммарное электрическое возмущение, возникшее в одном постсинаптическом участке, будет распространяться на другие участки благодаря пассивным кабельным свойствам мембраны дендрита.
Тело нейрона, где сходятся все эффекты от ПСП, обычно невелико (менее 100 мкм в диаметре) по сравнению с дендритным деревом, длина ветвей которого может измеряться миллиметрами. Поэтому мембранный потенциал тела клетки и ближайших к нему частей отростков будет примерно одинаков — это будет совокупный результат эффектов, производимых всеми входными сигналами с учетом удаленности того или иного синапса от тела нейрона. Таким образом, можно сказать, что суммарный постсинаптический потенциал тела клетки — это результат пространственной суммации всех полученных стимулов. Если преобладают возбуждающие входные сигналы, то тело клетки деполяризуется, если тормозные — обычно гиперполяризуется.
В то время как пространственная суммация интегрирует эффекты сигналов, принятых разными участками мембраны, временная суммация объединяет сигналы, поступившие в разное время. Нейромедиатор, высвобождающийся после прибытия потенциала действия к синапсу, создает на постсинаптической мембране ПСП, который быстро достигает пика (благодаря кратковременному открытию лиганд-зависимых ионных каналов) и затем экспоненциально (что определяется емкостью мембраны) снижается до исходного уровня. Если второй импульс придет до того, как первый ПСП полностью затухнет, то этот второй ПСП суммируется с оставшимся «хвостом» первого. Когда после некоторого периода покоя приходит длинный залп быстро повторяющихся импульсов, каждый последующий ПСП будет накладываться на предыдущий, давая в итоге большой ПСП, величина которого отражает частоту разряда пресинаптического нейрона (рис. 19-31). Таким образом, суть временной суммации состоит в том, что частота получаемых сигналов преобразуется в величину суммарного ПСП.

Рис. 19-31. Временная суммация. Перекрывающиеся черные кривые накрашенной области показывают индивидуальный вклад каждого из последовательных пресинаптических импульсов в суммарный постсинаптический потенциал.
19.4.2. Для передачи информации на большие расстояния суммарный ПСП снова преобразуется в частоту нервных импульсов [27]
Благодаря временной и пространственной суммации мембранный потенциал тела одного постсинаптического нейрона регулируется частотой разрядов множества пресинаптических нейронов. В результате интеграции всех входных сигналов постсинаптическая клетка формирует определенный ответ, обычно в виде импульсов для передачи сигналов другим клеткам, нередко находящимся в отдаленных частях организма. Этот ответный сигнал отражает величину суммарного ПСП в теле клетки. Однако, хотя суммарный ПСП все время плавно изменяется, потенциалы действия имеют постоянную амплитуду и подчиняются закону «всё или ничего». Единственной переменной величиной при передаче сигналов с помощью импульсов остается временной интервал между последовательными импульсами. Поэтому для передачи информации на большие расстояния величина суммарного ПСП должна быть преобразована, или перекодирована, в частоту импульсного разряда (рис. 19-32) Такое кодирование достигается с помощью специальной группы потен циал-зависимых ионных каналов, сосредоточенных у основания аксона в области, называемой аксонным холмиком (см. рис. 19-30).
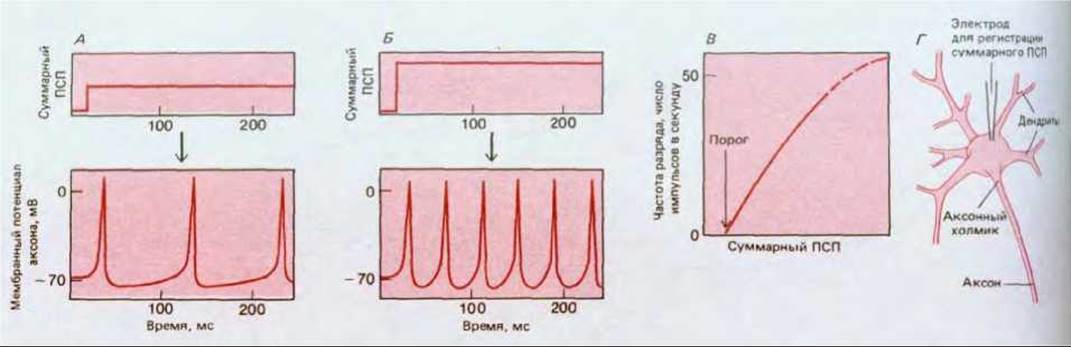
Рис. 19-32. Перекодирование суммарного ПСП в частоту импульсного разряда в аксоне. Из графиков А и Б видно, как частота импульсов в аксоне возрастает с увеличением суммарного ПСП; на графике В представлена общая зависимость. Г-способ измерения суммарного ПСП. А и Б: на верхних графиках — суммарная интенсивность синаптических стимулов, получаемых телом клетки; на нижних графиках соответствующие разряды, посылаемые по аксону. На верхних графиках показано, как выглядели бы ПСП, если бы импульсный разряд был каким-то образом блокирован.
Прежде чем объяснить, как действуют такие каналы, необходимо сделать некоторые уточнения. Само по себе возникновение импульса приводит к резким изменениям мембранного потенциала всего тела клетки, который уже не будет прямо отражать суммарную синаптическую стимуляцию, получаемую клеткой. Поэтому очень трудно провести точный анализ кодирующего механизма. В последующем чисто качественном описании мы будем употреблять выражения «сила синаптической стимуляции» или «суммарный ПСП, имея в виду тот суммарный ПСП, который создавался бы при условии, что генерирование импульсов каким-то образом подавляется; и мы будем предполагать что именно этот основной суммарный ПСП и является причиной возникновения нервных импульсов.
19.4.3. Кодирование требует совместного действия различных ионных каналов [28]
Проведение нервных импульсов зависит главным образом, а во многих аксонах позвоночных почти полностью, от потенциал-зависимых натриевых каналов. Первоначально импульсы генерируются мембраной аксонного холмика, где таких каналов очень много. Но для осуществления особой функции кодирования мембрана аксонного холмика должна содержать еще по меньшей мере четыре класса ионных каналов — три избирательно проницаемых для ионов калия и один проницаемый для Са2+. Три разновидности калиевых каналов обладают различными свойствами — мы будем называть их медленными, быстрыми и Са2+ — зависимыми калиевыми каналами. Кодирующие функции этих канала наиболее изучены на гигантских нейронах моллюсков, но те же принципы используются, по-видимому, и в других нейронах.
Чтобы понять, для чего нужны каналы нескольких типов, посмотрим, как будет вести себя мембрана нервной клетки, содержащая только один вид потенциал-зависимых каналов — натриевые каналы. При слабой синаптической стимуляции, не доводящей деполяризацию мембраны аксонного холмика до порогового уровня, потенциал действия не буди возникать. При постепенном усилении стимуляции порог будет достигнут, натриевые каналы откроются и возникнет потенциал действия.
В результате последующей инактивации натриевых каналов появившийся потенциал действия исчезнет. Прежде чем сможет возникнуть другой потенциал действия, натриевые каналы должны будут выйти из инактивированного состояния. Но для этого необходимо, чтобы величина мембранного потенциала вновь достигла большого отрицательного значения, а этого не произойдет, пока поддерживается сильный деполяризующий стимул (от ПСП). Поэтому для того, чтобы реполяризовать мембрану после импульса и подготовить клетку для проведения следующего импульса, нужен еще один вид каналов. Эту функцию выполняют медленные калиевые каналы, которые уже упоминались, когда речь шла о распространении потенциала действия (разд. 19.2.2). Эти каналы зависимы от потенциала и открываются при деполяризации мембраны так же, как и натриевые, но происходит это с некоторым запаздыванием. Открываясь во время спада потенциала действия, калиевые каналы пропускают ионы К+ из клетки наружу, в результате чего устраняется эффект даже длительного деполяризующего стимула и мембранный потенциал возвращается к уровню равновесного калиевого потенциала. Этот потенциал настолько отрицателен, что натриевые каналы выводятся из состояния инактивации. Кроме того, прекращается и выход калия из клетки: реполяризация мембраны приводит к тому, что медленные калиевые каналы опять закрываются (так и не успев инактивироваться). Как только произошла реполяризация, деполяризующие стимулы от синаптических входов могут вновь поднять мембранный потенциал до порогового уровня и вызвать очередной потенциал действия. Благодаря этому непрерывная стимуляция дендритов и тела клетки приводит к многократно повторяющемуся возбуждению аксона.
Однако недостаточно только непрерывности разряда — нужно еще, чтобы его частота отражала интенсивность стимуляции. Детальные расчеты показывают, что простая система натриевых и медленных калиевых каналов не отвечает этому требованию. Если сила постоянного стимула ниже определенного порогового уровня, потенциалов действия не будет вовсе; если же сила стимуляции превысит порог, то сразу же начнется частая импульсация. Проблему решают быстрые калиевые каналы (называемые также А-каналами). Эти каналы тоже потенциалзависимы и открываются при деполяризации мембраны, но специфическая зависимость их от потенциала и кинетика инактивации такова, что они снижают частоту разряда при уровнях стимуляции, которые лишь ненамного выше порога. Таким образом, быстрые калиевые каналы помогают устранить разрыв непрерывности в соотношении между интенсивностью стимула и частотой разряда; в результате частота импульсов пропорциональна силе деполяризующего стимула в очень широком диапазоне (см. рис. 19-32).
19.4.4. Адаптация уменьшает реакцию на постоянный стимул [29]
Обычно процесс кодирования видоизменяют еще два типа каналов, имеющихся в аксонном холмике. О них уже упоминалось — это потенциал-зависимые кальциевые каналы и Са2+-зависимые калиевые каналы. Первые подобны кальциевым каналам, участвующим в высвобождении медиатора из окончаний аксона: в области аксонного холмика эти каналы открываются при возникновении потенциала действия и пропускают Са2+ внутрь аксона. Са2+-зависимые калиевые каналы отличаются от всех других каналов, описанных ранее. Они открываются при повышении концентрации кальция у внутренней поверхности мембраны нервной клетки.
Предположим, что сильный и продолжительный деполяризующий стимул приводит к длительной импульсации. В результате каждого импульса в клетку через потенциал-зависимые кальциевые каналы переходит небольшое количество ионов Са2+, так что их внутриклеточная концентрация постепенно поднимается до высокого уровня. Это ведет к открытию Са2+-зависимых калиевых каналов, и проницаемость мембраны для калия повышается, что затрудняет деполяризацию и увеличивает интервалы между последовательными импульсами. Таким образом, при длительном воздействии постоянного стимула сила ответа нейрона постепенно снижается. Это явление, в основе которого могут лежать и другие механизмы, называют адаптацией (рис. 19-33). Благодаря адаптации нейрон, так же как и нервная система в целом, способен с высокой чувствительностью реагировать на изменение стимула, даже если оно происходит на фоне сильной постоянной стимуляции (разд. 19.6.8). Это одно из общих приспособлений, благодаря которым мы, например, не замечаем постоянного давления одежды на наше тело, но в то же время быстро реагируем на внезапное прикосновение.

Рис. 19-33. Адаптация. При длительной стимуляции постоянной силы реакция клетки на стимул постепенно ослабевает, что выражается в уменьшении частоты импульсного разряда.
19.4.5. Сигналы могут передаваться не только по аксонам [30]
В типичном нейроне, о котором говорилось выше, дендриты и аксон резко различаются между собой по строению и функции. Однако некоторые нейроны не соответствуют такой схеме, хотя молекулярные основы их функционирования те же. Например, у большинства беспозвоночных нейроны чаще всего имеют униполярную организацию: тело клетки связано одним-единственным «стебельком» с системой ветвящихся клеточных отростков, среди которых не всегда можно отличить дендриты от аксона (рис. 19-34). Функциональные отличия тоже мот быть «смазаны», что встречается и у позвоночных, и у беспозвоночных: отростки, которые по их строению можно отнести к дендритам, часто образуют как пре-, так и постсинаптические структуры и способны как принимать сигналы, так и передавать их другим клеткам. И наоборот, входные синаптические сигналы иногда воспринимаются «стратегическими» участками аксона — например, вблизи окончания, где эти сигналы могут усиливать или тормозить высвобождение нейромедиатора из данного окончания, не влияя на передачу в окончаниях других ветвей того же аксона (рис. 19-35). Пример этого важного механизма пресинаптического торможения и пресинаптического облегчения будет рассмотрен позже (разд. 19.5.4).

Рис. 19-34. Нейроны мухи. Строе-кие их типично для большинства нейронов беспозвоночных животных; тело клетки соединено с системой отростков при помощи стебелька, так что нет дендритов, отходящих прямо от тела. Сходной организацией обладают и сенсорные нейроны из спинномозговых ганглиев позвоночных. (N. Strausfield, Atlas of an Insect Brain, New York, Springer, 1976.)
Синапсы, через которые дендриты передают стимулы другой клетке, играют важную роль в коммуникации между нейронами, расположенными друг от друга на расстоянии нескольких миллиметров или еще ближе. На такие расстояния электрические сигналы могут передаваться по дендриту пассивно от постсинаптического участка, где они были восприняты, до пресинаптического участка, где они регулируют высвобождение медиатора. Встречаются даже нейроны, совсем не имеющие аксона, не проводящие потенциалов действия и передающие все сигналы через дендриты. Более того, если дендритное дерево велико, то отдельные части его могут более или менее независимо использоваться для связи и обработки информации. Диапазон возможностей некоторых нейронов расширяется еще больше благодаря наличию в мембране дендритов потенциал-зависимых каналов, что позволяет дендритам проводить потенциалы действия. Таким образом, даже отдельный нейрон способен функционировать как очень сложное вычислительное устройство.
Заключение
Дендриты и тело типичного нейрона принимают множество различных возбуждающих и тормозных синаптических сигналов, которые подвергаются пространственной и временной суммации и создают суммарный постсинаптический потенциал тела клетки. Для передачи сигналов на большие расстояния величина этого потенциала преобразуется в частоту импульсного разряда при помощи системы ионных каналов в мембране аксонного холмика. Механизму такого кодирования часто свойственна способность к адаптации, и тогда клетка слабо реагирует на постоянный стимул, но чувствительна ко всякому изменению его силы. Существует много вариантов описанной общей схемы; например, не все нейроны дают ответ в виде потенциалов действия, дендриты могут быть не только постсинаптическими, но и пресинаптическими, а аксон — не только пресинаптическим, но и постсинаптическим.

