Рассмотрим более подробно строение
отростков нейрона и различия между
ними. Как уже было сказано, определяющее
отличие отростков — функциональное,
т.е. направление проведения нервного
импульса: по аксону он проводится от
тела клетки, по дендриту — к телу.
Существует и ряд анатомических различий,
однако они не абсолютны и возможен ряд
исключений из них. Тем не менее, для
типичных аксонов и дендритов характерны
следующие признаки:
-
Аксон один, а дендритов несколько (хотя
существуют нейроны и с одним дендритом). -
Дендрит короче аксона. Длина дендрита
обычно не более700 мкм, а аксон может
достигать длины 1 м. -
Дендрит плавно отходит от
тела нейрона и постепенно истончается.
Аксон, отходя от тела клетки, практически
не меняет диаметр на всем своем
протяжении. Диаметр различных аксонов
колеблется от 0,3 до 16 мкм. От их толщины
зависит скорость проведения нервного
импульса — чем аксон толще, тем скорость
больше. Участок, примыкающий к телу
нейрона (аксонный холмик), имеет большую
толщину, чем остальная часть аксона. -
Дендриты ветвятся на всем
своем протяжении под острым углом,
дихотомически (вильчато), ветвление
начинается от тела клетки. Аксон обычно
ветвится только на конце, образуя
контакты (синапсы) с другими клетками.
Конечные разветвления аксона называют
терминалами. В некоторых местах от
аксонов могут отходить под прямым углом
тонкие ответвления — коллатерали. -
Дендриты (по крайней мере,
в ЦНС) не имеют миелиновой оболочки,
аксоны часто окружены миелиновой
оболочкой (о миелиновой оболочке см.
ниже).
Кроме того, иногда на веточках
дендрита есть выросты
— шипики, являющиеся
характерной структурной особенностью
дендритов, особенно в коре больших
полушарий (рис. 6). Шипик состоит из двух
частей — тела и головки, размеры и форма
которых варьируют. Шипики значительно
увеличивают постсинаптическую поверхность
дендрита. Они являются лабильными
образованиями и при различных воздействиях
(или разных функциональных состояниях)
могут менять свою конфигурацию,
дегенерировать и вновь появляться. В
результате увеличивается либо уменьшается
число синапсов, меняется эффективность
передачи в них нервного сигнала и т.д.

Рис. 6. Шипик на
дендрите нейрона и контактирующие с
ним пресинаптические окончания. Стрелками
показано направление проведения
информации
Теперь, когда мы рассмотрели
строение дендритов и аксонов, следует
несколько детальнее изучить строение
синапса. Синапс, состоящий из одного
пре- и одного постсинаптического
окончаний, называют простым. Однако
большинство синапсов в ЦНС являются
сложными. В таких синапсах один аксон
может контактировать сразу с несколькими
дендритами благодаря нескольким
мембранным выростам на его окончании.
И наоборот, один дендрит за счет своих
шипиков может контактировать с несколькими
аксонами. Еще более сложную структуру
имеют синаптические гломерулы
(клубочки)— компактные
скопления окончаний нервных отростков
разных клеток, формирующие большое
количество взаимных синапсов. Обычно
гломерулы окружены оболочкой из глиальных
клеток. Особенно характерно присутствие
гломерул в тех зонах мозга, где происходит
наиболее сложная обработка сигналов —
в коре больших полушарий и мозжечка, в
таламусе.
Итак, нейрон состоит из тела
(сомы) и отростков. Как правило, один из
отростков существенно длиннее остальных.
Такой длинный отросток называют нервным
волокном. В ЦНС это
всегда аксон; в периферической нервной
системе это может быть как аксон, так и
дендрит. По волокнам проводятся нервные
импульсы, имеющие электрическую природу,
в связи с чем, каждое волокно нуждается
в изолирующей оболочке.
По типу такой оболочки все
волокна делятся на миелиновые
(мякотные) и безмиелиновые
(безмякотные).
Безмиелиновые нервные волокна покрыты
только оболочкой, образованной телом
шванновской (нейроглиальной) клетки.
Эти волокна имеют малый диаметр и
полностью либо частично погружены во
впячивание шванновской клетки. Одна
шванновская клетка может образовывать
оболочку вокруг нескольких аксонов
разного диаметра. Такие волокна называются
волокнами кабельного типа (рис. 7). Так
как длина аксона существенно больше
размеров шванновских клеток, оболочку
аксона образуют цепочки нейроглиальных
клеток. Скорость проведения нервного
импульса по таким волокнам — 0,5-2 м/с.
Многие нервные волокна
имеют миелиновую оболочку. Она также
образуется нейроглиальными клетками.
При формировании такой оболочки
олигодендроцит (в ЦНС) или шванновская
клетка (в периферической нервной системе)
обхватывает участок нервного волокна
(рис. 8). После этого образуется вырост
в виде язычка, который закручивается
вокруг волокна, образуя мембранные слои
(цитоплазма при этом из «язычка»
выдавливается). Таким образом, миелиновая
оболочка представляет собой двойные
слои клеточной мембраны и по своему
химическому составу является липопротеидом,
т.е. соединением липидов (жироподобных
веществ) и белков. Миелиновая оболочка
осуществляет электрическую
изоляцию нервного волокна наиболее
эффективно. Нервный импульс проводится
по такому волокну быстрее,
чем по лишенному миелина (скорость
проведения может достигать
120 м/с). Миелиновая оболочка начинается
немного
отступя от тела нейрона и заканчивается
примерно в 2
мкм от синапса. Она состоит из цилиндров
длиной 1,5-2
мм,
каждый из которых образован своей
глиальной клеткой.
Цилиндры разделяют перехваты Ранвье —
не покрытые
миелином участки волокна (их длина 0,5 —
2,5 мкм), играющие большую роль в быстром
проведении нервного импульса. В перехватах
от аксона могут отходить коллатерали.
Поверх миелиновой оболочки у мякотных
волокон есть еще
наружная оболочка — неврилемма,
образованная цитоплазмой и ядром
нейроглиальных клеток.

Рис. 7. Строение
нервных волокон:
А
—
миелиновое;
Б
—
безмиелиновая;
I
—
волокно;
2 —
миелиновый
слой; 3—
ядро
шванновской клетки; 4 —
микротрубочки;
5—Нейрофиламенты;
6 —
митохондрии;
7—соединительнотканная
оболочка
Рис.
8.
Строение миелиновой оболочки (А).
Образование
миелиновой оболочки шванновской клеткой
(Б): ‘
1
—
аксон;
2 —
слои
миелиновой оболочки; 3
— перехваты
Ранвье;
4
—
ядро
шванновской клетки. Стрелкой показано
направление
продвижения
выроста цитоплазматической мембраны
Миелин
имеет белый цвет. Именно это его свойство
позволило
разделить вещество нервной системы на
серое и белое. Тела
нейронов и их короткие отростки образуют
более темное серое
вещество, а
волокна — белое
вещество.
-
Классификация
нейронов
Нейроны очень
разнообразны по форме, величине,
количеству и способу отхождения от тела
отростков, химическому строению (имеется
в виду, в первую очередь, синтез тех или
иных нейромедиаторов) и т.д. (рис. 9). Тела
самых крупных нейронов достигают в
диаметре 100 — 120 мкм (гигантские пирамиды
Беца в коре больших полушарий), самых
мелких — 4-5 мкм (зернистые клетки коры
мозжечка). Приведем основные способы
классификации нервных клеток.



Рис.
9.
Различные типы нейронов:
А
—
псевдоуниполярный
нейрон спинномозгового ганглия;
Б
—
биполярный
нейрон сетчатки; В —
мотонейрон
спинного мозга;
Г
—
пирамидная
клетка коры больших полушарий (видно,
что дендриты покрыты
шипиками); Д —
клетка
Пуркинье мозжечка; I
—
тело
клетки;
2
—
дендрит;
3 —
аксон;
4 —
коллатерали
аксона
-
Функционально
нейроны подразделяются на чувствительные
(сенсорные), вставочные (переключательные,
интернейроны)
и исполнительные (двигательные или
мотонейроны и
др.). Сенсорные
нейроны —
это нервные клетки, воспринимающие
раздражения из внешней или внутренней
среды организма.
Интернейроны
(вставочные
нейроны) обеспечивают связь
между чувствительными и исполнительными
нейронами рефлекторных дугах. Общее
направление эволюции нервной системы
связано с увеличением числа интернейронов.