Рис. 19-35. Аксоаксонный синапс. Нейромедиатор, выделяемый окончанием аксона клетки В, воздействует на каналы в окончании аксона клетки А, изменяя тем самым число квантов нейромедиатора, поступающих на клетку В при возбуждении Б. Если возбуждение Б ослабляет силу стимуляции В клеткой А, то говорят, что Б осуществляет пресинаптическое торможение. Противоположный эффект называют пресинаптическим облегчением.
Содержание:
Описание

Значительную часть нейробиологии составляют разделы о том, как работают отдельные нейроны и как информация передается от клетки к клетке через синапсы. Должно быть очевидным, что без этих сведений мы окажемся в положении человека, желающего понять работу радиоприемника или телевизора, но ничего не знающего о резисторах, конденсаторах и транзисторах. За последние десятилетия благодаря изобретательности ряда нейрофизиологов, из которых наиболее известны Эндрю Хаксли, Алан Ходжкин, Бернард Катц, Джон Экклз и Стивен Куффлер, были хорошо изучены физико-химические механизмы проведения нервных импульсов и синаптической передачи. Однако столь же очевидно, что сведения такого рода сами по себе еще не могут привести к пониманию работы мозга, подобно тому как одни лишь сведения о резисторах, конденсаторах и транзисторах не позволят понять работу радиоприемника или телевизора, а знание химии чернил — прочитать пьесу Шекспира.
Эту статью я начинаю с суммирования части того, что нам известно о нервном проведении и синаптической передаче. Большой подмогой в верном понимании существа дела будет знание основ физической химии и электричества, но я думаю, что и без этого читатель сможет получить о предмете достаточное представление. В любом случае для того, чтобы следить за изложением в последующих главах, вам понадобится лишь элементарное понимание этих вопросов.
Задача нервной клетки состоит в том, чтобы принимать информацию от клеток, которые ее передают, суммировать, или интегрировать, эту информацию и доставлять интегрированную информацию другим клеткам. Информация обычно передается в форме кратковременных процессов, называемых нервными импульсами. Во всякой клетке каждый импульс бывает точно таким же, как и любой другой, т. е. импульс — это стереотипный процесс. В любой момент частота импульсов, посылаемых нейроном, определяется сигналами, только что полученными им от передающих клеток, и передает информацию клеткам, по отношению к которым этот нейрон является передающим. Частота импульсов варьирует от одного в каждые несколько секунд или еще ниже до максимума около тысячи в секунду.
Что происходит, когда информация передается от одной клетки к другой через синапс? В первой — пресинаптической — клетке около основания аксона возникает электрический сигнал, или импульс. Импульс перемещается по аксону к его окончаниям. Из каждого окончания в результате этого импульса в узкий (0,02 мкм) заполненный жидкостью промежуток, отделяющий одну клетку от другой, — синаптическую щель — высвобождается химическое вещество, которое диффундирует ко второй — постсинаптической — клетке. Оно влияет на мембрану этой второй клетки таким образом, что вероятность возникновения в ней импульсов либо уменьшается, либо возрастает. После этого краткого описания вернемся назад и рассмотрим весь процесс подробно.
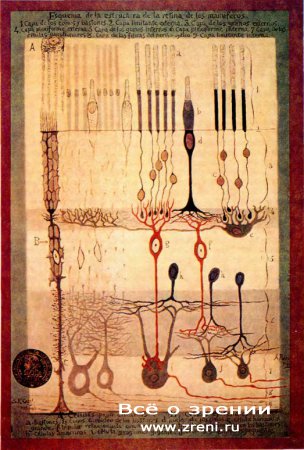
Рис. 8. Схема расположения нервных клеток на поперечом срезе сетчатки, нарисованная Сантьяго Рамон-и-Кахалом, величайшим нейроанатомом всех времен. От верхнего слоя, где показаны более тонкие палочки и более толстые колбочки, до нижнего, где направо выходят волокна зрительного нерва, толщина сетчатки составляет четверть миллиметра.
Нервная клетка омывается солевым раствором и содержит его внутри. В число солей входит не только хлористый натрий, но также хлористый калий, хлористый кальций и ряд других, менее обычных солей. Поскольку большинство молекул соли диссоциировано, жидкости как внутри, так и снаружи клетки содержат ионы хлора, калия, натрия и кальция (Cl-, К+, Na+ и Са2+).
В состоянии покоя электрические потенциалы внутри и снаружи клетки различаются примерно на одну десятую долю вольта, причем плюс находится снаружи. Точное значение ближе к величине 0,07 вольта, или 70 милливольт. Передаваемые нервами сигналы представляют собой быстрые изменения потенциала, перемещающиеся по волокну от тела клетки к окончаниям аксона. Я начну с описания того, как на клеточной мембране возникает разность потенциалов.