Из более
чем ста миллиардов нейронов человека
более 70% составляют
вставочные нейроны.
Исполнительные
нейроны,
управляющие сокращениями поперечно —
полосатых мышечных волокон, называют
двигательными
(мотонейронами). Они
образуют нервно-мышечные синапсы.
Исполнительные нейроны, называемые
вегетативными, управляют
работой внутренних органов, включая
гладкомышечные волокна, железистые
клетки и др.
2. По количеству отростков
нейроны делятся на униполярные,
псевдоуниполярные, биполярные и
мультиполярные.Большинство
нейронов нервной системы (и почти все
нейроны в ЦНС) — это мультиполярные
нейроны (см.
рис. 9, В — Д), они имеют
один аксон и несколько дендритов.
Биполярные
нейроны (см.
рис. 9, Б) имеют один аксон и один дендрит
и характерны для
периферических отделов анализаторных
систем. Униполярных
нейронов, имеющих только один отросток,
у человека практически
нет. Из тела псевдоуниполярного
нейрона (см.
рис.9,
А) выходит один отросток, который
практически сразу делится
на две ветви. Одна из них выполняет
функцию дендрита, а
другая — аксона. Такие нейроны находятся
в чувствительных спинномозговых
и черепных ганглиях. Их дендрит
морфологически
(по строению) похож на аксон: он гораздо
длиннее аксона и часто имеет миелиновую
оболочку.
3. По форме тела и характеру
ветвления отростков выделяют звездчатые,
пирамидные, веретеновидные, корзинчатые,
зернистые
и др. нейроны.
-
По длине аксона нейроны делят
на клетки типа Гольджи I и типа Гольджи
II (эта классификация разработана
итальянским ученым К.
Гольджи). Клетки Гольджи I
имеют длинный
аксон, выходящий за пределы области, в
которой находится
тело нейрона. Это, например, пирамидные
клетки коры
больших полушарий. У клеток Гольджи II
короткий и, как
правило, очень разветвленный аксон, не
выходящий за пределы
области, в которой находится тело
нейрона. Примером
таких нейронов могут быть корзинчатые
клетки коры мозжечка. -
Каждый нейрон синтезирует
только один основной нейромедиатор.
Для того чтобы определить нервную
клетку с этой
точки зрения к названию медиатора
добавляют окончание
«-ергический». Например, ацетилхолинергический
нейрон образует ацетилхолин,
глицинергический — глицин и т.д.
Соседние файлы в предмете Анатомия и физиология
- #
01.06.201515.58 Mб544ОТ НЕЙРОНА К МОЗГУ.doc
- #
- #
- #
- #
- #
Аксон
-
Аксон (др.-греч. ἄξων «ось») — это нейрит (длинный цилиндрический отросток нервной клетки), по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.
Каждый нейрон состоит из одного аксона, тела (перикариона) и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные или мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону, а затем сгенерированный потенциал действия от начального сегмента аксона передаётся назад к дендритам. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС).
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
Источник: Википедия
Связанные понятия
Си́напс (греч. σύναψις, от συνάπτειν — соединение, связь) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём, посредством прохождения ионов из одной клетки в другую.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться один с другим, формируя биологические…
Дендрит (от греч. δένδρον (dendron) — дерево) — разветвлённый отросток нейрона, который получает информацию через химические (или электрические) синапсы от аксонов (или дендритов и сомы) других нейронов и передаёт её через электрический сигнал телу нейрона (перикариону), из которого вырастает. Термин «дендрит» ввёл в научный оборот швейцарский ученый В. Гис в 1889 году.
Миелин (в некоторых изданиях употребляется некорректная теперь форма миэлин) — вещество, образующее миелиновую оболочку нервных волокон.
Вставочный нейрон (синонимы: интернейрон, промежуточный нейрон; англ. interneuron, relay neuron, association neuron, bipolar neuron) — нейрон, связанный только с другими нейронами, в отличие от двигательных нейронов, иннервирующих мышечные волокна, и сенсорных нейронов, преобразующих стимулы из внешней среды в электрические сигналы.
Упоминания в литературе
Аксон — длинное нервное волокно, отходящее от тела некоторых нейронов и заканчивающееся терминалями (до нескольких сотен) на клетках-мишенях: нейронах, миоцитах, клетках железы и др. В зависимости от наличия или отсутствия аксона выделяют нейроны I и II типа. Область тела нейрона, которой начинается аксон, называют начальным сегментом аксон, или аксонным холмиком. Мембрана аксонного холмика генерирует потенциалы действия (нервные импульсы), распространяющиеся по аксону. Некоторые аксоны покрыты прерывистыми цепочками миелиновых оболочек (муфт) и называются миелинизированными (волокна спинномозговых нервов). Аксоны, свободные от миелина, называются немиелинизированными (волокна вегетативных нервов). Между муфтами имеются промежутки – перехваты Ранвье. В участках, лишенных миелиновой оболочки, мембрана аксона контактирует непосредственно с внеклеточной средой. В ЦНС миелиновые муфты образованы глиальными клетками, олигодендритами. Миелиновые муфты аксонов, выходящих за пределы ЦНС (периферических волокон), образованы шванновскими клетками. Одна шванновская клетка может образовывать муфты, покрывая до девяти нервных волокон. Главная функция аксонов – передача нервных импульсов от тела нейрона к терминалям, которые заканчиваются на клетках-мишенях утолщениями, синаптическими бляшками. Скорость движения информации по аксонам составляет от нескольких миллиметров в секунду в мелких немиелинизированных волокнах до 90 м/с в крупных миелинизированных. Миелиновая оболочка увеличивает скорость передачи потенциалов действия по аксону.
НЕЙРО́Н, нервная клетка; основная структурная и функциональная единица нервной системы. Нейроны обладают особыми свойствами – возбудимостью и проводимостью. Из нейронов построена нервная ткань. Как и другие клетки, нейрон имеет ядро, цитоплазму и клеточную мембрану. Ядро содержится в теле нейрона (его расширенная часть), от которого отходят отростки – короткие (дендриты) и один длинный (аксон). Длина аксона может достигать 1 м и более. Аксоны образуют тяжки, которые называются нервами (наиболее крупные – нервными стволами). Дендриты принимают сигналы из внешней среды или от другой нервной клетки и проводят возбуждение к аксону, который передаёт его в удалённые от тела нейрона области. Тела нейронов образуют серое вещество головного и спинного мозга, а дендриты и аксоны – белое мозговое вещество.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка – интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Нейрон. Основной функциональной и структурной единицей нервной системы является нейрон, строение которого показано на рис. 1.1. Нейрон состоит из клеточного тела, или сомы, коротких нервных отростков, называемых дендритами, и длинного нервного волокна – аксона. Область выхода аксона из клеточного тела нейрона называется аксонным холмиком. Хотя формально и дендриты и аксоны являются нервными волокнами, термин «нервное волокно» в основном употребляется при упоминании аксона. Дендриты передают нервный импульс к телу нейрона, тогда как аксон направляет импульс из тела нейрона на другие нервные клетки или на мышечные волокна.
Связанные понятия (продолжение)
Клетки Пуркинье (англ. Purkinje cells) — крупные нервные клетки коры мозжечка. Своё название клетки получили в честь их первооткрывателя, чешского врача и физиолога Яна Эвангелисты Пуркинье.
Пирамидальные нейроны, или пирамидные нейроны, — основные возбудительные нейроны мозга млекопитающих. Также обнаруживаются у рыб, птиц, рептилий. Напоминают по форме пирамиду, из которой вверх ведёт большой апикальный дендрит; имеют один аксон, идущий вниз, и множество базальных дендритов. Впервые были исследованы Рамон-и-Кахалем. Отмечены в таких структурах, как кора мозга, гиппокамп, миндалевидное тело (амигдала), но отсутствуют в обонятельной луковице, стриатуме, среднем мозге, ромбовидном мозге…
Подробнее: Пирамидальный нейрон
Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов. Термин ввёл в 1846 году Рудольф Вирхов.
Перикарион (др.-греч. περι- — приставка со значением «около, вокруг, кругом» + κάρυον — «орех») — сома (тело) нейрона, может иметь различную величину и форму. На цитолемме перикариона образуются многочисленные синаптические контакты с отростками других нейронов.
Нервная ткань — ткань эктодермального происхождения, представляет собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию.
Мотонейро́н (от лат. motor — приводящий в движение и нейрон; двигательный нейро́н) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса.
Шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Астроцит (лат. astrocytus; от греч. astron — звезда; и kýtos, здесь — клетка) — тип нейроглиальной клетки звездчатой формы с многочисленными отростками. Совокупность астроцитов называется астроглией.
Гранулярные клетки — несколько разновидностей мелких нейронов мозга. Название «гранулярная клетка» («зернистая клетка», «клетка-зерно») используется анатомами для нескольких разных типов нейронов, единственной общей особенностью которых является крайне малый размер тел этих клеток.
Подробнее: Гранулярная клетка
Гипотала́мус (лат. hypothalamus, от греч. ὑπό — «под» и θάλαμος — «комната, камера, отсек, таламус») — небольшая область в промежуточном мозге, включающая в себя большое число групп клеток (свыше 30 ядер), которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма. Гипоталамус связан нервными путями практически со всеми отделами центральной нервной системы, включая кору, гиппокамп, миндалину, мозжечок, ствол мозга и спинной мозг. Вместе с гипофизом гипоталамус образует гипоталамо-гипофизарную…
Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие.
Корзинчатые нейроны — тормозные ГАМК-эргические вставочные нейроны молекулярного слоя мозжечка. Длинные аксоны корзинчатых нейронов образуют корзиноподобные синапсы с телами клеток Пуркинье. Корзинчатые нейроны многополярны, их дендриты свободно ветвятся.
Подробнее: Корзинчатый нейрон
Тала́мус, иногда — зри́тельные бугры (лат. Thalamus; от др.-греч. θάλαμος — «камера, комната, отсек») — отдел головного мозга, представляющий собой большую массу серого вещества, расположенную в верхней части таламической области промежуточного мозга хордовых животных, в том числе и человека. Впервые описан древнеримским врачом и анатомом Галеном. Таламус — это парная структура, состоящая из двух половинок, симметричных относительно межполушарной плоскости. Таламус находится глубже структур большого…
Полоса́тое те́ло (лат. corpus striatum) — анатомическая структура конечного мозга, относящаяся к базальным ядрам полушарий головного мозга.
Гиппока́мп (от др.-греч. ἱππόκαμπος — морской конёк) — часть лимбической системы головного мозга (обонятельного мозга). Участвует в механизмах формирования эмоций, консолидации памяти (то есть перехода кратковременной памяти в долговременную). Генерирует тета-ритм при удержании внимания.
Латеральное коленчатое тело (наружное коленчатое тело, ЛКТ) — легко распознаваемая структура мозга, которая помещается на нижней латеральной стороне подушки таламуса в виде достаточно большого плоского бугорка. В ЛКТ приматов и человека морфологически определено шесть слоев: 1 и 2 — слои крупных клеток, 3-6 — слои мелких клеток. Слои 1, 4 и 6 получают афференты от контрлатерального (расположенного в противоположном по отношению к ЛКТ полушарии) глаза, а слои 2, 3 и 5 — от ипсилатерального (расположенного…
Зубчатая извилина (лат. gyrus dentatus) или зубчатая фасция гиппокампа (лат. fascia dentata hippocampi) — зазубренная извилина, расположенная в глубине борозды гиппокампа и переходящая в ленточную извилину. В некоторых классификациях она вместе с аммоновым рогом считается частью самого гиппокампа, однако большинство авторов относят её к гиппокамповой формации. В её структуре выделяют три слоя: полиморфный хилус, гранулярный слой и молекулярный слой, который непрерывно переходит в молекулярный слой…
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в виде кратковременного изменения мембранного потенциала на небольшом участке возбудимой клетки (нейрона или кардиомиоцита), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к внутренней поверхности мембраны, в то время, как в покое она заряжена положительно. Потенциал действия является физиологической основой нервного импульса.
Обонятельная луковица — часть обонятельного мозга, парное образование, состоящее из тел вторых нейронов биполярного типа обонятельного анализатора. Располагается во внутричерепной полости между лобной долей сверху и решётчатой пластинкой решётчатой кости снизу, через отверстие которой в неё поступают нервные волокна обонятельной области носа, а сзади продолжается в обонятельный тракт.
Реце́птор — объединение из терминалей (нервных окончаний) дендритов чувствительных нейронов, глии, специализированных образований межклеточного вещества и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражитель) в нервный импульс. В некоторых рецепторах (например, вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками эпителиального происхождения или…
Ганглий (др.-греч. γάγγλιον — узел), или нервный узел — скопление нервных клеток, состоящее из тел, дендритов и аксонов нервных клеток и глиальных клеток. Обычно ганглий имеет также оболочку из соединительной ткани. Имеются у многих беспозвоночных и всех позвоночных животных. Часто соединяются между собой, образуя различные структуры (нервные сплетения, нервные цепочки и т. п.).
Голубое пятно (голубоватое место, голубоватое пятно, синее пятно/место, лат. locus coeruleus) — ядро, расположенное в стволе мозга на уровне моста (участок голубоватого цвета в верхнелатеральной части ромбовидной ямки ствола головного мозга кнаружи от верхней ямки), часть ретикулярной формации. Система его проекций очень широка — аксоны восходят к верхним слоям коры больших полушарий, гиппокампу, миндалине, перегородке, полосатому телу, коре мозжечка. Нисходящие проекции идут в спинной мозг к симпатическим…
Афферентация (от лат. afferens — «приносящий») — постоянный поток нервных импульсов, поступающих в центральную нервную систему от органов чувств, воспринимающих информацию как от внешних раздражителей (экстерорецепция), так и от внутренних органов (интерорецепция). Находится в прямой зависимости от количества и силы воздействующих раздражителей, а также от состояния — активности или пассивности — индивида.
Белое вещество (лат. substantia alba) — компонент центральной нервной системы позвоночных животных и человека, состоящий главным образом из пучков аксонов, покрытых миелином. Противопоставляется серому веществу мозга, состоящему из клеточных тел нейронов. Цветовая дифференциация белого и серого вещества нервной ткани обусловлена белым цветом миелина.
Нервно-мышечный синапс (также нейромышечный, либо мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в этом синапсе является ацетилхолин.
Ретикулярная формация (лат. reticulum — сеточка, formatio — образование) — это образование, тянущееся вдоль всей оси ствола головного мозга. Своим названием оно обязано сетчатой структуре, образуемой его нервными клетками с очень сложными связями. Формация состоит из ретикулярных ядер и большой сети нейронов с разветвлёнными аксонами и дендритами, представляющих единый комплекс, который осуществляет активацию коры головного мозга и контролирует рефлекторную деятельность спинного мозга. Эта сеть нейронов…
Средний мозг, или мезэнцефалон (англ. Midbrain, лат. Mesencephalon; термин «мезэнцефалон» происходит от др.-греч. μέσος — «месос» — средний, и ἐγκέφᾰλος — «энкефалос» — буквально «находящийся внутри головы», то есть головной мозг) — это отдел головного мозга хордовых животных, развивающийся из среднего из трёх первичных мозговых пузырей эмбриона. Этот отдел мозга ответствен за осуществление многих важных физиологических функций, таких, как зрение, слух, контроль движений, регуляция циклов сна и бодрствования…
Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
Мозгово́й ствол, или ствол головного мозга, — традиционно выделяющийся отдел третьего мозга, представляющий собой протяжённое образование, продолжающее спинной мозг.
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг…
Конечный мозг (лат. telencephalon) — самый передний отдел головного мозга. Состоит из двух полушарий большого мозга (покрытых корой), мозолистого тела, полосатого тела и обонятельного мозга. Является наиболее крупным отделом головного мозга. Это также самая развитая структура, покрывающая собой все отделы головного мозга.
Дендритный шипик — мембранный вырост на поверхности дендрита, способный образовать синаптическое соединение. Шипики обычно имеют тонкую дендритную шейку, оканчивающуюся шарообразной дендритной головкой. Дендритные шипики обнаруживаются на дендритах большинства основных типов нейронов мозга. В создании шипиков участвует белок калирин.