Рис. 9. На этой электронной микрофотографии (срез коры мозжечка крысы) синапс выглядит как узкая темная полоска в нижней части рисунка посередине. Слева от синапса можно видеть поперечное сечение аксона, заполненного мельчайшими круглыми синаптическими пузырьками, в которых хранится нейромедиатор. Справа от синапса виден выступ дендрита (называемый шипиком); он отходит от крупной дендритной ветви, расположенной горизонтально в верхней части рисунка (два темных колбасовидных образования в этом дендрите — митохондрии). В синапсе сближены две мембраны — мембраны аксона и дендрита; здесь они утолщены и выглядят более плотными. Их разделяет щель шириной 20 нанометров.
Мембрана нервной клетки, покрывающая весь нейрон, — структура чрезвычайно сложная. Она не сплошная, как надувной шарик или шланг, а содержит миллионы «пор», через которые вещества могут переходить с одной стороны на другую. Некоторые из них — это действительно поры различной величины; как сейчас выяснилось, они представляют собой белки в форме трубок, насквозь пронизывающих жировое вещество мембраны. В других случаях это не просто поры, а миниатюрные белковые механизмы, называемые насосами; они способны улавливать ионы одного типа и выбрасывать их из клетки, одновременно захватывая другие ионы внутрь из наружного пространства. Такая перекачка требует затраты энергии, которую клетка в конечном счете получает в процессе окисления глюкозы. Существуют также поры, называемые каналами, — это «клапаны», которые могут открываться и закрываться. Какие воздействия приводят к их открытию или закрытию, зависит от типа пор. На некоторые из них влияет мембранный потенциал, другие открываются или закрываются при наличии определенных веществ во внутренней и наружной жидкости.
Разность потенциалов на мембране в любой момент определяется концентрацией ионов внутри и снаружи, а также тем, открыты или закрыты различные поры. (Выше я говорил, что потенциал влияет на поры, а теперь утверждаю, что поры влияют на потенциал. Давайте пока просто скажем, что эти две вещи могут быть взаимозависимы. Несколько позже будет дано более детальное объяснение.) Так как имеется несколько видов пор и несколько видов ионов, легко понять, что вся система довольно сложна. Когда Ходжкин и Хаксли в 1952 году сумели разобраться в ней, это было огромным достижением.
[banner_centerrs]
{banner_centerrs}
[/banner_centerrs]
Зададимся прежде всего вопросом: как создается разность потенциалов? Предположим, что вначале никакой разности нет и концентрации ионов внутри и снаружи одинаковы. Пусть далее включается насос, который выводит из клетки ионы одного вида, например натрия, и вместо каждого выведенного иона переносит внутрь ион другого вида, например калия. Сам по себе насос не создает какого-либо потенциала, так как сколько положительно заряженных ионов накачивается внутрь, столько же и откачивается (ионы натрия и калия несут одинаковые положительные заряды). Но представим теперь, что по какой-то причине открылось большое число пор одного типа, например калиевых. Ионы калия начнут перетекать по ним, причем скорость потока через каждую открытую пору будет зависеть от концентрации калия: чем больше ионов возле отверстия поры, тем больше будет их утечка через мембрану; а так как внутри ионов калия больше, чем снаружи, то выходить их будет больше, чем входить внутрь. Но если выходит больше зарядов, чем входит, наружное пространство быстро станет электроположительным по отношению к внутреннему. Это накопление положительного заряда снаружи вскоре начнет противодействовать дальнейшему выходу ионов калия из клетки, так как одноименные заряды отталкивают друг друга.
Очень быстро — прежде чем выход ионов К4 приведет к заметному изменению их концентрации — положительный заряд снаружи достигнет величины, при которой он в точности скомпенсирует тенденцию ионов К+ выходить из клетки (с внутренней стороны поры ионов калия больше, но они отталкиваются наружным зарядом). Начиная с этого момента перемещение заряда прекращается, и мы говорим, что система приходит в равновесие. Таким образом, открытие калиевых пор приводит к возникновению на мембране разности потенциалов с положительным полюсом снаружи.
Но предположим, что вместо этого открылись натриевые поры. Повторяя все рассуждения с заменой слов «внутренний» на «наружный», вы можете легко убедиться, что результат будет прямо противоположным: снаружи возникнет отрицательный заряд. При одновременном открытии пор того и другого типа езультат был бы «компромиссным». Для оценки величины мембранного потенциала мы должны знать относительные концентрации двух ионов и отношение числа открытых и закрытых пор для каждого иона, а затем произвести соответствующие расчеты.
Когда нерв находится в покое, большинство калиевых каналов открыто, а большинство натриевых закрыто; поэтому снаружи будет положительный заряд. Во время импульса на коротком отрезке нервного волокна внезапно открывается большое число натриевых каналов, что приводит к кратковременному преобладанию потока ионов натрия, и этот участок быстро становится электроотрицательным снаружи по отношению к внутреннему пространству. Затем натриевые поры вновь закрываются, в то время как калиевые остаются открытыми, причем даже в большем количестве, чем в состоянии покоя. Оба процесса — закрытие натриевых пор и дополнительное открытие калиевых пор — приводят к быстрому восстановлению потенциала покоя с положительным полюсом снаружи. Вся последовательность событий занимает примерно тысячную долю секунды.
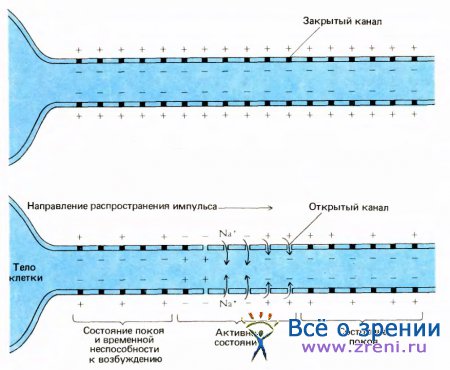
Рис. 10. Вверху: участок аксона в состоянии покоя. Натриевый насос перекачал наружу излишние ионы натрия, а внутрь — недостающие ионы калия. Натриевые каналы в основном закрыты. Поскольку открыто много калиевых каналов, клетку покинуло достаточное количество ионов калия, чтобы мембранный потенциал достиг равновесного в таких условиях уровня — около 70 милливольт с плюсом снаружи.
Внизу: слева направо перемещается нервный импульс. На крайнем правом конце аксон еще пребывает в состоянии покоя. В среднем участке развертываются события, связанные с импульсом: натриевые каналы открыты, ионы натрия переходят внутрь (хотя и не в таком количестве, чтобы их концентрация после одного импульса заметно изменилась); мембранный потенциал 40 милливольт с плюсом внутри На крайнем левом конце мембрана возвращается в исходное состояние, так как открылись (а затем закрылись) добавочные калиевые каналы, а натриевые каналы автоматически закрылись. Поскольку натриевые каналы не способны сразу же повторно открыться, второй импульс не может возникнуть раньше чем спустя примерно миллисекунду. Это позволяет понять, почему импульс не может повернуть назад к телу клетки.
Все происходящее зависит от обстоятельств, влияющих на открытие и закрытие пор. Как натриевые, так и калиевые каналы чувствительны к мембранному потенциалу. Уменьшение наружного положительного заряда — деполяризация мембраны относительно состояния покоя — приводит к открытию пор. Это влияние неодинаково для пор двух типов: натриевые поры, открывшись, снова закрываются сами по себе, даже если мембрана остается деполяризованной, и неспособны вновь открыться на протяжении нескольких тысячных долей секунды; калиевые поры остаются открытыми, пока поддерживается деполяризация. При определенном уровне деполяризации число ионов натрия, входящих внутрь, вначале превышает число выходящих ионов калия и наружная поверхность мембраны становится электроотрицательной по отношению к внутренней; позднее начинает преобладать поток калия и восстанавливается потенциал покоя.
В этой последовательности событий, составляющих импульс (открываются поры, ионы проходят через мембрану и мембранный потенциал дважды претерпевает изменения), число ионов, фактически проходящих через мембрану — Na+ внутрь, а К4 наружу, — ничтожно, и его недостаточно для измеримого изменения ионных концентраций внутри или снаружи клетки. За несколько минут, однако, нейрон способен разрядиться тысячу раз, и в результате концентрации ионов могли бы заметно измениться, если бы не насос, который все время выводит натрий и накачивает внутрь калий, поддерживая таким образом их концентрации на надлежащих уровнях покоя.
Почему во время импульса столь малый перенос заряда ведет к таким большим колебаниям потенциала? Это следствие одного из простых законов электричества: емкость мембраны мала, а потенциал равен перенесенному заряду, деленному на емкость.
Деполяризация мембраны — уменьшение электроотрицательности внутри относительно состояния покоя — вот что обеспечивает первоначальный запуск импульса. Если быстро ввести в покоящееся волокно некоторое количество ионов натрия, вызвав небольшую начальную деполяризацию, то в результате откроется небольшое число натриевых пор; но, поскольку много калиевых пор уже открыто, изнутри может выйти достаточно калия, чтобы скомпенсировать этот эффект и быстро вернуть мембрану в исходное состояние покоя. Предположим, однако, что начальный перенос заряда столь велик и открылось так много натриевых пор, что натрий приносит внутрь больше заряда, чем может быть выведено с калием; тогда мембрана деполяризуется еще сильнее. Это приведет к открытию еще большего числа натриевых пор, к еще большей деполяризации и так далее — возникнет самоусиливающийся взрывоподобный процесс. Когда откроются все натриевые поры, которые могут открыться, мембранный потенциал изменит свой знак на обратный по отношению к потенциалу покоя: вместо 70 милливольт с положительным полюсом снаружи он составит 40 милливольт с отрицательным полюсом снаружи.
Уменьшение потенциала на мембране с последующим изменением его знака (реверсией) не происходит сразу по всей длине волокна, так как перенос заряда требует времени. Активный участок возникает в одном месте и перемещается по волокну со скоростью от 0,1 до примерно 10 метров в секунду. В любой момент времени существует один активный участок с реверсированным потенциалом, и эта область реверсии передвигается, удаляясь от тела нейрона; впереди нее находится участок с еще не открытыми каналами, а позади — участок, где каналы снова закрылись и временно неспособны к повторному открытию.
Это и есть процесс распространения импульса. Вы видите, что он вовсе не похож на прохождение тока по медной проволоке. По всей длине нерва не перемещаются электрические заряды, ионы или вообще нечто материальное, точно так же как при смыкании лезвий ножниц ничто не перемещается от соединительного винта до их кончиков. (Ионы образуют лишь местные токи, переходя внутрь и наружу, подобно тому как лезвия ножниц движутся вверх и вниз.) Перемещается некоторое событие или процесс — перекрещивание лезвий ножниц или импульс в нерве.
Поскольку подготовка натриевых каналов к следующему открытию и закрытию требует некоторого времени, наибольшая частота, с которой клетка или аксон способны генерировать импульсы, составляет около 800 в секунду. Однако столь высокая частота необычна; даже для сильно активированных нервных волокон характерны частоты 100—200 импульсов в секунду.
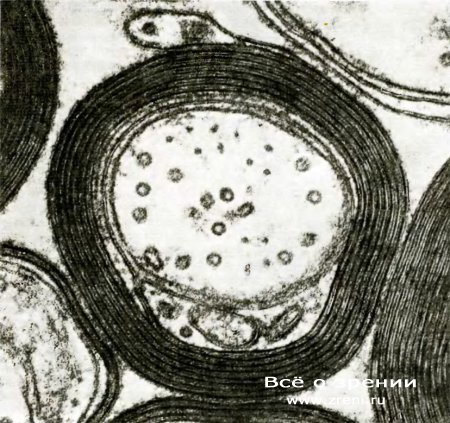
Рис. 11. Мембрана глиальной клетки многократно обернута вокруг аксона, как это видно на электронной микрофотографии поперечного среза нервного волокна. Такая мембрана состоит из миелина, который ускоряет проведение нервных импульсов, повышая сопротивление и уменьшая емкость между внутренностью аксона и окружающим пространством. В аксоне видны (в поперечном сечении) органеллы, называемые микротрубочками.
Важной особенностью нервного импульса является то, что он возникает по принципу всё или ничего. При достаточной начальной деполяризации — если она превышает некоторую пороговую величину — процесс становится само-усиливающимся и реверсия происходит всегда до 0,02 вольт (минус снаружи). Величина потенциала, распространяющегося по нерву (т. е. импульса), определяется самим нервом, а не степенью начальной деполяризации, которая привела к его возникновению. Здесь уместна аналогия с любым взрывообразным процессом. Скорость полета пули никак не связана с тем, насколько резко вы нажали на спусковой крючок.
Для многих функций мозга скорость проведения импульса представляется весьма важной, и нервная система выработала особый механизм ее повышения. Плазматическая мембрана глиальных клеток многократно обертывается вокруг аксона, образуя слоистую оболочку, значительно повышающую эффективную толщину нервной мембраны. Это утолщение снижает емкость мембраны, а тем самым и величину заряда, необходимую для ее деполяризации. Слоистое вещество, богатое жировым материалом, называется миелином. Через каждые несколько миллиметров оболочка прерывается в так называемых перехватах Ранвъе, что позволяет токам, связанным с импульсом, входить в аксон или выходить из него. В результате нервный импульс фактически перескакивает от одного перехвата к следующему, а не передвигается непрерывно вдоль аксона, что намного ускоряет передачу нервных сигналов. Большинство крупных волокнистых пучков в мозгу миелинизировано, что придает им блестящий белый цвет на свежеприготовленных срезах. Белое вещество головного и спинного мозга состоит из миелинизированных аксонов, и в нем отсутствуют тела нервных клеток, дендриты и синапсы. Серое вещество состоит в основном из клеточных тел, дендритов, аксонных окончаний и синапсов, но может содержать и миелинизированные аксоны.
Основные пробелы в нашем понимании природы импульса, равно как и основные направления современных исследований в этой области, связаны со структурой и функцией белковых каналов.
Как первоначально возникают импульсы, и что происходит на дальнем конце аксона, когда импульс прибывает туда?
Участок клеточной мембраны у окончания аксона, образующий первую половинку синапса (пресинаптическую мембрану), обладает удивительной специализированной структурой. Прежде всего, он содержит особые каналы, которые при деполяризации открываются и пропускают положительно заряженные ионы кальция. Поскольку концентрация кальция (как и натрия) снаружи выше, чем внутри клетки, открытие этих каналов позволяет кальцию переходить внутрь. Каким-то пока не известным образом это поступление кальция внутрь клетки приводит к выбрасыванию через мембрану наружу небольших порций особых веществ, называемых нейромедиаторами. Уже идентифицировано около двадцати химических медиаторов, и, судя по темпу новых открытий, их общее число может превышать пол сотни. Молекулы медиаторов намного меньше белковых молекул, но обычно крупнее ионов натрия или кальция. Примерами нейромедиаторов могут служить ацетилхолин и норадреналин. Когда эти вещества высвобождаются из пресинаптической мембраны, они быстро диффундируют через синаптическую щель шириной 0,02 мкм к постсинаптической мембране.
Постсинаптическая мембрана тоже специализирована: в ней имеются белковые рецепторы, которые реагируют на нейромедиатор открытием соответствующих каналов, позволяя ионам одного или нескольких типов проходить через них. От того, какие именно ионы (натрий, калий, хлор) смогут проходить, зависит, будет ли сама постсинаптическая клетка деполяризована или же ее мембранный потенциал будет стабилизирован, т. е. ее деполяризация затруднена.
Подытожим сказанное. Нервный импульс приходит к окончанию аксона и вызывает здесь высвобождение специальных нейромедиаторных молекул. Эти нейромедиаторы воздействуют на постсинаптическую мембрану так, что либо понижают ее мембранный потенциал, либо предотвращают его понижение. При понижении мембранного потенциала частота импульсации возрастает; мы назовем такой синапс возбуждающим. Если же мембранный потенциал вместо этого стабилизируется на подпороговом уровне, импульсы не возникают или возникают с меньшей частотой, и тогда синапс называют тормозным.
Будет ли данный синапс возбуждающим или тормозным, зависит от того, какой в нем высвобождается медиатор и каковы здесь рецепторные молекулы. Ацетилхолин, самый известный медиатор, в некоторых синапсах оказывает возбуждающее действие, а в других — тормозное, он возбуждает мышцы конечностей и туловища, но тормозит сокращения сердца. Норадреналин обычно служит возбуждающим медиатором, гамма-аминомасляная кислота (ГАМК) — тормозящим. Насколько нам известно, характер действия каждого данного синапса остается постоянным на протяжении всей жизни животного.
С дендритами и телом нейрона могут контактировать десятки, сотни или тысячи аксонных окончаний; поэтому в любой момент одни входные синапсы стремятся деполяризовать клетку, а другие противодействуют этому. Импульс, приходящий к возбуждающему синапсу, будет деполяризовать постсинаптическую клетку; если одновременно придет также импульс к тормозному синапсу, эффекты обоих импульсов будут стремиться погасить друг друга. В любой момент уровень мембранного потенциала является результатом сложения вместе возбуждающих и тормозных влияний. Одиночный импульс, приходящий к одному синапсу, обычно лишь очень слабо воздействует на постсинаптическую клетку, и его эффект через несколько миллисекунд исчезает. При поступлении импульсов от нескольких других нейронов клетка суммирует, или интегрирует, их влияния. При достаточном понижении мембранного потенциала — если возбуждающие сигналы приходят к достаточному числу синапсов и с достаточно высокой частотой — суммарная деполяризация сможет привести к возникновению импульсов, обычно в виде целой их серии. Импульсы обычно возникают в том месте, где от тела клетки отходит аксон: деполяризация данной величины здесь с наибольшей вероятностью может вызвать импульс — по-видимому, благодаря особенно высокой плотности расположения натриевых каналов в мембране. Чем сильнее в этом месте деполяризована мембрана, тем больше импульсов возникает каждую секунду.
Почти все клетки нервной системы имеют входные синапсы от нескольких других клеток. Это называют конвергенцией. В то же время аксоны почти всех клеток многократно ветвятся и обслуживают большое число других нейронов — возможно, сотни или тысячи. Мы называем это дивергенцией. Легко видеть, что без конвергенции и дивергенции нервная система немногого бы стоила: возбуждающий синапс просто передавал бы каждый импульс следующей клетке, не выполняя никакой полезной функции, а тормозному синапсу, который был бы единственным входом данной клетки, нечего было бы тормозить, если только постсинаптическая клетка не обладала бы каким-то специальным механизмом, заставляющим ее разряжаться спонтанно.
Мне хотелось бы сделать заключительное замечание о сигналах, передаваемых нервными волокнами. Хотя аксоны почти всегда проводят импульсы по принципу «всё или ничего», имеются некоторые исключения. Если локальная деполяризация оказывается подпороговой, т. е. недостаточной для запуска взрывоподобного импульса типа «все или ничего», она тем не менее имеет некоторую тенденцию распространяться вдоль волокна, уменьшаясь со временем и с расстоянием от начального пункта. (При обычном проведении нервного импульса именно такое локальное распространение и доводит потенциал соседнего, покоящегося участка нервной мембраны до пороговой деполяризации, при которой начинается самоусиливающийся процесс.) Некоторые аксоны столь коротки, что не возникает нужды в распространяющихся импульсах: деполяризация клеточного тела или дендритов путем пассивного распространения способна создавать в окончании аксона деполяризацию, достаточную для высвобождения медиатора. У млекопитающих известные случаи передачи информации без импульсов немногочисленны, но важны. В наших сетчатках нервные клетки двух или трех из пяти типов функционируют без импульсов.
Важное отличие этих пассивно передаваемых сигналов от импульсов (помимо их малой и прогрессивно убывающей амплитуды) состоит в том, что их величина меняется в зависимости от силы стимула. Поэтому их часто называют градуальными сигналами. Чем больше сигнал, тем сильнее деполяризация окончания и тем больше высвобождается медиатора. Следует напомнить, что импульсы, напротив, не увеличиваются по амплитуде с ростом стимула — вместо этого возрастает частота их повторения. И чем чаще возникают импульсы, тем больше медиатора выделяется в окончаниях. Таким образом, конечный результат не слишком различен. Нередко говорят, что градуальные потенциалы представляют собой пример аналоговых сигналов, а импульсы, будучи событиями типа «все или ничего», носят характер цифровой сигнализации. Я нахожу эту аналогию неверной, поскольку точное положение каждого импульса в серии в большинстве случаев не имеет никакого значения. Что важно, так это среднее число их в данном временном интервале, а не тонкие детали. Таким образом, оба вида сигналов существенно аналоговые.
Теперь, когда мы кое-что знаем об импульсах, синапсах, возбуждении и торможении, можно задаться вопросом, как из нейронов строятся более крупные образования. Мы можем представить центральную нервную систему — головной и спинной мозг — как некоторый ящик со входом и выходом. Входные сигналы оказывают воздействие на особые нервные клетки, называемые рецепторами; эти клетки реагируют не на синаптические входы от других клеток, а на то, что мы можем обозначить нестрогим термином «внешняя информация». Эта информация может принимать форму света, проникающего в наши глаза; механической деформации нашей кожи, барабанной перепонки или полукружных каналов; химических веществ, как в наших органах обоняния или вкуса. Во всех этих случаях под влиянием стимула в рецепторах возникает электрический сигнал и в результате изменяется скорость высвобождения медиатора в окончаниях их аксонов.
(Вас не должно смущать двойное значение термина рецептор; вначале он означал клетку со специализированной реакцией на сенсорные стимулы, но впоследствии был применен также к белковым молекулам со специализированной реакцией на нейромедиаторы.)
На другом конце нервной системы мы имеем выход — двигательные нейроны, отличающиеся от всех остальных тем, что их аксоны оканчиваются не на других нейронах, а на мышечных клетках. Все выходные сигналы нашей нервной системы вызывают мышечные сокращения, за редким исключением сигналов, адресуемых клеткам желез. Это, по-существу, единственный способ, которым мы можем оказывать влияние на окружающую нас среду. Отключите у какого-либо животного мышцы, и вы полностью отрежете его от остального мира; отключите вместо этого входные сигналы — и прекратятся все внешние влияния, что опять-таки превратит животное фактически в растение. Согласно одному из возможных определений, животное — это организм, реагирующий на внешние события и своими действиями оказывающий влияние на внешний мир.
Рис. 12. Нервно-мышечное соединение у лягушки. Более тонкое нервное волокно извивается рядом с двумя мышечными волокнами, образуя синапс в нижней левой части снимка.
Центральная нервная система, лежащая между входными и выходными нейронами, является тем аппаратом, который позволяет нам воспринимать, реагировать и помнить; он же в конечном счете должен быть ответствен за наше сознание, совесть и душу. Одна из главных целей нейробиологии состоит в выяснении того, что происходит на этом пути — как информация, приходящая к некоторой группе нейронов, преобразуется и затем передается дальше, а также какой смысл имеют эти преобразования для успешной адаптации животного к среде.
Хотя схемы связей для многих отделов центральной нервной системы значительно варьируют в деталях, в большинстве случаев в основе их лежит сравнительно простой общий план, показанный на рис. 13.
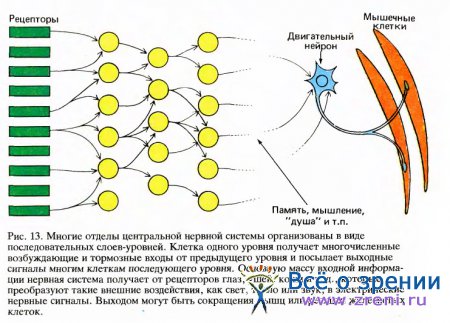
Приведенная здесь схема — это скорее карикатура, которую не следует понимать буквально и которая требует уточнений, обсуждаемых ниже. Слева на рисунке показаны рецепторы — ряд преобразующих информацию нейронов, каждый из которых обслуживает один из видов ощущений, например осязание, вибрационное чувство или зрительное восприятие. Мы можем считать эти рецепторы первым уровнем какого-то сенсорного пути. Волокна от рецепторов образуют синаптические контакты со вторым рядом нервных клеток — вторым уровнем нашей схемы; эти клетки в свою очередь образуют контакты с третьим уровнем и так далее. «Уровень» не является техническим или широко применяемым нейро-анатомическим термином, но мы еще увидим его полезность.
Иногда три или четыре таких уровня объединяются вместе в более крупный элемент, который я за недостатком лучшего или общепринятого термина назову структурой. Структуры — это скопления клеток, обычно в виде пластин или сферических ядер, упоминавшихся в главе 1. В случае пластинчатой структуры каждый из образующих ее уровней может быть дискретным слоем клеток. Хорошим примером служит сетчатка, в которой три таких слоя и, грубо говоря, три уровня. Если несколько уровней группируются, образуя более крупную структуру, то нервные волокна, приходящие к ней от предыдущей структуры и идущие от нее к следующей, обычно группируются в пучки, называемые трактами.
На схеме (рис. 13) можно видеть, насколько распространены процессы дивергенции и конвергенции сигналов: аксон почти каждой клетки данного уровня ветвится при подходе к следующему уровню и оканчивается на нескольких или многих клетках; и наоборот, клетка любого уровня, кроме первого, имеет синаптические входы от нескольких или многих клеток предыдущего уровня.
Очевидно, что мы должны усовершенствовать и конкретизировать эту упрощенную схему, и у нас для этого есть некоторая отправная модель. Прежде всего на входном конце мы имеем не одну, а много сенсорных систем — зрение, осязание, вкус, обоняние и слух, и каждая система имеет свой собственный набор уровней в мозгу. Когда и где в мозгу различные наборы уровней сводятся вместе, если они действительно сводятся, все еще неясно.
Прослеживая одну систему, например зрительную или слуховую, от рецепторов до мозга, мы можем обнаружить, что она разделяется на отдельные подсистемы. В случае зрения эти подсистемы имеют отношение специально к движениям глаз, сокращению зрачка, восприятию формы, движения, глубины или цвета объектов. Таким образом, вся система дивергирует на отдельные каналы (пути); таких каналов может быть много и они могут сильно разниться по своей длине. Уже сразу видно, что некоторые пути содержат много структур на своем протяжении, а другие — мало. При более детальном анализе оказывается, что какой-то аксон одного уровня может идти дальше, не заходя на один или два последующих уровня сенсорной системы, и даже прямо направиться к двигательному нейрону. (Такой пропуск уровней в нейроанатомии аналогичен тому, что часто случается в генеалогии. Современный английский монарх не связан с Вильгельмом Завоевателем однозначным числом поколений: число приставок «пра» к слову «дед» неопределенно из-за родственных браков между племянниками и тетками и других еще более сомнительных обстоятельств.)
Если путь от входа к выходу очень короток, мы называем это рефлексом. В зрительной системе сужение зрачка при ярком свете служит примером рефлекса, в котором число синапсов, вероятно, около шести. В наиболее простом случае аксон от рецептора оканчивается прямо на двигательном нейроне, так что мы имеем от входа к выходу всего три клетки (рецептор, двигательный нейрон и мышечное волокно) и только два синапса; такую ситуацию мы называем моносинаптической рефлекторной дугой. (Возможно, человек, сочинивший этот термин, не рассматривал нервно-мышечное соединение как настоящий синапс или не умел считать до двух.) Этот короткий путь активируется, когда врач стучит молоточком по вашему колену и нога подпрыгивает. Джон Николлс обычно говорил своей аудитории в Гарвардской медицинской школе, что есть только два резона проверять этот рефлекс: убить время и проверить, нет ли у больного сифилиса.
На выходном конце мы находим не только различные группы мышц, расположенные на туловище, в конечностях, глазах и языке, которыми мы можем управлять произвольно, но также мускулатуру, обслуживающую менее произвольные или непроизвольные, «хозяйственные» функции, такие как взбалтывание содержимого желудка, мочеиспускание или дефекация, а в промежутках между этими актами — закрытие выводных протоков с помощью сфинктеров.
Мы должны также дополнить нашу модель путями обратной передачи информации. На нашей схеме (рис. 13), разумеется, доминирует направление слева направо, от входа к выходу. Но почти во всех случаях, когда сигналы передаются от одного уровня к следующему, возвратные связи направляют их также назад, со второго уровня на первый. (Иногда мы можем догадываться, для чего такая обратная связь могла бы служить, но почти во всех случаях у нас нет на этот счет ясного понимания.) Наконец, даже в пределах какого-то одного уровня мы часто обнаруживаем богатую сеть соединений между соседними клетками. Таким образом, утверждение, что та или иная структура содержит определенное число уровней, почти всегда будет чрезмерным упрощением.
Когда я начинал работать в области неврологии в начале 50-х годов, этот основной план нервной системы был хорошо известен. Но в те дни никто не имел четкого представления о том, как понимать организацию этой информационной эстафеты по цепи уровней. Сейчас мы значительно больше знаем о способах преобразования информации в некоторых частях мозга; о других частях нам еще почти ничего не известно. Оставшиеся главы этой книги посвящены зрительной системе, которую мы сегодня понимаем лучше всего. Ниже я попытаюсь дать обзор некоторых фактов, известных нам об этой системе….
—
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.
ФИЗИКА ЖИЗНЕННЫХ ПРОЦЕССОВ
АКСОН И НЕРВНЫЙ ИМПУЛЬС
Генерация и распространение нервного импульса в организмах животных, возбуждение нервных и мышечных клеток — важнейшие мембранные явления. Мембранная теория возбуждения была сформулирована Бернштейном еще в 1902 г. [1]. Согласно его теории, возбуждение определяется электрохимическими процессами, локализованными в мембранах нервных и мышечных клеток. Эти процессы сводятся к перемещению малых ионов. Позднее Лазарев опубликовал монографию об ионной природе нервного возбуждения [2]. Исследования, проведенные Ходжки- ном, Катцем, Хаксли, Тасаки и другими учеными [3—6], раскрыли принципиальные механизмы генерации нервного импульса и его распространения (см. также [7—9]).
Нервное возбуждение распространяется по нервным волокнам— аксонам. Принято разделять нервную систему высших организмов на центральную и периферическую. Периферическая система содержит аксоны, служащие для передачи сигналов, а также ганглии вегетативной нервной системы. Аксоны являются коммуникациями для афферентных сообщений от органов чувств, направляемых в центральную систему, и для эфферентных сигналов, направляющихся от центральной системы к мышцам. Аксоны представляют собой отростки центрально расположенных клеток. Нервная система беспозвоночных имеет иное строение, но она также содержит коммуникационные аксоны.
Исследование генерации нервного импульса и его распространения в аксоне — старая и традиционная проблема биофизики/ Уже Гельмгольц измерял скорость распространения нервного возбуждения/В настоящее время решены важные физические задачи, относящиеся к функции аксона. Напротив, современное состояние науки позволяет лишь формально моделировать работу центральной нервной системы и мы еще далеки от понимания физической природы высших ее функций — памяти и мышления.
На рис. 4.1, заимствованном из [5], представлена схема строения нервной клетки, именуемой иначе нейроном. Нейрон полу
чает сообщения от многих других нейронов через их тонкие ответвления, образующие контакты — синапсы — с телом клетки и его короткими отростками — дендритами. Аксон имеет макроскопические протяженность и сечение. Длины аксонов в теле крупных животных достигают нескольких метров.
Контакты с другими клетками образуются не только в синапсах.^ Боль — шая часть поверхности нейрона покрыта прилегающими к нему глиальными, или иіванновскими, клетками^ роль которых пока не выяснена. С другой стороны^из мембран шванновских клеток образуется миелиновая оболочка миелинизирован — ных аксонов, показанная ^схематически на рис. 4.1. ^Оболочка прерывается через каждые 1—2 мм длины аксона перехватами Ранвье, имеющими протяженность /Т около 1 мкм. В области перехватов мембрана аксона непосредственно контактирует с окружающей средойЛ^
Немиелинизированные во — ЛОКНа могут быть объеди — Рис. 4.1. Схема строения нервной клетки нены одной шванновской (двигательный нейрон).
К-ТТР. ТКПЙ чтп ППКЯ1ЯНП R гр — і тн~тел0 нейрона,1Ца — аксои1Смо — миелиновая клеткой, ЧТО UOKdJdHU В се ~ оболочка, перехват Ранвье^д-дендрит,
ЧЄНИИ на рис. 4.2. Погру — X*— ядро,^с —синапсыЛодн—окончание двига о г — г тельного нерва,мв — мышечное волокно.
Женныи в шванНЬвскую 1

Клетку аксон создает глубокую инвагинацию мембраны шванновской клетки, причем образуется мезаксон — две шванновские мембраны, расположенные рядом^В правом нижнем углу схематически показана двойная мембрана, разделяющая цитоплазму шванновской клетки и цитоплазму аксона, аксоплазму [9].
Герен [10] установила, что миелиновая оболочка аксона образована многократным спиральным закручиванием мембраны шванновской клетки вокруг аксоиаГМиелин представляет собой, — т-а«им~ойразом,/многослойную (доЧїбО слоев) мембрану, строение которой наблюдается с помощью электронного микроскопа. Миелии защищает мембрану аксона от окружающей среды.
6 М. В. Волькеиштейн
Структура миелина была детально изучена методом рентгенографии с разрешением 10 А [11]. Определены профили электронной плотности миелина седалищного нерва кролика и лягушки и миелина зрительного нерва кролика. На рнс. 4.3 показаны эти кривые для нервных волокон кролика и схема структуры миелина, соответствующая данным кривым. Миелин построен асимметрично, холестерин сосредоточен преимущественно на стороне

Рис. 4.2. Схема строения немиелинизированнного нервного волокна позвоночного.
А — аксон, м — мезаксон— шванновская клетка.
Миелина, соприкасающейся с мембраной аксона. Схема согласуется с липидным составом миелина млекопитающих, установленным ранее [12]. Белковые слои граничных областей миелина содержат йоду. Внутренняя область построена из гибких углевОдорОдных «хвостов» липидов.
/Уже давно установлено, что нервы и мышцы способны генерировать электродвижущие силы, биопотенциалы. Электрическая активность клеток проявляется в форме коротких разрядов, каждый из которых длится около 1 мс. Современная экспериментальная техника позволяет усиливать эти сигналы и регистрировать их с помощью осциллографа или другим путем. Установлено, что активность нерва всегда сопровождается электрическими явлениями — и при внешнем возбуждении и при посылке сигнала мозгом в соответствующем аксоне наблюдаются электрические импульсы. На рис. 4.4 показаны импульсы в зрительном нервном волокні краба Limulus, вызванные вспышкой света длительностью 1 с [13]. Частота и характер последовательности импульсов, имеющие несомненное информационное значе
ние, зависят от интенсивности и спектрального состава света [14, 15] (см. гл. 7) .1 Величина и длительность отдельного импульса не зависят отТгрироды и силы раздражения.»
О 20 40А
-J 1 1 1 1

0 0 .::•:
![]()
»w0#0
^ V J 1 O^^-v^.p..
Г У. Го 0

Внугрен^/7о/тярже=Стероид щ^Гибх/гя ^Стероид ^//плярная Внешний
Нийбеяоцт. группы^ +ягесткая цепь +жесткая белок
+еоЗа ‘— » цепь — цепь — —~ +вода углеводороды
Рис. 4.3. Схема миелиновой мембранной структуры.
Вверх* — профили электронной плотности для глазного (пунктир) н седалищного (сплошная
Лнння) нервов кролика.
Электрический сигнал, отвечающий отдельному импульсу, распространяющемуся вдоль аксона, называется потенциалом действия или спайком. Это — основная единица информации, передаваемой по нервному волокну.

Скорость распространения импульса по аксону равна по порядку величины 1—100 м/с, она меньше для немиелинизиро — ванных и больше для миелинизированных аксонов.


0,01
0,001
Рис. 4.4. Импульсы в зрительном нервном волокне Limulus, вызванные вспышкой света длительностью I с.
Цифры справа указывают относительную интенсивность вспышки. Период освещения указан разрывом верхней белой линии, отметка времени на нижней белой линии 0,2 с.
В табл. 4.1 [3] приведены некоторые данные. Проведение нервного возбуждения есть проведение электрического импульса. Несмотря на то, что аксоплазма представляет
Таблица 4.1
Скорость проведения возбуждения в нервном н мышечном волокнах
|
Волокно |
Температура, °С |
Диаметр волокна мкм |
Скорость проведения, м/с |
|
Миелинизированные нервные волок |
38 |
2-20 |
10-100 |
|
На кошки |
|||
|
Немиелинизированные нервные во |
38 |
0,3-1,3 |
0,7—2,3 |
|
Локна кошки |
|||
|
Миелинизированные нервные волок |
20 |
3-16 |
6-32 |
|
На лягушки |
35 |
||
|
Миелинизированные нервные во |
20 |
20 |
|
|
Локна креветки |
|||
|
Немиелинизированные нервные во |
20 |
30 |
5 |
|
Локна краба |
|||
|
Гигантский аксон кальмара (немие- |
20 |
500 |
Около 25 |
|
Линизированное волокно) |
|||
|
Мышечное волокно лягушки |
20 |
60 |
1,6 |
‘) Для миелинизированных волокон указаны значения наружного диа^ метра миелиновой оболочки.
Собой раствор электролитов, никоим образом нельзя считать, что ионный ток в ней определяет распространение импульса. («Удельное сопротивление аксоплазмы равно по порядку величины 10—100 Ом-см. Следовательно, сопротивление на единицу длины волокна диаметром 1 мкм составляет 109—Ю10Ом-см, т. е. превышает сопротивление медного провода того же диаметра в 108 раз. В таком проводнике очень велики потери и утечки. Между тем аксон передает первичный импульс на расстояния свыше метра б^а-затухания. и без иска^кения.^.
В нейрофизиологии и в биофизике нервнЪ’го»»возбуждения сыграло большую роль развитие методов работы на изолированных аксонах, техники введения микроэлектродов в аксон и метода перфузии, т. е. выдавливания аксоплазмы из волокна и ее замены искусственными растворамиДОсобенно удобна работа на гигантских аксонах кальмара, имеющих диаметр до 0,5 мм (см. табл. 4.1 ^Элементарный опыт состоит во введении в аксон двух микроэлектродов: первый служит для электрической стимуляции возбуждения, второй-^- для измерения генерируемого потенциала (рис. 4.5) [4]./Опыт показывает, что «»величина и временной ход токов действия совершенно не зависят от величины стимулирующего тока и что потенциал действия не возникает, если электрический стимул не достигает некоторого порогового потенциала. Тем самым, нервное волокно
Гл. 4. нервный импульс
Подчиняется закону «все или ничего». Иными словами, потенциал действия постоянной величины либо возникает, либо не возникает вовсе.
Для возбуждения аксона необходимо некоторое минимальное количество электричества.,’ Соответственно по мере уменьшения
Раздражение Осцилмерар
ЕЖЖ^,..»,, ,
Аксон
Рис. 4.5. Схема исследования нервного импульса при помощи двух микроэлектродов.
Длительности At подаваемого импульса необходимо увеличивать силу тока / — эти две величины обратно пропорциональны. Для длительных прямоугольных импульсов существует минимальная
Сила тока, достаточная для возбуждения; более слабый ток неэффективен при любой длительности.
Fta рис. 3/S показана кривая зависимости силы порогового тока от длительности импульса. Кривая имеет гиперболическую форму. Было предложено описывать ее эмпирической формулой
/п = а/At — f b,
Где Іп— пороговый ток, At— длительность импульса, b — так называемая реобаза — минимальное значение возбуждающего тока при At-> оо. При очень коротких импульсах, т. е. при At—>-0 /п = а/At, т. е. произведение IaAt есть константа, характеризующая пороговое значение количества электричества при коротких импульсах. Из приведенной формулы следует, что при /п = 2b получим А^хр = а/Ь. В действительности, однако, оказалось, что при At = ajb /п~1,4Ь и простая эмпирическая формула несправедлива во всем интервале значений A^.j
166