Эпиталамус (буквально «надталамус») — это самая дорсальная (верхняя) задняя часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо эпиталамуса, входят также таламус, субталамус и метаталамус, но не входят гипоталамус и гипофиз, причисляемые к гипоталамической области. Эпиталамус возвышается над таламусом. В число его структур входят поводок эпиталамуса, также называемый поводком мозга, треугольник поводка, спайка поводков, подспаечный орган…
Передний мозг, или прозэнцефалон (лат. prosencephalon, англ. forebrain) — это отдел головного мозга хордовых животных, выделяемый на основании особенностей его эмбрионального развития у этих видов животных. Передний мозг (прозэнцефалон) является одним из трёх первичных мозговых пузырей, образующихся в первичной нервной трубке вскоре после нейруляции и образования нейромер, на так называемой трёхпузырьковой стадии развития ЦНС эмбриона. Двумя другими первичными мозговыми пузырями являются средний…
Миоци́ты, или мы́шечные клетки — особый тип клеток, составляющий основную часть мышечной ткани. Миоциты представляют собой длинные, вытянутые клетки, развивающиеся из клеток-предшественников — миобластов. Существует несколько типов миоцитов: миоциты сердечной мышцы (кардиомиоциты), скелетной и гладкой мускулатуры. Каждый из этих типов обладает особыми свойствами. Например, кардиомиоциты, помимо прочего, генерируют электрические импульсы, задающие сердечный ритм.
Головно́й мозг (лат. cerebrum, др.-греч. ἐγκέφαλος) — главный орган центральной нервной системы подавляющего большинства хордовых, её головной конец; у позвоночных находится внутри черепа. В анатомической номенклатуре позвоночных, в том числе человека, мозг в целом чаще всего обозначается как encephalon — латинизированная форма греческого слова; изначально латинское cerebrum стало синонимом большого мозга (telencephalon).
Спинно́й мозг (лат. medulla spinalis) — орган центральной нервной системы позвоночных, расположенный в позвоночном канале. Принято считать, что граница между спинным и головным мозгом проходит на уровне перекреста пирамидных волокон (хотя эта граница весьма условна). Внутри спинного мозга имеется полость, называемая центральным каналом (лат. canalis centralis). Спинной мозг защищён мягкой, паутинной и твёрдой мозговой оболочкой. Пространства между оболочками и спинномозговым каналом заполнены спинномозговой…
Бледный шар (лат. globus pallidus s. pallidum) — парная структура переднего мозга, относящаяся к базальным ядрам, часть чечевицеобразного ядра, вентромедиальная часть полосатого тела. Подразделяется на латеральную и медиальную части.
Промежуточный мозг, или диэнцефалон (лат. Diencephalon, англ. Diencephalon; термин «диэнцефалон» происходит от др.-греч. διά — «диа-», обозначающее «через», «между», и ἐγκέφαλος — «энкефалос», буквально «находящийся внутри головы», то есть головной мозг) — отдел головного мозга хордовых животных, который образуется в процессе эмбрионального развития из задней части зародышевого переднего мозга (прозэнцефалона). На пятипузырьковой стадии из задней части зародышевого переднего мозга (прозэнцефалона…
Олигодендроциты, или олигодендроглия — это вид нейроглии, открытый Пио дель Рио-Ортегой (1928 год). Олигодендроциты есть только в центральной нервной системе, которая у позвоночных включает в себя головной мозг и спинной мозг.
Синаптогенез — процесс формирования синапсов между нейронами в нервной системе. Синаптогенез происходит на протяжении всей жизни здорового человека, а взрыв формирования синапсов, т. н. избыточный синаптогенез (exuberant synaptogenesis), наблюдается на ранних стадиях развития головного мозга. Синаптогенез особенно важен в ходе критического периода развития особи (в биологии развития, такого периода, когда нервная система особенно чувствительна к экзогенным стимулам), когда имеет место быть интенсивное…
Хвостатое ядро (лат. nucleus caudatus) — парная структура головного мозга, относящаяся к стриатуму. Расположена спереди от таламуса, от которого (на горизонтальном срезе) его отделяет белая полоска вещества — внутренняя капсула. Передний отдел хвостатого ядра утолщён и образует головку, caput nuclei caudati, которая составляет латеральную стенку переднего рога бокового желудочка. Головка хвостатого ядра примыкает внизу к переднему продырявленному веществу, в этом месте головка соединяется с чечевицеобразным…
Продолговатый мозг (лат. myelencephalon, medulla oblongata), или луковица головного мозга (лат. bulbus cerebri), — задний отдел головного мозга, непосредственное продолжение спинного мозга. Происходит из ромбовидного мозга и входит в ствол головного мозга. Регулирует такие основные процессы жизнедеятельности, как дыхание и кровообращение, поэтому в случае повреждения продолговатого мозга мгновенно наступает смерть.
Химический синапс — особый тип межклеточного контакта между нейроном и клеткой-мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество.
Электри́ческий си́напс (англ. electrical synapse) — место высокоспециализированных контактов (щелевых контактов) между нейронами, где происходит прямое перетекание электрических токов от одного нейрона к другому. В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм, в то время как в химическом синапсе расстояние между двумя нейронами составляет от 20 до 40 нм. У многих животных в нервной системе имеются как химические, так и электрические синапсы. По сравнению с химическими…
Упоминания в литературе (продолжение)
Основная единица нервной системы — нервная клетка (нейрон). Нейроны покрыты глиальными клетками, доставляющими им питание и удерживающими их на месте. Каждая нервная клетка состоит из тела, дендрита и аксона. В периферической нервной системе аксоны собраны в пучки – нервы. Бо́льшая часть аксонов покрыта миелиновой оболочкой, которая увеличивает скорость передачи информации в нервной системе, а также препятствует передаче нервных импульсов близлежащим клеткам. Современные исследователи опровергли бытовавшее ранее мнение о том, что «нервные клетки не восстанавливаются». Доказано, что клетки в зонах мозга, связанных с памятью и процессом обучения, продолжают развиваться и делиться в течение жизни человека.
Нервные клетки имеют несколько отростков – чувствительных древовидно ветвящихся дендритов, которые проводят к телу нейронов возбуждение, возникающее на их чувствительных нервных окончаниях, расположенных в органах, и одних двигательный аксон, по которому нервный импульс передается от нейрона к рабочему органу или другому нейрону. Нейроны вступают друг с другом в контакт с помощью окончаний отростков, образуя рефлекторные цепи, по которым передаются (распространяются) нервные импульсы.
От нервных центров по основному отростку (аксону) идут сигналы к каждой клетке и каждому органу тела, заставляя их путем электрической стимуляции выполнять определенную функцию. Нервные центры состоят из сотен и даже тысяч нервных клеток. Соответственно, существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Основной ее элемент – нейрон: микроскопически малая нервная клетка с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Вспомогательные клетки нейроглии (астроциты) не только создают физическую опору для нейронов, но вместе с сосудами обеспечивают потребности нервной ткани в кислороде и необходимых для жизни веществах, включая аминокислоты, липиды, гликопротеиды. Тело нервной клетки имеет микроскопические размеры, но длина аксона может достигать одного метра! Отростки нейронов, как правило, укрыты миелиновой оболочкой, которая обеспечивает стабильность обмена веществ в длинных нервных проводниках и высокую скорость передачи возбуждения.
Нейрон представляет собой микроскопически малую нервную клетку с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Главная структурная и функциональная единица нервной системы – нейрон, представляющий собой клетку с отростками – длинным аксоном и короткими дендритами. Нейроны соединяются между собой синапсами, образуя нейронные цепи, приводимые в действие рефлекторно: в ответ на раздражение, поступаемое из внешней или внутренней среды, возбуждение из нервных окончаний передается по центростремительным волокнам в головной и спинной мозг, оттуда импульсы по центробежным волокнам поступают в различные органы, а по двигательным – к мышцам.
Аксон – вырост цитоплазмы, приспособленный для проведения информации, которая собирается ден-дритами и перерабатывается в нейроне. Аксон дендритной клетки имеет постоянный диаметр и покрыт миелиновой оболочкой, которая образована из глии, у аксона разветвленные окончания, в которых находятся митохондрии и секреторные образования.
Два слова о строении нейрона. У него есть центральная толстенькая часть – «тело», в котором находится клеточное ядро с генами. От тела отходят два вида отростков: «входные» (дендриты) и «выходные» (аксоны). Дендритов обычно много, а аксон, как правило, один, но на конце он может ветвиться.
При поражении центров коры головного мозга очень важна локализация, ведь повреждения происходят на микроскопическом уровне. Головной мозг состоит из нейронов и глиальных клеток, которые соединены между собой сложными проводимыми путями – аксонами и дендритами. По ним передаются нервные импульсы. Все мы еще со школьного курса биологии знаем, что в головном мозге есть серое и белое вещества. Серым веществом является скопление нейронов и коротких проводимых путей между ними. А вот белое вещество составляют длинные, сложные проводимые пути – нервные волокна, которые покрыты особой миелиновой оболочкой – она необходима для нормального прохождения импульсов и подобна изоляционной ленте на электрическом проводе.
В состоянии относительного физиологического покоя синапс находится в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается и ее содержимое в виде одного кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
Таким образом, нейрофизиологические и психофизиологические механизмы tDCS и ТКМП достаточно многообразны, даже при приложении стандартизированных параметров тока и площади электродов (при tDCS). Нам представляется маловероятным сведение механизмов поляризаций к делоляризации или гиперполяризации мембраны нейронов. Если для пирамидных нейронов с их пространственной ориентацией в коре можно себе представить, что, например, при анодной поляризации входящий (гиперполяризующий) ток протекает преимущественно через дендрита, а выходящий (деполяризующий) ток – через начальный сегмент аксона (где генерируются спайки), вызывая его деполяризацию и увеличение возбудимости моторной коры, то это допущение плохо приложимо к другим областям коры, где ориентация нейронов не столь очевидна. Кроме того, возбуждающие или тормозные потенциалы длятся не более 10 мсек. Пассивная деполяризация мембраны еще короче. Установленные эффекты tDCS длятся десятки минут, а то и дольше. Следовательно, де– или гиперполяризация, если и играют существенную роль в возникновении эффектов, то она сводится к пусковой роли, запускающей каскад синаптических и метаболических реакций которые и лежат в основе более длительных эффектов поляризаций.
Надо сказать, что нейрон устроен очень интересно: у него, как у всякого компьютера, есть корпус – тело. И все же славится нейрон не этим, главное в нем – это его отростки. Весь нейрон чем-то напоминает дерево, которое имеет развитую крону и ствол. Крону у дерева образуют ветки, а у нейрона это короткие отростки, они называются дендритами. У дерева ствол, а у нейрона – длинный отросток, аксон. А теперь о месте соединения нервных клеток – это синапс. Если два компьютера соединить шнуром, то контакт компьютера и шнура будет как раз тем самым синапсом в нервной системе.
Активность генов в нейронах основана на свойствах молекул ДНК, т. е. активность вызвана причинно-следственными связями, которые определяют особенности контактов молекул ДНК с окружающими атомами и молекулами. В свою очередь, активность генов является причиной всех событий клеточного метаболизма и, значит, обмена веществ между нейронами и с окружающей средой. Все процессы в сообществе нервных клеток (клеточном сообществе – КС) – рождение клеток, их возбуждение и торможение, функциональная специализация и формирование в них устойчивых молекулярных структур, синтез в клетках нейромедиаторов и образование у них аксонов, дендритов и шипиков, образование и разрыв межклеточных связей, распространение и прекращение возбуждений в сообществе, гибель клеток – это детерминистские процессы. Так у нейронов в течение суток изменяются параметры метаболизма: количество и соотношение синтезируемых и разлагающихся веществ, в т. ч. количество синтезируемых нейромедиаторов и гормонов. У нейронов, как и у всех организмов, имеются суточные ритмы. В соответствии с этими ритмами в продолжение суток изменяются состав и количество клеток, возбуждающихся от воздействий окружающей среды и следов таких воздействий, результаты конкуренции клеток и узлов за прекращение своего возбуждения, используемые возбуждениями маршруты связей между клетками, конфигурация формирующихся способов прекращения возбуждений, распространяющихся из одних и тех же узлов.
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь. Коллатерали, идущие от задних корешков шейных сегментов, входят в состав клиновидного пучка, идут вверх к его ядру и к дополнительному клиновидному ядру. Его аксоны соединяются с мозжечком. Третья группа коллатеральных афферентных волокон заканчивается в задних рогах спинного мозга. Там расположены вторые нейроны, чьи аксоны образуют передний спино-мозжечковый путь.
Соответственно существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Двигательная единица – это отдельный мотонейрон и мышечные волокна, которые он иннервирует. Таким образом, нейрон определяет, являются ли волокна медленно или быстро сокращающимися. Мотонейрон в МС двигательной единице имеет небольшое клеточное тело и иннервирует группу из 10-180 мышечных волокон. У мотонейрона в БС двигательной единице большое клеточное тело и больше аксонов, и он иннервирует от 300 до 800 мышечных волокон. Отсюда следует, что каждый МС-мотонейрон в состоянии активировать значительно меньшее количество мышечных волокон в противоположность БС-мотонейрону. При этом необходимо отметить, что сила, производимая отдельными МС- и БС-волокнами по величине отличается незначительно. МС- и БС-волокна имеют разные функции во время физической активности. МС-волокнам присущ высокий уровень аэробной выносливости, они эффективны в производстве АТФ на основе окисления углеводов и жиров и более приспособлены к выполнению длительной работы невысокой интенсивности. Быстро сокращающиеся мышечные волокна приспособлены к анаэробной деятельности (без кислорода), и при их работе АТФ образуется благодаря анаэробным реакциям. Б С двигательные единицы производят большую силу, однако легко устают ввиду ограниченной выносливости и используются главным образом при выполнении кратковременной работы высокой интенсивности.
• длинный отросток, отходящий от тела клетки, который тянется на большое расстояние – до 1,5–1,7 м. Он составляет основной, или осевой, отросток нервной клетки. Его называют аксоном (в переводе с латыни axis — ось, основание, основной).
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь.
Формирование мышечной ткани начинается на 4-6-й неделе внутриутробного развития. В это время формируются первичные мышечные волокна. Несколько позже в мышцы прорастают аксоны мотонейронов спинного мозга. С этой стадии начинается синхронное формирование нервно-мышечного аппарата, причем определяющее значение имеет развитие нервных элементов, происходящее на 6-7-м месяцах внутриутробного развития. К моменту рождения примерно половина мышечных волокон уже прошла стадию первичной дифференцировки, и уже определены как «белые» или «красные». Дифференцировочные процессы усиливаются в возрасте от 1 до 2 лет, а затем на стадии полового созревания.
Нейросекреторные клетки, как и обычные нервные клетки, воспринимают сигналы, поступающие к ним от других отделов нервной системы, но далее передают полученную информацию уже гуморальным путем (не по аксонам, а по сосудам) – посредством нейрогормонов. Таким образом, совмещая свойства нервных и эндокринных клеток, нейросекреторные клетки объединяют нервные и эндокринные регуляторные механизмы в единую нейроэндокринную систему. Этим обеспечивается, в частности, способность организма адаптироваться к изменяющимся условиям внешней среды. Объединение нервных эндокринных механизмов регуляции осуществляется на уровне гипоталамуса и гипофиза.
Гидроцефалический (интерстициальный) отек. Еще одной формой отека головного мозга, сопровождающейся увеличением объема интерстициального пространства, является гидроцефалический отек, обусловленный блокадой путей, соединяющих интерстициальное пространство головного мозга с макроскопическими ликворосодержащими пространствами. Для клинициста эта форма отека имеет практическое значение. Так, у больных с острой гидроцефалией в начале происходит увеличение объема интерстициальной жидкости в перивентрикулярных отделах. Узкие в норме пространства между глиальными клетками и аксонами расширяются. Астроциты набухают, атрофируются и погибают. У больных с хронической гидроцефалией деструкция аксонов, разрушение миелина, фагоцитоз липидов микроглии являются характерными гистологическими признаками. Кроме стаза внеклеточной жидкости, причиной отека у этих больных может быть и обратный ток спинномозговой жидкости из желудочков мозга. Так же, как и при любой иной форме отека мозга, в зоне отек; снижается регионарный мозговой кровоток. По-видимому, часть функциональных расстройств в ЦНС, наблюдаемых у больных с гидроцефалией, обусловлена снижением регионарного кровотока в зоне отека.
Эти два гормона относятся к особой группе, поскольку, синтезируясь в гипоталамусе, транспортируются по аксонам (отросткам нейронов) в задний гипофиз и там выделяются в системный кровоток.