Рисі 4.6. Зависимость силы порогового тока от длительности импульса.
Если сообщить волокну два последовательных стимула, разделенных некоторым интервалом времени, то возникающая ситуация зависит от этого интервала. Немедленно после генерации импульса данное место волокна находится в абсолютном
Рефрактерном состоянии, т. е. не может быть возбуждено вновь. Затем следует относительное рефрактерное состояние. В таком состоянии заметно увеличено значение порогового потенциала и может возникнуть лишь медленно распространяющийся импульс. Продолжительность всего рефрактерного состояния варьирует от одной до нескольких миллисекунд.
В состоянии покоя аксоплазма имеет отрицательный заряд по отношению к внешней среде, характеризуемый потенциалом покоя около 80 мВ. Пороговое возбуждение при этом имеет потенциал, примерно равный —50 мВ, а потенциал действия при надпороговом возбуждении равен потенциалу противоположного знака, составляющему около 40 мВ. Это изображено схематически на рис. 4.7 [4].
Прохождение импульса определяется изменением состояния мембраны аксона при ее стимуляции. Как уже говорилось, в состоянии покоя клеточной мембраны в результате активного транспорта ионов концентрация ионов К+ в цитоплазме (аксо — плазме) значительно превосходит их концентрацию во внешней среде, а для ионов Na+ ситуация противоположна^ В табл. 4.2 [3] приведен состав аксоплазми аксонов Loligo и состав плазмы крови, характерный для внешней среды, и близкий к нему состав морской воды.
Таблица 4.2 Концентрация нонов н других веществ в аксонах Loligo
Концентрация, ммоль на 1 кг НаО
|
Вещество |
Морская |
||
|
Аксоплазма |
Кровь |
Вода |
|
|
К+ |
400 |
20 |
10 |
|
Na+ |
50 |
440 |
460 |
|
Сг |
40—150 |
560 |
540 |
|
Са++ |
0,4 |
10 |
10 |
|
Mg++ |
10 |
54 |
53 |
|
Изэтионовая кислота |
250 |
— |
— |
|
Аспарагнновая кислота |
75 |
— |
— |
|
Глутаминовая кислота |
12 |
— |
— |
|
Янтарная н фумаровая кислоты |
17 |
— |
— |
|
Ортофосфат |
2,5-9 |
— |
— |
|
АТФ |
0,7—1,7 |
— |
— |
|
Аргннннфосфат |
1,8-5,7 865 г/кг |
— |
— |
|
Вода |
870 г/кг |
966 г/кг |
При наложении на мембрану стимулирующего потенциала вначале увеличивается ее проницаемость для ионов Na+, в состоянии покоя значительно меньшая проницаемости для ионов К+. Ионы Na+ входят в аксон, в результате чего внутренняя поверхность мембраны изменяет знак своего заряда с отрицательного на положительный. Иными словами, происходит деполяризация мембраныЛЕсли воспользоваться для выражения разности потенциалов на двух сторонах мембраны аксона формулой (3.105)
+ + (4.1)
^ РКСК + PNacNa + РС1сС ‘
То состояние покоя мембраны характеризуется следующим отношением ионных проницаемостей:
Рк ■ ^Na •’ Реї = 1 : 0,04:0,45.
В состоянии возбуждения, отвечающем генерации потенциала действия, имеем
Рк: Pnа: Ра = 1 :20: 0,45.
Подчеркнем, что истолкование возбуждения как результата изменения ионных проницаемостей является феноменологическим и не раскрывает молекулярный механизм процесса.
Наружная среда
![]()
Активный участок
+ + + — — — — + + + + +
Импульс Э-
Электрические токи, возникающие при деполяризации мембраны аксона.
При деполяризации мембраны возникают токи, замыкающиеся через наружную проводящую среду (рис. 4.8) [5]. Они возбуждают следующий участок аксона.
Таким образом, согласно мембранной теории при генерации импульса в мембране аксона открывается некий «канал», сквозь который ионы Na+ проникают внутрь аксона, вызывая деполяризацию мембраны. Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Ионы К+ выходят наружу, что приводит к восстановлению нормального отрицательного заряда на внутренней стороне мембраны. Происходит реполяризация мембраны. После прохождения рефрактерного периода мембрана приобретает способность передавать новый импульс (рис. 4.9) (4].
+40 О
-за
3
Оотенциол действия
Птещиал пот
+ к — — у + + + + + + + + + + + +
Їфі TtII ‘МрЖ. 111111
+ T —+ + + + + + + + + + + ®
+40
![]()
О
+ + + + + + + + І » + + + + + + + + + + + + + + ї—+ + + + + +
+40 О
-ВО

Рис. 4.9. Распространение нервного импульса сопровождается изменениями нонной проницаемости мембранного аксона.
Наружный раствор
/ / і ±
А/гсоплаэма
Рнс. 4.10. Эквивалентная схема элемента возбудимой мембраны аксона.
Как видно из рис. 4.8, суммарный продольный ток через сечение аксона и окружающую среду равен нулю — в любом месте внутренние токи равны по силе и противоположны по направлению. Но плотность продольного тока и продольная разность потенциалов между двумя точками внутри аксона иные, чем снаружи.
Мембрана аксона имеет сопротивление 1000 Ом-см2, ее емкость равна 1 мкФ/см2, что соответствует бимолекулярному липидному слою толщиной 50 А с диэлектрической проницаемостью е=5 и удельным сопротивлением 2-Ю9 Ом-см. Во время генерации импульса проводимость мембраны увеличивается примерно в 1000 раз. Можно моделировать электрические свойства мембраны эквивалентной схемой, показанной на рис. 4.10 [3]. Рисунок изображает лишь один элемент мембраны, и следует представить себе длинную линейную последовательность таких элементов, моделирующих непрерывный кабель. Сопротивление R характеризует аксоплазму, наружный раствор имеется в большом избытке и изображается проводником без сопротивления. Натриевая и калиевая «батареи» и S’k,
Определяют генерацию импульса, добавочная «батарея» Sy ответственна за движение других ионов, не изменяющееся при возбуждении.
Таким образом, распространение импульса представляет собой самоподдерживающийся процесс, подобный горению бикфордова шнура. Очевидно, что процесс этот сводится к превращению химической энергии в электрическую.
Как уже сказано, распространение импульса в миелинизированном волокне происходит быстрее, чем в немиелинизированном. Доказано, что в миелинизированном аксоне проведение импульса является сальтаторным, т. е. происходит путем перескока импульса от одного перехвата Ранвье к другому. Калиевые и натриевые каналы открываются и закрываются только в перехватах Ранвье — миелинизированные участки хорошо изолированы. Миелин имеет малую емкость, чем и объясняется большая скорость проведения импульса. Схема сальтаторного проведения импульса показана на рис. 4.11. ^
![]()
РирГ 4.Т7Т^ Схема сальтаторного проведения импульса. пР—перехват Ранвье, л —миелин, а — аксо-
Прямые доказательства сальтаторной їеории, впервые предложенной Лилли [16], были получены в работах Кубо, Оно и Юге (см. [17]). В одиночном нервно-мышечном препарате, погруженном в раствор Рингера, пороговая величина стиму
лирующего тока зависит от расстояния между стимулирующим микрозлектродом и перехватом Ранвье. Она имеет наименьшее значение, если микрозлектрод помещен непосредственно в перехвате. Количественная теория этого эффекта была развита Та — саки (см. [6]).
Обозначим потенциалы трех последовательных перехватов — 1, 0 и 1 буквами ifi_i, if)0 и ij)j. Если миелин представляет собой идеальный изолятор, то ток, протекающий внутри аксона от 1 к 0, пропорционален if>i— if>o, от 0 к —1 — пропорционален ■ф-i — if>o. Через перехват 0 идет ток І, пропорциональный (грі — ij)0) — f (j)_i — %,). Имеем
‘ = І{(+-,-*о) + (+,-+о)}. (4.2)
Где R — сопротивление между двумя соседними перехватами (оно равно примерно 20 МОм).
Расстояния между микроэлектродом и перехватами 0, 1 и —1 равны Хо, Х, х-. Разность потенциалов, создаваемая микроэлектродом, дающим ток I, равна
Где р — удельное сопротивление среды (для раствора Рингера оно равно 100 Ом-см). Из (4.2) и (4.3) получаем
TOC o «1-3» h z i = Ч (4-4)
4яR х0 х х-1 ) 4 ‘
И если микроэлектрод очень близок к перехвату 0, т. е. Хи Х-1 » *0, то
= (4-5)
В опытах варьировалось расстояние х0 и при каждом значении Хо определялась величина /, отвечающая критическому значению і — іс. Опытные данные действительно согласуются с линейной зависимостью I от х0
І= — Ісх0. (4.6)
Установлено, что факторы, блокирующие распространение импульса (кокаин, уретан, ультрафиолетовое облучение), действуют именно на перехваты Ранвье, но не на участки между ними.
Прямые и строгие доказательства сальтаторной теории получены в работах Тасаки и его сотрудников [6, 18, 19]. Основной опыт ставился на одиночном волокне, лежащем в трех каплях раствора Рингера, разделенных двумя воздушными промежутками, расстояние между которыми равнялось приблизительно 1 мм (рис. 4.12) [3]. В первом опыте (рис. 4.12, а) в средней капле находилась только миелинизированная часть аксона, во втором (рис. 4.12,6)—в средней капле находился перехват Ранвье. Измерялся радиальный ток. Через сопротивление R проходит ток, равный il2— г23, где i2— ток, текущий в наружной цепи от капли 1 к капле 2, і23— от капли 2 к капле 3. Эта разность равна току, входящему в волокно из капли 2.