Между концами может остаться небольшой диастаз, но он не должен превышать 1 мм. Свободный промежуток между концами нерва заполнит гематома, а в дальнейшем образуется соединительно—тканная прослойка, через эту гематому и соединительную ткань будут прорастать тяжи швашювских клеток и вновь сформированные аксоны.
| Аксон | |
|---|---|

Аксон многополярного нейрона |
|
| Идентификаторы | |
| MeSH | D001369 |
| Анатомическая терминология
[редактировать в Викиданных ] |
An аксон (от греч. ἄξων áxōn, ось), или нервное волокно (или же нерв волокно: видеть орфографические различия ), представляет собой длинный тонкий выступ нервной клетки, или нейрон, у позвоночных, который обычно проводит электрические импульсы, известные как потенциалы действия подальше от тело нервной клетки. Функция аксона — передавать информацию различным нейронам, мышцам и железам. В определенных сенсорные нейроны (псевдоуниполярные нейроны ), например аксоны прикосновения и тепла, аксоны называются афферентные нервные волокна и электрический импульс проходит по ним от периферия к телу клетки и от тела клетки к спинному мозгу по другой ветви того же аксона. Дисфункция аксонов является причиной многих наследственных и приобретенных неврологических расстройств, которые могут поражать как периферические, так и центральные нейроны. Нервные волокна классифицированный на три типа — нервные волокна группы А, нервные волокна группы B, и нервные волокна группы C. Группы A и B являются миелинизированный, и группа C немиелинизированы. Эти группы включают как сенсорные волокна, так и двигательные волокна. Другая классификация группирует только сенсорные волокна как Тип I, Тип II, Тип III и Тип IV.
Аксон — это один из двух типов цитоплазматический выступы из тела клетки нейрона; другой тип — это дендрит. Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно имеют постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты получают сигналы, тогда как аксоны передают их). Некоторые типы нейронов не имеют аксона и передают сигналы от своих дендритов. У некоторых видов аксоны могут исходить из дендритов, известных как дендриты, несущие аксоны.[1] Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга.[2]
Аксоны покрыты мембраной, известной как аксолемма; цитоплазма аксона называется аксоплазма. Большинство аксонов разветвляются, в некоторых случаях очень обильно. Концевые ветви аксона называются телодендрия. Распухший конец телодендрона известен как терминал аксона который присоединяется к дендрону или телу клетки другого нейрона, образуя синаптическое соединение. Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсы. В некоторых случаях аксон одного нейрона может образовывать синапс с дендритами того же нейрона, что приводит к autapse. В синапсе мембрана аксона близко примыкает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Некоторые синаптические соединения появляются вдоль аксона по мере его расширения — они называются мимоходом («попутно») синапсов и может быть сотнями или даже тысячами вдоль одного аксона.[3] Другие синапсы выглядят как терминалы на концах аксональных ветвей.
Отдельный аксон со всеми его ветвями, взятыми вместе, может иннервировать несколько частей мозга и генерируют тысячи синаптических окончаний. Связка аксонов делает нервный тракт в Центральная нервная система,[4] и пучок в периферическая нервная система. В плацентарные млекопитающие самый большой белое вещество тракт в головном мозге мозолистое тело, образованный примерно из 200 миллионов аксонов в человеческий мозг.[4]
Анатомия

Типичный миелинизированный аксон

Аксоны — это основные линии передачи нервная система, и как пучки они образуют нервы. Некоторые аксоны могут достигать одного метра и более, а другие — всего лишь один миллиметр. Самые длинные аксоны в человеческом теле — аксоны седалищный нерв, которые идут от основания спинной мозг к большому пальцу каждой стопы. Диаметр аксонов также варьируется. Большинство отдельных аксонов микроскопические в диаметре (обычно около одного микрометр (мкм) в поперечнике). Самые большие аксоны млекопитающих могут достигать в диаметре до 20 мкм. В гигантский аксон кальмара, который специализируется на очень быстром проведении сигналов, близок к 1 миллиметр в диаметре, размером с грифель карандаша. Количество аксональных телодендрий (разветвленных структур на конце аксона) также может отличаться от одного нервного волокна к другому. Аксоны в Центральная нервная система (ЦНС) обычно показывают несколько телодендрий с множеством синаптических конечных точек. Для сравнения: гранулярная клетка мозжечка аксон характеризуется одним Т-образным узлом ветви, от которого два параллельные волокна продлевать. Продуманное ветвление позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
Есть два типа аксонов в нервная система: миелинизированный и немиелинизированный аксоны.[5] Миелин представляет собой слой изолирующего жирового вещества, который образован двумя типами глиальные клетки Клетки Шванна и олигодендроциты. в периферическая нервная система Шванновские клетки образуют миелиновую оболочку миелинизированного аксона. в Центральная нервная система олигодендроциты образуют изолирующий миелин. Вдоль миелинизированных нервных волокон пробелы в миелиновой оболочке, известные как узлы Ранвье происходят через равные промежутки времени. Миелинизация обеспечивает особенно быстрый режим распространения электрических импульсов, называемый скачкообразное проведение.
Миелинизированные аксоны из корковые нейроны образуют основную часть нервной ткани, называемой белое вещество в мозгу. Миелин придает белому цвету ткань в отличие от серое вещество коры головного мозга, содержащей тела нейрональных клеток. Аналогичное расположение можно увидеть на мозжечок. Связки миелинизированных аксонов составляют нервные пути в ЦНС. Там, где эти тракты пересекают среднюю линию мозга и соединяют противоположные области, они называются комиссур. Самый крупный из них — мозолистое тело что соединяет два полушария головного мозга, а это около 20 миллионов аксонов.[4]
Видно, что структура нейрона состоит из двух отдельных функциональных областей или компартментов — тела клетки вместе с дендритами в качестве одной области и аксональной области в качестве другой.
Аксональная область
Аксональная область или компартмент включает бугорок аксона, начальный сегмент, остальную часть аксона и телодендрии аксона, а также терминалы аксона. Он также включает миелиновую оболочку. В Тела Nissl которые продуцируют нейрональные белки, отсутствуют в аксональной области.[3] Белки, необходимые для роста аксона и удаления отходов жизнедеятельности, нуждаются в транспортном каркасе. Этот аксональный транспорт обеспечивается в аксоплазме за счет расположения микротрубочки и промежуточные нити известный как нейрофиламенты.
Аксонный бугорок

Деталь, показывающая микротрубочки на бугорке аксона и начальном сегменте.
В аксональный бугорок это область, образованная клеточным телом нейрона по мере того, как она расширяется, чтобы стать аксоном. Он предшествует начальному сегменту. Полученные потенциалы действия, которые подведены в нейроне передаются на бугорок аксона для генерации потенциала действия из начального сегмента.
Начальный сегмент
В аксональный начальный сегмент (AIS) представляет собой структурно и функционально отдельный микродомен аксона.[6][7] Одна из функций начального сегмента — отделить основную часть аксона от остальной части нейрона; другая функция — помочь инициировать потенциалы действия.[8] Обе эти функции поддерживают нейрон полярность ячейки, в котором дендриты (а в некоторых случаях сома ) нейрона получают входные сигналы в базальной области, а в апикальной области аксон нейрона обеспечивает выходные сигналы.[9]
Начальный сегмент аксона немиелинизирован и содержит специализированный комплекс белков. Его длина составляет примерно от 20 до 60 мкм, и он функционирует как место инициации потенциала действия.[10][11] Как положение на аксоне, так и длина AIS могут изменяться, показывая степень пластичности, которая может точно настроить вывод нейронов.[10][12] Более длинный AIS связан с большей возбудимостью.[12] Пластичность также проявляется в способности AIS изменять свое распределение и поддерживать активность нейронных схем на постоянном уровне.[13]
AIS узкоспециализирована для быстрого проведения нервные импульсы. Это достигается за счет высокой концентрации напряжение-управляемые натриевые каналы в начальном сегменте, где возникает потенциал действия.[13] Ионные каналы сопровождаются большим количеством молекулы клеточной адгезии и каркасные белки, которые прикрепляют их к цитоскелету.[10] Взаимодействие с Анкирин Г важны, так как это главный организатор в САИ.[10]
Аксональный транспорт
В аксоплазма эквивалентен цитоплазма в клетка. Микротрубочки образуются в аксоплазме на бугорке аксона. Они расположены по длине аксона в перекрывающихся участках и все направлены в одном направлении — к окончанию аксона.[14] Об этом говорят положительные окончания микротрубочек. Такое перекрывающееся расположение обеспечивает маршруты транспортировки различных материалов из тела клетки.[14] Исследования аксоплазмы показали движение многочисленных пузырьков всех размеров, которые можно увидеть вдоль цитоскелетных филаментов — микротрубочек и нейрофиламенты, в обоих направлениях между аксоном и его окончаниями и телом клетки.