1ме 1ма
Рис. 4.12. Мембранный ток:
А—через участок аксона, расположенный между перехватами Раивье, б —через перехват Ранвье. Отклонение осциллограммы вверх соответствует выходящему току.
В первом опыте радиальный ток направлен наружу, что объясняется пассивным разрядом миелинизированного участка через перехват. Правый перехват разряжается позже левого, соответственно на осциллограмме видны два пика. Во втором опыте первая фаза тока направлена наружу, что означает пассивную деполяризацию перехвата. Во второй фазе ток направлен внутрь — перехват Ранвье находится в активном состоянии. Эти опыты однозначно подтверждают теорию (дальнейшие подробности см. в [3, 6, 18, 19]).
Мембранная теория в целом непосредственно подтверждается опытами с перфузией аксонов. Оказалось возможным выдавить аксоплазму из гигантского аксона Loligo без повреждения мембраны и заменить аксоплазму искусственными растворами (см. [3, 6, 20—22]). Перфузированные аксоны способны проводить до 105—106 импульсов в течение нескольких часов.
Потенциал покоя исчезает при равенстве наружной и внутренней концентраций калия. При замене в аксоплазме КС1 на NaCl потенциал покоя падает до нуля. В то же время потенциал покоя малочувствителен к концентрациям К+, меньшим 20 мМ (см. также § 3.6). Перфузионные опыты показывают, что потенциал покоя действительно регулируется ионами калия.
При указанной замене К. С1 на NaCl и значительном увеличении концентрации К (до 600 мМ) создается положение, обратное нормальному, и внутренняя часть волокна заряжается положительно, а не отрицательно по отношению к наружной среде. И в самом деле, при заполнении волокна, погруженного в изотонический раствор КС1, таким же раствором NaCl внутренний раствор заряжен положительно по отношению к наружному и <р составляет 50—60 мВ.
Прямые опыты с мечеными атомами показывают, что проведение импульса связано с возрастанием скорости движения калия и натрия по градиентам концентрации. В гигантском аксоне каракатицы Sepia при каждом импульсе наблюдается входящий поток Na+, равный 10,3-Ю-12 моль/см2, и выходящий поток 6,6-Ю-12 моль/см2. Чистый вход Na+ 3,7-Ю-12 моль/см2 за импульс примерно равен выходу К+. Действительно, для изменения напряжения конденсатора емкостью 1 мкФ на 120 мВ нужен заряд 0,12-Ю-6 Кл, что эквивалентно 1,3-Ю-12 моль/см2 одновалентного катиона. Таким образом, измеренный вход Na+ более чем достаточен для возникновения потенциала действия.
За один импульс в немиелинизированном аксоне через 1 мкм2 поверхности проходит около 20 000 ионов Na+. В миелинизиро — ванном аксоне за один импульс через каждый перехват Ранвье входит 6-Ю6 ионов Na+. Площадь мембраны в перехвате примерно равна 20 мкм2, следовательно, через 1 мкм2 проходит 300 000 ионов Na+. Плотность ионного тока в перехвате примерно в 10 раз больше, чем в немиелинизированных гигантских аксонах.
Мембранная теория встречается, однако, с трудностями при попытках объяснить тепловые явления в нерве. Согласно мембранной теории генерация импульса происходит за счет свободной энергии, определяемой градиентом концентрации ионов Na+. Если перенос ионов адиабатичен, то система, совершая электрическую работу, должна охлаждаться. Однако оказалось, что во время генерации импульса сначала происходит нагревание нерва, за которым следует охлаждение [23]. Аналогичное явление наблюдается при разряде электрического органа угря на внешнее сопротивление [24]. Эти тепловые явления до сих пор не объяснены. Несомненно, что мембрана аксона представляет собой диссипативную систему, и можно думать, что к рассмотрению генерации нервного импульса применима неравновесная термодинамика. Соответствующая теория еще не построена, она должна описать и теплопродукцию нерва. Подлинное объяснение этих явлений требует, однако, раскрытия их молекулярных механизмов.
В заключение приведем электрические характеристики мие- линизированного нервного волокна лягушки (табл. 4.3) [3].
Таблица 4.3
Электрические характеристики аксона
|
Диаметр волокна |
14 мкм |
|
Толщина миелиновой оболочки |
2 мкм |
|
Расстояние между перехватами Ранвье |
2 мм |
|
Площадь мембраны в перехвате Ранвье |
22 мкм2 |
|
Сопротивление на единицу длины осевого ци |
140 МОм/см |
|
Линдра |
110 Ом • см |
|
Удельное сопротивление аксоплазми |
|
|
Емкость на единицу длины миелиновой оболочки |
10—16 пФ/см |
|
Емкость на едниицу площади миелиновой обо |
0,0025—0,005 мкФ/см2 |
|
Лочки |
|
|
Диэлектрическая проницаемость миелиновой обо |
5-10 |
|
Лочки |
|
|
Удельное сопротивление миелиновой оболочки |
500—800 МОм/см |
|
Емкость перехвата Ранвье |
0,6-1,5 пФ |
|
Емкость на единицу площади перехвата Ранвье |
3—7 мкФ/см2 |
|
Сопротивление перехвата Ранвье в покое |
40—80 МОм |
|
Сопротивление, умноженное на единицу площади |
10-20 Ом-см2 |
|
Мембраны в перехвате Ранвье |
|
|
Потенциал действия |
116 мВ |
|
Потенциал покоя |
71 мВ |
|
Максимальная плотность входящего тока |
20 мА/см2 |
|
Скорость проведения |
23 м/с |
В биологии особое значение имеют автокаталитические химические системы. Достаточно указать, что авторепродукция КДеток и организмов эквивалентна автокатализу. Вернемся сначала к феноменологическому термодинамическому рассмотрению. Как мы видели, для химических процессов критерий …
Неотъемлемой особенностью биологических объектов — клеток и организмов — является их историчность, т. е. возникновение и развитие изучаемой системы в конечном интервале времени. Развитие биологической системы всегда необратимо, и в …
Самые общие соображения показывают, что перенос электрона, сдвиг электронной плотности в конденсированной ср. еде должны сопровождаться изменениями положений атомов, атомных ядер среды. Все степени свободы молекулярной системы, т. е. системы, …
НЕРВНАЯ СИСТЕМА, сложная сеть структур, пронизывающая весь организм и обеспечивающая саморегуляцию его жизнедеятельности благодаря способность реагировать на внешние и внутренние воздействия (стимулы). Основные функции нервной системы – получение, хранение и переработка информации из внешней и внутренней среды, регуляция и координация деятельности всех органов и органных систем. У человека, как и у всех млекопитающих, нервная система включает три основных компонента: 1) нервные клетки (нейроны); 2) связанные с ними клетки глии, в частности клетки нейроглии, а также клетки, образующие неврилемму; 3) соединительная ткань. Нейроны обеспечивают проведение нервных импульсов; нейроглия выполняет опорные, защитные и трофические функции как в головном, так и в спинном мозгу, а неврилемма, состоящая преимущественно из специализированных, т.н. шванновских клеток, участвует в образовании оболочек волокон периферических нервов; соединительная ткань поддерживает и связывает воедино различные части нервной системы.
Также по теме:
ГОЛОВНОЙ МОЗГ ЧЕЛОВЕКА
Нервную систему человека подразделяют по-разному. Анатомически она состоит из центральной нервной системы (ЦНС) и периферической нервной системы (ПНС). ЦНС включает головной и спинной мозг, а ПНС, обеспечивающая связь ЦНС с различными частями тела, – черепно-мозговые и спинномозговые нервы, а также нервные узлы (ганглии) и нервные сплетения, лежащие вне спинного и головного мозга.

Нейрон.
Также по теме:
КРОВЕНОСНАЯ СИСТЕМА
Структурно-функциональной единицей нервной системы является нервная клетка – нейрон. По оценкам, в нервной системе человека более 100 млрд. нейронов. Типичный нейрон состоит из тела (т.е. ядерной части) и отростков, одного обычно неветвящегося отростка, аксона, и нескольких ветвящихся – дендритов. По аксону импульсы идут от тела клетки к мышцам, железам или другим нейронам, тогда как по дендритам они поступают в тело клетки.
В нейроне, как и в других клетках, есть ядро и ряд мельчайших структур – органелл (см. также КЛЕТКА). К ним относятся эндоплазматический ретикулум, рибосомы, тельца Ниссля (тигроид), митохондрии, комплекс Гольджи, лизосомы, филаменты (нейрофиламенты и микротрубочки).
Скорость передачи нервного импульса
Не́рвный и́мпульс, распространение по нервным волокнам возбуждения (биоэлектрического импульса) в ответ на раздражение нейронов.
Во второй половине 19 века в работах Г. Гельмгольца и Э. Геринга на нерве лягушки было показано, что биоэлектрический сигнал (ток, или потенциал действия), в отличие от электрического тока в обычном проводнике, распространяется по нервному волокну с конечной скоростью (3-120 м/сек).
Возможность распространения нервных импульсов по нервным волокнам определяется их строением, напоминающим строение электрического кабеля, где роль проводника играют аксоны, а роль изолятора — миелиновая оболочка аксона, представляющая собой мембрану шванновской клетки, намотанную на аксон в несколько слоев.
Основной компонент миелиновой оболочки — липопротеид миелин, обладающий свойствами диэлектрика. Скорость распространения нервных импульсов зависит как от диаметра нервных волокон (чем толще волокно, тем выше скорость), так и от степени их электрической изоляции, так как покрытые миелином волокна при прочих равных условиях быстрее проводят нервные импульсы. Миелиновая оболочка покрывает волокно не непрерывно по всей его длине, а образует подобие изолирующих керамических «муфт», плотно нанизанных на аксон, как на стержень электрического кабеля.
Между соседними «муфтами» из миелина остаются лишь небольшие электрически неизолированные участки, через которые ионный ток может легко вытекать из аксона в наружную среду и обратно, раздражая мембрану и вызывая генерацию потенциала действия исключительно в неизолированных участках аксона, получивших название перехватов Ранвье. Нервный импульс распространяется по миелинизированному нервному волокну скачками — от одного перехвата Ранвье до следующего, что значительно повышает скорость распространения возбуждения от клетки к клетке.
Скорость распространения нервного импульса по толстым миелинизированным волокнам (диаметром 10-20 микрон) у человека достигает 70-120 м/сек, а по самым тонким немиелинизированным нервным волокнам — на два порядка ниже (менее 2 м/сек).
Способность вырабатывать нервные импульсы — одно из основополагающих свойств нейронов.
Нервные импульсы обеспечивают быстрое проведение однотипных сигналов (потенциалов действия) по аксонам на большие расстояния и поэтому являются важнейшим средством обмена информацией как между нервными клетками, так и между нервными и другими типами клеток. Информация о силе раздражения нервной клетки кодируется и передается другим клеткам путем изменения частоты следования нервных импульсов.
Частота следования может варьировать от единиц до сотни нервных импульсов в секунду. Частотный код предполагает сложную периодику следования нервных импульсов, в том числе группирование их в «пачки» с разным числом и характером следования сигналов. Сложная пространственная и временная суммация нервных импульсов составляет основу ритмической электрической активности мозга, регистрируемой с помощью электроэнцефалограммы.
Скорость распространения нервных импульсов может быть различной: меньше 1 метра в секунду в очень тонких аксонах и около 100 метров в секунду в толстых аксонах {например, в аксонах, иннервирующих мышцы).
Распространяющийся по аксону электрический импульс, доходя до окончаний аксона на другой нервной клетке, внезапно исчезает. Чарлз Шеррингтон, заложивший основы так называемой синаптологии, назвал точки контакта окончаний аксона с другой нервной клеткой «синапсами».
Для того чтобы «перейти» через синапс, нервный импульс должен быть заново генерирован по другую сторону синапса. Еще 15 лет назад некоторые физиологи считали, что передача импульса через синапс — явление в основном (если не полностью) электрического порядка. Теперь, однако, имеются многочисленные доказательства того, что при такой передаче происходит выделение особых веществ, вызывающих регенерацию импульса. Первое убедительное доказательство того, что в синапсе действует вещество-передатчик, было получено более 40 лет назад Г. Дейлом и О. Лёви.
Как известно, центральная нервная система человека (включая, конечно, не только головной мозг, но и спинной) состоит примерно из 10 миллиардов (1010) нервных клеток. Почти все нервные клетки, за редким исключением, получают информацию непосредственно в форме импульсов (смотрите рисунок ниже) сразу от нескольких нервных клеток (нередко от сотен их) и передают ее столь же большому числу клеток.
Нервный импульс.
Также по теме:
НЕЙРОПСИХОЛОГИЯ
Если раздражение нейрона превышает определенную пороговую величину, то в точке стимуляции возникает серия химических и электрических изменений, которые распространяются по всему нейрону. Передающиеся электрические изменения называются нервным импульсом. В отличие от простого электрического разряда, который из-за сопротивления нейрона будет постепенно ослабевать и сумеет преодолеть лишь короткое расстояние, гораздо медленнее «бегущий» нервный импульс в процессе распространения постоянно восстанавливается (регенерирует).
Концентрации ионов (электрически заряженных атомов) – главным образом натрия и калия, а также органических веществ – вне нейрона и внутри него неодинаковы, поэтому нервная клетка в состоянии покоя заряжена изнутри отрицательно, а снаружи положительно; в результате на мембране клетки возникает разность потенциалов (т.н. «потенциал покоя» равен примерно –70 милливольтам). Любые изменения, которые уменьшают отрицательный заряд внутри клетки и тем самым разность потенциалов на мембране, называются деполяризацией.
Плазматическая мембрана, окружающая нейрон, – сложное образование, состоящее из липидов (жиров), белков и углеводов. Она практически непроницаема для ионов. Но часть белковых молекул мембраны формирует каналы, через которые определенные ионы могут проходить. Однако эти каналы, называемые ионными, открыты не постоянно, а, подобно воротам, могут открываться и закрываться.
При раздражении нейрона некоторые из натриевых (Na+) каналов открываются в точке стимуляции, благодаря чему ионы натрия входят внутрь клетки. Приток этих положительно заряженных ионов снижает отрицательный заряд внутренней поверхности мембраны в области канала, что приводит к деполяризации, которая сопровождается резким изменением вольтажа и разрядом – возникает т.н. «потенциал действия», т.е. нервный импульс. Затем натриевые каналы закрываются.
Во многих нейронах деполяризация вызывает также открытие калиевых (K+) каналов, вследствие чего ионы калия выходят из клетки. Потеря этих положительно заряженных ионов вновь увеличивает отрицательный заряд на внутренней поверхности мембраны. Затем калиевые каналы закрываются. Начинают работать и другие мембранные белки – т.н. калий-натриевые насосы, обеспечивающие перемещение Na+ из клетки, а K+ внутрь клетки, что, наряду с деятельностью калиевых каналов, восстанавливает исходное электрохимическое состояние (потенциал покоя) в точке стимуляции.
Электрохимические изменения в точке стимуляции вызывают деполяризацию в прилегающей точке мембраны, запуская в ней такой же цикл изменений. Этот процесс постоянно повторяется, причем в каждой новой точке, где происходит деполяризация, рождается импульс той же величины, что и в предыдущей точке. Таким образом, вместе с возобновляющимся электрохимическим циклом нервный импульс распространяется по нейрону от точки к точке.
Распространение нервных импульсов
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны
. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых
дендритами
, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется
аксоном
и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия
. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом
. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (
пресинаптическими
)
пузырьками
, в каждом из которых находятся особые соединения —
нейромедиаторы
. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом
). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Нервы, нервные волокна и ганглии.
Нерв – это пучок волокон, каждое из которых функционирует независимо от других. Волокна в нерве организованы в группы, окруженные специализированной соединительной тканью, в которой проходят сосуды, снабжающие нервные волокна питательными веществами и кислородом и удаляющие диоксид углерода и продукты распада. Нервные волокна, по которым импульсы распространяются от периферических рецепторов к ЦНС (афферентные), называют чувствительными или сенсорными. Волокна, передающие импульсы от ЦНС к мышцам или железам (эфферентные), называют двигательными или моторными. Большинство нервов смешанные и состоят как из чувствительных, так и из двигательных волокон. Ганглий (нервный узел) – это скопление тел нейронов в периферической нервной системе.
Волокна аксонов в ПНС окружены неврилеммой – оболочкой из шванновских клеток, которые располагаются вдоль аксона, как бусины на нити. Значительное число этих аксонов покрыто дополнительной оболочкой из миелина (белково-липидного комплекса); их называют миелинизированными (мякотными). Волокна же, окруженные клетками неврилеммы, но не покрытые миелиновой оболочкой, называют немиелинизированными (безмякотными). Миелинизированные волокна имеются только у позвоночных животных. Миелиновая оболочка формируется из плазматической мембраны шванновских клеток, которая накручивается на аксон, как моток ленты, образуя слой за слоем. Участок аксона, где две смежные шванновские клетки соприкасаются друг с другом, называется перехватом Ранвье. В ЦНС миелиновая оболочка нервных волокон образована особым типом глиальных клеток – олигодендроглией. Каждая из этих клеток формирует миелиновую оболочку сразу нескольких аксонов. Немиелинизированные волокна в ЦНС лишены оболочки из каких-либо специальных клеток.
Миелиновая оболочка ускоряет проведение нервных импульсов, которые «перескакивают» от одного перехвата Ранвье к другому, используя эту оболочку как связующий электрический кабель. Скорость проведения импульсов возрастает с утолщением миелиновой оболочки и колеблется от 2 м/с (по немиелинизированным волокнам) до 120 м/с (по волокнам, особенно богатым миелином). Для сравнения: скорость распространения электрического тока по металлическим проводам – от 300 до 3000 км/с.
Строение
Волокна, по которым проходит возбуждение, имеют специфическую оболочку. Грубо говоря, эта система напоминает электрический кабель. По своему составу оболочка может быть миелиновая и безмиелиновая. Самый главной составляющей миелиновой оболочки является – миелин, который играет роль диэлектрика.
Скорость прохождения импульса зависит от нескольких факторов, например, от толщины волокон, при чем оно толще, тем скорость развивается быстрее. Еще один фактором в повышении скорости проведения, является сам миелин. Но при этом он располагается не по всей поверхности, а участками, как бы нанизывается. Соответственно между этими участками есть те, которые остаются «голыми». По ним происходит утечка тока из аксона.
Аксоном называется отросток, с помощью него обеспечивается передача данных от одной клетки к остальным. Регулируется этот процесс с помощью синапса – непосредственной связи между нейронами или нейроном и клеткой. Еще существует, так называемое синаптическое пространство или щель. Когда поступает раздражительный импульс к нейрону, то в процессе реакции высвобождаются нейромедиаторы (молекулы химического состава). Они проходят через синаптическое отверстие, в итоге попадая на рецепторы нейрона или клетки, которой нужно донести данные. Для проведения нервного импульса необходимы ионы кальция, так как без этого не происходит высвобождение нейромедиатора.
Вегетативная система обеспечивается в основном безмиелиновыми тканями. По ним возбуждение распространяется постоянно и беспрерывно.
Принцип передачи основан на возникновении электрического поля, поэтому возникает потенциал, раздражающий мембрану соседнего участка и так по всему волокну.
При этом потенциал действия не передвигается, а появляется и исчезает в одном месте. Скорость передачи по таким волокнам составляет 1-2 м/с.