Исходящий антероградный транспорт из тела клетки по аксону переносит митохондрии и мембранные белки, необходимые для роста, к концу аксона. Входящий ретроградный транспорт переносит отходы клетки от терминала аксона к телу клетки.[15] Исходящие и входящие треки используют разные наборы моторные белки.[14] Исходящий транспорт обеспечивает кинезин, а входящий обратный трафик обеспечивается динеин. Динеин направлен на минус-конец.[15] Существует множество форм моторных белков кинезина и динеина, и считается, что каждая из них несет свой груз.[14] Исследования транспорта в аксоне привели к названию кинезина.[14]
Миелинизация

ТЕМ миелинизированного аксона в поперечном сечении.

В нервной системе аксоны могут быть миелинизированный, или немиелинизированные. Это обеспечение изолирующего слоя, называемого миелиновой оболочкой. Миелиновая мембрана уникальна своим относительно высоким соотношением липидов к белку.[16]
В периферической нервной системе аксоны миелинизируются глиальные клетки известный как Клетки Шванна. В центральной нервной системе миелиновую оболочку представляют глиальные клетки другого типа, олигодендроцит. Клетки Шванна миелинизируют единственный аксон. Олигодендроцит может миелинизировать до 50 аксонов.[17]
Состав миелина у этих двух типов разный. В ЦНС основным белком миелина является протеолипидный белок, а в ПНС это основной белок миелина.
Узлы Ранвье
Узлы Ранвье (также известен как щели миелиновой оболочки) — короткие немиелинизированные сегменты миелинизированный аксон, которые находятся периодически вкраплениями между сегментами миелиновой оболочки. Следовательно, в точке узла Ранвье аксон уменьшается в диаметре.[18] Эти узлы являются областями, где могут быть созданы потенциалы действия. В скачкообразное проведение электрические токи, возникающие в каждом узле Ранвье, передаются с небольшим затуханием к следующему узлу в линии, где они остаются достаточно сильными, чтобы генерировать другой потенциал действия. Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Терминалы Axon
Аксон может делиться на множество ветвей, называемых телодендриями (греч. Конец дерева). В конце каждого телодендрон является аксонный терминал (также называемый синаптическим бутоном или терминальным бутоном). Терминалы Axon содержат синаптические везикулы которые хранят нейротрансмиттер для выпуска в синапс. Это делает возможными множественные синаптические связи с другими нейронами. Иногда аксон нейрона может синапсировать с дендритами того же нейрона, когда он известен как autapse.
Потенциалы действия
| Структура типового химический синапс |
|---|
|
Постсинаптический Напряжение- Синаптический Нейротрансмиттер Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит |

Большинство аксонов несут сигналы в виде потенциалы действия, которые представляют собой дискретные электрохимические импульсы, которые быстро проходят по аксону, начиная с тела клетки и заканчивая в точках, где аксон производит синаптический контакт с клетками-мишенями. Определяющей характеристикой потенциала действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Когда потенциал действия достигает пресинаптического терминала, он активирует процесс синаптической передачи. Первый шаг — это быстрое открытие каналов для ионов кальция в мембране аксона, позволяя ионам кальция проходить внутрь через мембрану. Возникающее в результате повышение концентрации внутриклеточного кальция вызывает синаптические везикулы (крошечные контейнеры, окруженные липидной мембраной), заполненные нейротрансмиттер химикат, чтобы сливаться с мембраной аксона и выводить их содержимое во внеклеточное пространство. Нейромедиатор высвобождается из пресинаптического нерва через экзоцитоз. Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или каким-либо образом изменяющем ее метаболизм. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор везикул перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона.[5]

(A) пирамидная ячейка, интернейрон и форма волны короткой длительности (Axon), наложение трех средних форм волны;
(B) Средняя и стандартная ошибка времени прохождения пика для интернейронов пирамидных клеток и предполагаемых аксонов;
(C) График разброса отношения сигнал / шум для отдельных единиц от времени пика для аксонов, пирамидных клеток (PYR) и интернейронов (INT).
Внеклеточные записи потенциал действия распространение в аксонах было продемонстрировано у свободно перемещающихся животных. В то время как внеклеточные соматические потенциалы действия использовались для изучения клеточной активности у свободно перемещающихся животных, таких как разместить клетки, аксональная активность в обоих белый и серое вещество также могут быть записаны. Внеклеточные записи распространения потенциала действия аксона отличаются от соматических потенциалов действия по трем причинам: 1. Сигнал имеет более короткую продолжительность пика и спада (~ 150 мкс), чем у пирамидных клеток (~ 500 мкс) или интернейронов (~ 250 мкс). 2. Изменение напряжения трехфазное. 3. Активность, записанная на тетроде, видна только на одном из четырех проводов записи. В записях от свободно движущихся крыс аксональные сигналы были изолированы в трактах белого вещества, включая альвеус и мозолистое тело, а также серое вещество гиппокампа.[19]
Фактически, поколение потенциалы действия in vivo носит последовательный характер, и эти последовательные всплески составляют цифровые коды в нейроны. Хотя предыдущие исследования указывают на аксональное происхождение одиночного спайка, вызванного кратковременными импульсами, физиологические сигналы in vivo запускают инициирование последовательных спайков в телах клеток нейронов.[20][21]
В дополнение к распространению потенциалов действия к окончанию аксона, аксон способен усиливать потенциалы действия, что гарантирует безопасное распространение последовательных потенциалов действия к окончанию аксона. Что касается молекулярных механизмов, напряжение-управляемые натриевые каналы в аксонах обладают нижними порог и короче период отражения в ответ на кратковременные импульсы.[22]
Развитие и рост
Разработка
Развитие аксона к его цели — один из шести основных этапов в общей развитие нервной системы.[23] Исследования проведены на культивированных гиппокамп нейроны предполагают, что нейроны изначально производят несколько невриты которые эквивалентны, но только одному из этих нейритов суждено стать аксоном.[24] Неясно, предшествует ли спецификация аксона удлинению аксона или наоборот,[25] хотя недавние свидетельства указывают на последнее. Если разрезать не полностью развитый аксон, полярность может измениться, и другие нейриты потенциально могут стать аксоном. Это изменение полярности происходит только в том случае, если аксон сокращен как минимум на 10 мкм короче, чем другие нейриты. После того, как разрез будет сделан, самый длинный нейрит станет будущим аксоном, а все остальные нейриты, включая исходный аксон, превратятся в дендриты.[26] Воздействие внешней силы на нейрит, заставляющее его удлиниться, превращает его в аксон.[27] Тем не менее, развитие аксонов достигается за счет сложного взаимодействия между внеклеточной передачей сигналов, внутриклеточной передачей сигналов и цитоскелет динамика.