Cинапс.
Каждый нейрон имеет специализированную связь с мышцами, железами или другими нейронами. Зона функционального контакта двух нейронов называется синапсом. Межнейронные синапсы образуются между различными частями двух нервных клеток: между аксоном и дендритом, между аксоном и телом клетки, между дендритом и дендритом, между аксоном и аксоном. Нейрон, посылающий импульс к синапсу, называют пресинаптическим; нейрон, получающий импульс, – постсинаптическим. Синаптическое пространство имеет форму щели. Нервный импульс, распространяющийся по мембране пресинаптического нейрона, достигает синапса и стимулирует высвобождение особого вещества – нейромедиатора – в узкую синаптическую щель. Молекулы нейромедиатора диффундируют через щель и связываются с рецепторами на мембране постсинаптического нейрона. Если нейромедиатор стимулирует постсинаптический нейрон, его действие называют возбуждающим, если подавляет – тормозным. Результат суммации сотен и тысяч возбуждающих и тормозных импульсов, одновременно стекающихся к нейрону, – основной фактор, определяющий, будет ли этот постсинаптический нейрон генерировать нервный импульс в данный момент.
У ряда животных (например, у лангуста) между нейронами определенных нервов устанавливается особо тесная связь с формированием либо необычно узкого синапса, т.н. щелевого соединения, либо, если нейроны непосредственно контактируют друг с другом, плотного соединения. Нервные импульсы проходят через эти соединения не при участии нейромедиатора, а непосредственно, путем электрической передачи. Немногочисленные плотные соединения нейронов имеются и у млекопитающих, в том числе у человека.
Законы проведения
В медицине присутствуют четыре основных закона:
- Анатомо-физиологическая ценность. Проводится возбуждение только в том случае, если нет нарушения в целостности самого волокна. Если не обеспечивать единство, например, по причине ущемления, принятия наркотиков, то и проведение нервного импульса невозможно.
- Изолированное проведение раздражения. Возбуждение может передаваться вдоль нервного волокна, никаким образом, не распространяясь на соседние.
- Двустороннее проведение. Путь проведения импульса может быть только двух видов – центробежно и центростремительно. Но в действительности направление происходит в одном из вариантов.
- Бездекрементное проведение. Импульсы не утихают, иными словами, проводятся без декремента.
Регенерация.
К моменту рождения человека все его нейроны и бóльшая часть межнейронных связей уже сформированы, и в дальнейшем образуются лишь единичные новые нейроны. Когда нейрон погибает, он не заменяется новым. Однако оставшиеся могут брать на себя функции утраченной клетки, образуя новые отростки, которые формируют синапсы с теми нейронами, мышцами или железами, с которыми был связан утраченный нейрон.
Перерезанные или поврежденные волокна нейронов ПНС, окруженные неврилеммой, могут регенерировать, если тело клетки осталось сохранным. Ниже места перерезки неврилемма сохраняется в виде трубчатой структуры, и та часть аксона, которая осталась связанной с телом клетки, растет по этой трубке, пока не достигнет нервного окончания. Таким образом восстанавливается функция поврежденного нейрона. Аксоны в ЦНС, не окруженные неврилеммой, по-видимому, не способны вновь прорастать к месту прежнего окончания. Однако многие нейроны ЦНС могут давать новые короткие отростки – ответвления аксонов и дендритов, формирующие новые синапсы. См. также РЕГЕНЕРАЦИЯ.
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
ЦНС состоит из головного и спинного мозга и их защитных оболочек. Самой наружной является твердая мозговая оболочка, под ней расположена паутинная (арахноидальная), а затем мягкая мозговая оболочка, сращенная с поверхностью мозга. Между мягкой и паутинной оболочками находится подпаутинное (субарахноидальное) пространство, содержащее спинномозговую (цереброспинальную) жидкость, в которой как головной, так и спинной мозг буквально плавают. Действие выталкивающей силы жидкости приводит к тому, что, например, головной мозг взрослого человека, имеющий массу в среднем 1500 г, внутри черепа реально весит 50–100 г. Мозговые оболочки и спинномозговая жидкость играют также роль амортизаторов, смягчающих всевозможные удары и толчки, которые испытывает тело и которые могли бы привести к повреждению нервной системы.
ЦНС образована из серого и белого вещества. Серое вещество составляют тела клеток, дендриты и немиелинизированные аксоны, организованные в комплексы, которые включают бесчисленное множество синапсов и служат центрами обработки информации, обеспечивая многие функции нервной системы. Белое вещество состоит из миелинизированных и немиелинизированных аксонов, выполняющих роль проводников, передающих импульсы из одного центра в другой. В состав серого и белого вещества входят также клетки глии.
Нейроны ЦНС образуют множество цепей, которые выполняют две основные функции: обеспечивают рефлекторную деятельность, а также сложную обработку информации в высших мозговых центрах. Эти высшие центры, например зрительная зона коры (зрительная кора), получают входящую информацию, перерабатывают ее и передают ответный сигнал по аксонам.
Результат деятельности нервной системы – та или иная активность, в основе которой лежит сокращение или расслабление мышц либо секреция или прекращение секреции желез. Именно с работой мышц и желез связан любой способ нашего самовыражения.
Поступающая сенсорная информация подвергается обработке, проходя последовательность центров, связанных длинными аксонами, которые образуют специфические проводящие пути, например болевые, зрительные, слуховые. Чувствительные (восходящие) проводящие пути идут в восходящем направлении к центрам головного мозга. Двигательные (нисходящие) пути связывают головной мозг с двигательными нейронами черепно-мозговых и спинномозговых нервов.
Проводящие пути обычно организованы таким образом, что информация (например, болевая или тактильная) от правой половины тела поступает в левую часть мозга и наоборот. Это правило распространяется и на нисходящие двигательные пути: правая половина мозга управляет движениями левой половины тела, а левая половина – правой. Из этого общего правила, однако, есть несколько исключений.
Теория проведения нервного импульса
Нервный импульс имеет электрическую природу. Изучение процесса может проводиться с помощью осциллографа. К прибору подключают микроэлектрод, который вводят в аксон. Другой электрод помещают в солевой раствор, омывающий аксон.
Когда нервная клетка находится в неактивном состоянии, прибор регистрирует разность потенциалов между внешней и внутренней сторонами мембраны аксона (потенциал покоя)
. Потенциал покоя обусловлен действием калиевого-натриевого насоса, постоянно удаляющего из аксона ионы натрия, и закачивающего в аксон ионы калия (на три выводимые иона натрия приходится два вводимые иона калия). Ионов натрия снаружи аксона становится больше, чем ионов калия внутри, поэтому наружная сторона мембраны заряжена положительно относительно внутренней.
При прохождении нервного импульса возрастает проницаемость мембраны для ионов натрия, они устремляются внутрь аксона, в результате чего происходит перезарядка (деполяризация мембраны),
возникает
потенциал действия.
На пике потенциала действия проницаемость мембраны для ионов натрия начинает падать, и постепенно восстанавливается потенциал покоя (происходит
реполяризация мембраны).
Последующий участок нейрона деполяризуется, и. т. д. Таким образом, прохождение нервного импульса – это процесс прохождения волны перезарядки мембраны. Такой способ позволяет проводить быстрое незатухающее проведение нервного импульса независимо от расстояния. Скорость проведения в аксонах, изолированных шванновскими клетками, повышается в результате того, что перезарядка происходит только в области перехватов Ранвье, в результате чего прохождение импульса имеет скачкообразный (
сальтаторный)
характер.
Синапсы
Синапс – место контакта нейронов. У беспозвоночных животных синапсы, преимущественно, электрические – представляют собой тесный контакт нейронов, по которому беспрепятственно проходит электрический импульс. У позвоночных проведение нервного импульса в синапсе носит химический характер. Нервные окончания аксона нейрона, передающего информацию, имеют луковицеобразные утолщения, называемые синаптическими бляшками.
В синаптических бляшках имеются многочисленные
синаптические пузырьки
, которые содержат химическое вещество — посредник при передаче нервного импульса (
медиатор).
Мембрана синаптической бляшки называется
пресинаптической
мембраной. Мембрана дендрита нейрона, воспринимающего информацию, в области синапса называется
постсинаптической мембраной.
Мембраны разделены промежутком –
синаптической щелью
шириной около 20 нм.
При достижении нервным импульсом синаптической бляшки возникает деполяризация пресинаптической мембраны, в результате чего она становится проницаемой для ионов кальция. Ионы кальция активизируют процесс слияния синаптических пузырьков с пресинаптической мембраной и освобождение медиатора в синаптическую щель. Молекулы медиатора связываются с распознающими их рецепторами постсинаптической мембраны, что вызывает ее деполяризацию и дальнейшее прохождение нервного импульса по воспринимающему нейрону. Основными медиаторами нервной системы позвоночных являются ацетилхолин и норадреналин.
В химическом синапсе в отличие от электрического синапса:
1) нервный импульс проводится только в одном направлении;
2) возможна суммация эффектов от нескольких передающих нейронов;
3) возможно модулирование (усиление или торможение эффекта) в зависимости от типа медиатора.
Рефлекторная дуга
Простейшим актом деятельности нервной системы является рефлекс.
Рефлекс – это элементарный ответ нервной системы на раздражение. Рефлекс характеризуется:
1) быстротой;
2) автоматичностью, непроизвольностью (не контролируется сознанием);
3) стереотипностью (является одинаковым при любых условиях).
Совокупность нейронов, обеспечивающая путь нервных импульсов при рефлекторном акте, называется рефлекторной дугой.
Моносинаптический рефлекс –рефлекс, имеющий простую рефлекторную дугу, образованную двумя нейронами:
Стимул®Рецептор®Афферентный нейрон®Синапс®Эфферентный нейрон®Рабочий орган®Реакция
Примером моносинаптического рефлекса является коленный рефлекс у человека (разгибание ноги в коленном суставе при ударе в подколенную ямку).
Полисинаптический рефлекс-рефлекс, имеющий рефлекторную дугу, включающую два и более синапса. Кроме чувствительного и двигательного нейронов дуга включает вставочный нейрон:
Стимул®Рецептор®Афферентный нейрон®Синапс®Вставочный нейрон®Синапс®Эфферентный нейрон®Рабочий орган®Реакция
Примером полисинаптического рефлекса является реакция отдергивания руки при прикосновении к горячему.
Головной мозг
состоит из трех основных структур: больших полушарий, мозжечка и ствола.
Большие полушария – самая крупная часть мозга – содержат высшие нервные центры, составляющие основу сознания, интеллекта, личности, речи, понимания. В каждом из больших полушарий выделяют следующие образования: лежащие в глубине обособленные скопления (ядра) серого вещества, которые содержат многие важные центры; расположенный над ними крупный массив белого вещества; покрывающий полушария снаружи толстый слой серого вещества с многочисленными извилинами, составляющий кору головного мозга.
Мозжечок тоже состоит из расположенного в глубине серого вещества, промежуточного массива белого вещества и наружного толстого слоя серого вещества, образующего множество извилин. Мозжечок обеспечивает главным образом координацию движений.
Ствол мозга образован массой серого и белого вещества, не разделенной на слои. Ствол тесно связан с большими полушариями, мозжечком и спинным мозгом и содержит многочисленные центры чувствительных и двигательных проводящих путей. Первые две пары черепно-мозговых нервов отходят от больших полушарий, остальные же десять пар – от ствола. Ствол регулирует такие жизненно важные функции, как дыхание и кровообращение. См. также ГОЛОВНОЙ МОЗГ ЧЕЛОВЕКА.
Спинной мозг.
Находящийся внутри позвоночного столба и защищенный его костной тканью спинной мозг имеет цилиндрическую форму и покрыт тремя оболочками. На поперечном срезе серое вещество имеет форму буквы Н или бабочки. Серое вещество окружено белым веществом. Чувствительные волокна спинномозговых нервов заканчиваются в дорсальных (задних) отделах серого вещества – задних рогах (на концах Н, обращенных к спине). Тела двигательных нейронов спинномозговых нервов расположены в вентральных (передних) отделах серого вещества – передних рогах (на концах Н, удаленных от спины). В белом веществе проходят восходящие чувствительные проводящие пути, заканчивающиеся в сером веществе спинного мозга, и нисходящие двигательные пути, идущие от серого вещества. Кроме того, многие волокна в белом веществе связывают различные отделы серого вещества спинного мозга.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
ПНС обеспечивает двустороннюю связь центральных отделов нервной системы с органами и системами организма. Анатомически ПНС представлена черепно-мозговыми (черепными) и спинномозговыми нервами, а также относительно автономной энтеральной нервной системой, локализованной в стенке кишечника.
Все черепно-мозговые нервы (12 пар) разделяют на двигательные, чувствительные либо смешанные. Двигательные нервы начинаются в двигательных ядрах ствола, образованных телами самих моторных нейронов, а чувствительные нервы формируются из волокон тех нейронов, тела которых лежат в ганглиях за пределами мозга.
От спинного мозга отходит 31 пара спинномозговых нервов: 8 пар шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковая. Их обозначают в соответствии с положением позвонков, прилежащих к межпозвоночным отверстиям, из которых выходят данные нервы. Каждый спинномозговой нерв имеет передний и задний корешки, которые, сливаясь, образуют сам нерв. Задний корешок содержит чувствительные волокна; он тесно связан со спинальным ганглием (ганглием заднего корешка), состоящим из тел нейронов, аксоны которых образуют эти волокна. Передний корешок состоит из двигательных волокон, образованных нейронами, клеточные тела которых лежат в спинном мозге.
Таблица: Черепно-мозговые нервы
| ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ | |||
| Номер | Название | Функциональная характеристика | Иннервируемые структуры |
| I | Обонятельный | Специальный сенсорный (обоняние) | Обонятельный эпителий полости носа |
| II | Зрительный | Специальный сенсорный (зрение) | Палочки и колбочки сетчатки |
| III | Глазодвигательный | Моторный | Большинство наружных мышц глаза Гладкие мышцы радужной оболочки и хрусталика |
| IV | Блоковый | Моторный | Верхняя косая мышца глаза |
| V | Тройничный | Общесенсорный Моторный | Кожа лица, слизистая оболочка носа и рта Жевательные мышцы |
| VI | Отводящий | Моторный | Наружная прямая мышца глаза |
| VII | Лицевой | Моторный Висцеромоторный Специальный сенсорный | Мимическая мускулатура Слюнные железы Вкусовые рецепторы языка |
| VIII | Преддверно-улитковый | Специальный сенсорный Вестибулярный (равновесие) Слуховой (слух) | Полукружные каналы и пятна (рецепторные участки) лабиринта Слуховой орган в улитке (внутреннее ухо) |
| IX | Языкоглоточный | Моторный Висцеромоторный Висцеросенсорный | Мышцы задней стенки глотки Слюнные железы Рецепторы вкусовой и общей чувствительности в задней части полости рта |
| X | Блуждающий | Моторный Висцеромоторный Висцеросенсорный Общесенсорный |
Мышцы гортани и глотки Мышца сердца, гладкая мускулатура, железы легких, бронхов, желудка и кишечника, в том числе пищеварительные железы Рецепторы крупных кровеносных сосудов, легких, пищевода, желудка и кишечника Наружное ухо |
| XI | Добавочный | Моторный | Грудино-ключично-сосцевидная и трапециевидная мышцы |
| XII | Подъязычный | Моторный | Мышцы языка |
| Определения «висцеромоторный», «висцеросенсорный» указывают на связь соответствующего нерва с внутренними (висцеральными) органами. |
Чем является нервный импульс?

ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Вегетативная, или автономная, нервная система регулирует деятельность непроизвольных мышц, сердечной мышцы и различных желез. Ее структуры расположены как в центральной нервной системе, так и в периферической. Деятельность вегетативной нервной системы направлена на поддержание гомеостаза, т.е. относительно стабильного состояния внутренней среды организма, например постоянной температуры тела или кровяного давления, соответствующего потребностям организма.
Сигналы от ЦНС поступают к рабочим (эффекторным) органам через пары последовательно соединенных нейронов. Тела нейронов первого уровня располагаются в ЦНС, а их аксоны оканчиваются в вегетативных ганглиях, лежащих за пределами ЦНС, и здесь образуют синапсы с телами нейронов второго уровня, аксоны которых непосредственно контактируют с эффекторными органами. Первые нейроны называют преганглионарными, вторые – постганглионарными.
В той части вегетативной нервной системы, которую называют симпатической, тела преганглионарных нейронов расположены в сером веществе грудного (торакального) и поясничного (люмбального) отделов спинного мозга. Поэтому симпатическую систему называют также торако-люмбальной. Аксоны ее преганглионарных нейронов оканчиваются и образуют синапсы с постганглионарными нейронами в ганглиях, расположенных цепочкой вдоль позвоночника. Аксоны постганглионарных нейронов контактируют с эффекторными органами. Окончания постганглионарных волокон выделяют в качестве нейромедиатора норадреналин (вещество, близкое к адреналину), и потому симпатическая система определяется также как адренергическая.
Симпатическую систему дополняет парасимпатическая нервная система. Тела ее преганглинарных нейронов расположены в стволе мозга (интракраниально, т.е. внутри черепа) и крестцовом (сакральном) отделе спинного мозга. Поэтому парасимпатическую систему называют также кранио-сакральной. Аксоны преганглионарных парасимпатических нейронов оканчиваются и образуют синапсы с постганглионарными нейронами в ганглиях, расположенных вблизи рабочих органов. Окончания постганглионарных парасимпатических волокон выделяют нейромедиатор ацетилхолин, на основании чего парасимпатическую систему называют также холинергической.
Как правило, симпатическая система стимулирует те процессы, которые направлены на мобилизацию сил организма в экстремальных ситуациях или в условиях стресса. Парасимпатическая же система способствует накоплению или восстановлению энергетических ресурсов организма.
Реакции симпатической системы сопровождаются расходом энергетических ресурсов, повышением частоты и силы сердечных сокращений, возрастания кровяного давления и содержания сахара в крови, а также усилением притока крови к скелетным мышцам за счет уменьшения ее притока к внутренним органам и коже. Все эти изменения характерны для реакции «испуга, бегства или борьбы». Парасимпатическая система, наоборот, уменьшает частоту и силу сердечных сокращений, снижает кровяное давление, стимулирует пищеварительную систему.
Симпатическая и парасимпатическая системы действуют координированно, и их нельзя рассматривать как антагонистические. Они сообща поддерживают функционирование внутренних органов и тканей на уровне, соответствующем интенсивности стресса и эмоциональному состоянию человека. Обе системы функционируют непрерывно, но уровни их активности колеблются в зависимости от ситуации. См. также СЕРДЦЕ; ЭМОЦИЯ.
Скорость распространения нервных импульсов
В 1830 г. один из крупнейших физиологов XIX века Иоганн Мюллер заявил, что скорость распространения ПД измерить невозможно. По его мнению, поскольку ПД – это электрический импульс, он должен проводиться со скоростью, примерно равной скорости света (3–1010 см/с); учитывая небольшие размеры биологических объектов, даже с помощью лучших инструментов того времени измерить такую скорость было невозможно.
Спустя 15 лет один из студентов Мюллера Герман фон Гельмгольц с помощью простого и изящного эксперимента, который легко воспроизвести на студенческом лабораторном практикуме (рис. 6–8), измерил скорость распространения импульсов в нерве лягушки. В своих опытах Гельмгольц раздражал нерв в двух участках, отстоящих друг от друга на 3 см, и измерял время от момента подачи стимула до максимума мышечного сокращения. Предположим, что при раздражении дистального (расположенного ближе к мышцам) участка это время уменьшается на 1 мс. Тогда скорость распространения импульсов V
равна
| V | = | d | / | t | = | 3 см | / | 1 мс | = | · | 103 | см/с |
Рис. 6.8. Экспериментальная установка, аналогичная той, с помощью которой Гельмгольц измерил скорость распространения импульсов в нерве лягушки. Стимулирующие электроды сначала подводились к точке Ст1, а затем к точке Ст2. К мышце был подсоединен рычаг, заостренный конец которого вычерчивал кривую на закопченном листе бумаги, быстро передвигаемом в продольном направлении.
Эта величина оказалась на семь порядков меньше, чем скорость распространения электрического тока в медном проводнике или в растворе электролита. Отсюда Гельмгольц сделал совершенно правильный вывод, что проведение нервного импульса –это более сложный процесс, чем простое продольное распространение тока в нервном волокне.
Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в некоторых крупных волокнах) до нескольких сантиметров в секунду (в очень тонких аксонах). Эти различия между скоростью проведения в разных волокнах иллюстрируют табл. 6–1 и рис. 6–9.
Скорость распространения импульса в значительной степени зависит от того, как быстро участок мембраны, расположенный на определенном расстоянии от места подачи стимула, деполяризуется местными токами до порогового уровня. Чем выше постоянная длины волокна, тем дальше могут распространяться эти токи, тем быстрее происходит деполяризация мембраны впереди от места возбуждения и, следовательно, тем выше скорость распространения импульса. Влияние постоянной длины на эту скорость можно продемонстрировать, если поместить аксон в масло или в воздух. При этом на поверхности аксона остается лишь тонкая пленка солевого раствора, и постоянная длины уменьшается из–за увеличения наружного продольного сопротивления [в уравнении (6–2) –r
0]. В этих условиях скорость проведения возбуждения будет ниже чем при погружении аксона в солевой раствор.
Таблица 6–1. Классификация нервных волокон лягушки по их диаметру и скорости проведения возбуждения (Erlanger, Gasser, 1937)
| Группа волокон | Диаметр, мкм | Скорость, м/с |
| A α | 18,5 | |
| β | 14,0 | |
| γ | 11,0 | |
| B | — | 4,2 |
| C | 2,5 | 0,4 –0,5 |
Рис. 6.9. Скорость распространения возбуждения в различных группах волокон нерва лягушки. А. Экспериментальная установка для стимуляции пучка нервных волокон и регистрации возникающих при этом потенциалов. Б. Составной потенциал действия, записанный с помощью внеклеточных электродов и представляющий собой сумму потенциалов во всех возбужденных волокнах пучка. Волокна группы α имеют наибольший диаметр и характеризуются самой высокой скоростью проведения. Напротив, у волокон группы γ как диаметр, так и скорость проведения наиболее низки (см табл. 6–1). Стимуляция осуществлялась до момента начала регистрации.
В процессе эволюции живые организмы выработали два способа увеличения постоянной длины аксона и тем самым–скорости распространения импульса. Один из них (типичным примером могут быть гигантские аксоны кальмаров, членистоногих кольчатых червей, костистых рыб) – это увеличение диаметра аксона, т. е. уменьшение внутреннего продольного сопротивления [в уравнении (6–2) – r
i] Подробнее этот вопрос рассмотрен в дополнении 6–2. Гигантские аксоны развились в процессе эволюции у некоторых видов животных для того, чтобы обеспечивать быструю синхронную активацию двигательных рефлексов, например движений мантии у кальмара и рефлекса отдергивания либо избегания у некоторых членистоногих (раков, тараканов) и кольчатых червей (например, земляных).
РЕФЛЕКСЫ
Когда на рецептор сенсорного нейрона воздействует адекватный стимул, в нем возникает залп импульсов, запускающих ответное действие, именуемое рефлекторным актом (рефлексом). Рефлексы лежат в основе большинства проявлений жизнедеятельности нашего организма. Рефлекторный акт осуществляет т.н. рефлекторная дуга; этим термином обозначают путь передачи нервных импульсов от точки исходной стимуляции на теле до органа, совершающего ответное действие.
Дуга рефлекса, вызывающего сокращение скелетной мышцы, состоит по меньшей мере из двух нейронов: чувствительного, тело которого расположено в ганглии, а аксон образует синапс с нейронами спинного мозга или ствола мозга, и двигательного (нижнего, или периферического, мотонейрона), тело которого находится в сером веществе, а аксон оканчивается двигательной концевой пластинкой на скелетных мышечных волокнах.
В рефлекторную дугу между чувствительным и двигательным нейронами может включаться и третий, промежуточный, нейрон, расположенный в сером веществе. Дуги многих рефлексов содержат два и более промежуточных нейрона.
Рефлекторные действия осуществляются непроизвольно, многие из них не осознаются. Коленный рефлекс, например, вызывается постукиванием по сухожилию четырехглавой мышцы в области колена. Это двухнейронный рефлекс, его рефлекторная дуга состоит из мышечных веретен (мышечных рецепторов), чувствительного нейрона, периферического двигательного нейрона и мышцы. Другой пример – рефлекторное отдергивание руки от горячего предмета: дуга этого рефлекса включает чувствительный нейрон, один или несколько промежуточных нейронов в сером веществе спинного мозга, периферический двигательный нейрон и мышцу.
Многие рефлекторные акты имеют значительно более сложный механизм. Так называемые межсегментарные рефлексы складываются из комбинаций более простых рефлексов, в осуществлении которых принимают участие многие сегменты спинного мозга. Благодаря таким рефлексам обеспечивается, например, координация движений рук и ног при ходьбе. К сложным рефлексам, замыкающимся в головном мозге, относятся движения, связанные с поддержанием равновесия. Висцеральные рефлексы, т.е. рефлекторные реакции внутренних органов, опосредуются вегетативной нервной системой; они обеспечивают опорожнение мочевого пузыря и многие процессы в пищеварительной системе. См. также РЕФЛЕКС.
Химия проведения импульса
Процесс раздражения так же контролируется ионами, в основном калием, натрием и некоторыми органическими соединениями. Концентрация расположения этих веществ разная, клетка заряжена внутри себя отрицательно, а на поверхности положительно. Этот процесс будет называться разностью потенциалов. При колебании отрицательного заряда, например, его уменьшении провоцируется разность потенциалов и этот процесс называется деполяризацией.

Раздражение нейрона влечет за собой открытие каналов натрия в месте раздражения. Это может способствовать вхождению положительно заряженных частиц во внутрь клетки. Соответственно отрицательный заряд снижается и происходит потенциал действия или происходит нервный импульс. После этого натриевые каналы снова прикрываются.
Часто встречается, что именно ослабление поляризации способствует открытию калиевых каналов, что провоцирует высвобождению положительно заряженных ионов калия. Этим действием уменьшается отрицательный заряд на поверхности клетки.
Потенциал покоя или электрохимическое состояние восстанавливается тогда, когда в работу включаются калий-натриевые насосы, с помощью которых ионы натрия выходят из клетки, а калия заходят в нее.
В результате можно сказать – при возобновлении электрохимических процессов и происходят импульсы, стремящиеся по волокнам.
ЗАБОЛЕВАНИЯ НЕРВНОЙ СИСТЕМЫ
Поражения нервной системы возникают при органических заболеваниях или травмах головного и спинного мозга, мозговых оболочек, периферических нервов. Диагностика и лечение заболеваний и травм нервной системы составляют предмет особой отрасли медицины – неврологии. Психиатрия и клиническая психология занимаются главным образом психическими расстройствами. Сферы этих медицинских дисциплин часто перекрываются. См. отдельные заболевания нервной системы: АЛЬЦГЕЙМЕРА БОЛЕЗНЬ; ИНСУЛЬТ; МЕНИНГИТ; НЕВРИТ; ПАРАЛИЧ; ПАРКИНСОНА БОЛЕЗНЬ; ПОЛИОМИЕЛИТ; РАССЕЯННЫЙ СКЛЕРОЗ; СТОЛБНЯК; ДЕТСКИЙ ЦЕРЕБРАЛЬНЫЙ ПАРАЛИЧ; ХОРЕЯ; ЭНЦЕФАЛИТ; ЭПИЛЕПСИЯ. См. также АНАТОМИЯ СРАВНИТЕЛЬНАЯ; АНАТОМИЯ ЧЕЛОВЕКА.