Внеклеточные сигналы, распространяющиеся через внеклеточный матрикс окружающие нейроны играют важную роль в развитии аксонов.[28] Эти сигнальные молекулы включают белки, нейротрофические факторы, внеклеточный матрикс и молекулы адгезии. Нетрин (также известный как UNC-6) секретируемый белок, участвующий в образовании аксонов. Когда UNC-5 рецептор нетрина мутирован, несколько нейритов нерегулярно проецируются из нейронов, и, наконец, один аксон выходит вперед.[29][30][31][32] Нейротрофические факторы — фактор роста нервов (NGF), нейротрофический фактор головного мозга (BDNF) и нейротрофин-3 (NTF3) также участвуют в развитии аксонов и связываются с Рецепторы Trk.[33]
В ганглиозид -превращающий фермент ганглиозид плазматической мембраны сиалидаза (PMGS), который участвует в активации TrkA на кончике нейтритов, требуется для удлинения аксонов. PMGS асимметрично распределяется по кончику нейрита, которому суждено стать будущим аксоном.[34]
Внутриклеточная передача сигналов
Во время развития аксонов активность PI3K увеличивается на конце предназначенного аксона. Нарушение активности PI3K тормозит развитие аксонов. Активация PI3K приводит к образованию фосфатидилинозит (3,4,5) -трисфосфат (PtdIns), которые могут вызывать значительное удлинение нейрита, превращая его в аксон. Таким образом, чрезмерное проявление фосфатазы что дефосфорилирование PtdIns приводит к нарушению поляризации.[28]
Цитоскелетная динамика
Неврит с самым низким актин содержимое филамента станет аксоном. Концентрация PGMS и f-актин содержание обратно коррелировано; когда PGMS становится обогащенным на кончике нейрита, содержание в нем f-актина существенно снижается.[34] Кроме того, воздействие препаратов, деполимеризующих актин, и токсина B (который инактивирует Rho-сигнализация ) вызывает образование множественных аксонов. Следовательно, разрыв актиновой сети в конусе роста будет способствовать превращению ее нейрита в аксон.[35]
Рост

Аксон девятидневной мыши с видимым конусом роста
Растущие аксоны перемещаются в окружающей среде через конус роста, который находится на кончике аксона. Конус роста имеет широкое пластинчатое расширение, называемое ламеллиподиум которые содержат выступы, называемые филоподия. Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует окружающую среду. Актин играет важную роль в мобильности этой системы. Среды с высоким уровнем молекулы клеточной адгезии (CAM) создают идеальную среду для роста аксонов. Похоже, что это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нейронных систем, включают: N-CAM, ТЕГ-1 — аксональный гликопротеин —[36]-и МАГ, все из которых являются частью иммуноглобулин надсемейство. Другой набор молекул называется внеклеточный матрикс -молекулы адгезии также обеспечивают липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин, фибронектин, тенасцин, и перлекан. Некоторые из них поверхностно связаны с клетками и, таким образом, действуют как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь эффекты длительного действия.
Ячейки называются клетки-указатели помочь в руководство роста аксонов нейронов. Эти клетки, которые помогают управление аксоном, как правило, другие нейроны, которые иногда незрелые. Когда аксон завершит свой рост в месте его соединения с мишенью, диаметр аксона может увеличиться до пяти раз, в зависимости от скорость проведения требуется.[37]
В ходе исследований также было обнаружено, что если аксоны нейрона были повреждены, то до тех пор, пока сома (тело клетки нейрон ) не повреждается, аксоны регенерируют и восстанавливают синаптические связи с нейронами с помощью клетки-указатели. Это также называется нейрорегенерация.[38]
Ного-А это тип компонента, ингибирующего рост нейритов, который присутствует в миелиновых мембранах центральной нервной системы (обнаружен в аксоне). Он играет решающую роль в ограничении регенерации аксонов в центральной нервной системе взрослых млекопитающих. В недавних исследованиях, если Nogo-A заблокирован и нейтрализован, можно вызвать регенерацию аксонов на большом расстоянии, что приводит к усилению функционального восстановления у крыс и спинного мозга мыши. Этого еще предстоит сделать на людях.[39] Недавнее исследование также показало, что макрофаги активируется специфическим воспалительным путем, активируемым Дектин-1 рецепторы способны способствовать восстановлению аксонов, однако также вызывают нейротоксичность в нейроне.[40]
Регулировка длины
Аксоны в значительной степени различаются по длине от нескольких микрометров до метров у некоторых животных. Это подчеркивает, что должен существовать механизм регулирования длины клетки, позволяющий нейронам ощущать длину своих аксонов и соответственно контролировать их рост. Было обнаружено, что моторные белки играют важную роль в регулировании длины аксонов.[41] Основываясь на этом наблюдении, исследователи разработали четкую модель роста аксонов, описывающую, как моторные белки могут влиять на длину аксона на молекулярном уровне.[42][43][44][45] Эти исследования предполагают, что моторные белки переносят сигнальные молекулы от сомы к конусу роста и наоборот, концентрация которых колеблется во времени с частотой, зависящей от длины.
Классификация
Аксоны нейронов человека периферическая нервная система могут быть классифицированы по их физическим характеристикам и свойствам проводимости сигнала. Известно, что аксоны имеют разную толщину (от 0,1 до 20 мкм).[3] и считалось, что эти различия связаны со скоростью, с которой потенциал действия может перемещаться по аксону — его скорость проводимости. Эрлангер и Гассер доказали эту гипотезу и идентифицировали несколько типов нервных волокон, установив связь между диаметром аксона и его скорость нервной проводимости. Они опубликовали свои открытия в 1941 году, дав первую классификацию аксонов.
Аксоны подразделяются на две системы. Первый, введенный Эрлангером и Гассером, сгруппировал волокна в три основные группы, используя буквы A, B и C. группа А, группа B, и группа C включают оба сенсорных волокна (афференты ) и двигательные волокна (эфференты ). Первая группа A была разделена на альфа, бета, гамма и дельта волокна — Aα, Aβ, Aγ и Aδ. Моторные нейроны различных моторных волокон были нижние двигательные нейроны – альфа двигательный нейрон, бета мотонейрон, и гамма мотонейрон с нервными волокнами Aα, Aβ и Aγ соответственно.
Позже другие исследователи обнаружили две группы волокон Aa, которые были сенсорными волокнами. Затем они были введены в систему, которая включала только сенсорные волокна (хотя некоторые из них были смешанными нервами, а также двигательными волокнами). Эта система называет сенсорные группы Типами и использует римские цифры: Тип Ia, Тип Ib, Тип II, Тип III и Тип IV.
Мотор
Нижние двигательные нейроны имеют два вида волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин | Проведение скорость (м / с) |
Связанный мышечные волокна |
|---|---|---|---|---|---|
| α | Aα | 13-20 | да | 80–120 | Экстрафузионные мышечные волокна |
| β | Aβ | ||||
| γ | Aγ | 5-8 | да | 4–24[46][47] | Внутрифузионные мышечные волокна |
Сенсорный
Разные Рецепторы чувств иннервируют разные типы нервных волокон. Проприоцепторы иннервируются сенсорными волокнами типа Ia, Ib и II, механорецепторы сенсорными волокнами II и III типов и ноцицепторы и терморецепторы сенсорными волокнами III и IV типа.
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин | Проведение скорость (м / с) |
Связанный Рецепторы чувств | Проприоцепторы | Механоцепторы | Ноцицепторы и терморецепторы |
|---|---|---|---|---|---|---|---|---|
| Я | Aα | 13-20 | да | 80–120 | Первичные рецепторы мышечное веретено (аннулоспиральное окончание) | ✔ | ||
| Ib | Aα | 13-20 | да | 80–120 | Орган сухожилия Гольджи | |||
| II | Aβ | 6-12 | да | 33–75 | Вторичные рецепторы мышечное веретено (окончание цветочно-спрей). Все кожные механорецепторы |
✔ | ||
| III | Aδ | 1-5 | Тонкий | 3–30 | Свободные нервные окончания прикосновения и давления Ноцицепторы из боковой спиноталамический тракт Холодный терморецепторы |
✔ | ||
| IV | C | 0.2-1.5 | Нет | 0.5-2.0 | Ноцицепторы из передний спиноталамический тракт Рецепторы тепла |
Автономный
В автономная нервная система имеет два вида периферических волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин[48] | Проведение скорость (м / с) |
|---|---|---|---|---|
| преганглионарные волокна | B | 1–5 | да | 3–15 |
| постганглионарные волокна | C | 0.2–1.5 | Нет | 0.5–2.0 |
Клиническое значение
В порядке степени тяжести повреждение нерва можно описать как неврапраксия, аксонотмезис, или же невротмезис.Сотрясение считается легкой формой диффузное повреждение аксонов.[49] Аксональное повреждение также может вызвать центральный хроматолиз. Нарушение функции аксонов нервной системы является одной из основных причин многих наследственных заболеваний. неврологические расстройства которые влияют как на периферические, так и на центральные нейроны.[5]
Когда аксон раздавлен, активный процесс дегенерация аксонов происходит в части аксона, наиболее удаленной от тела клетки. Эта дегенерация происходит быстро после травмы, когда часть аксона блокируется мембранами и разрушается макрофагами. Это известно как Валлеровское вырождение.[50] Отмирание аксона также может иметь место при многих нейродегенеративных заболеваниях, особенно при нарушении транспорта аксонов, это известно как дегенерация, подобная валлеровской.[51] Исследования показывают, что дегенерация происходит в результате действия аксонального белка. NMNAT2, будучи не в состоянии достичь всего аксона.[52]
Демиелинизация аксонов вызывает множество неврологических симптомов, обнаруженных при заболевании рассеянный склероз.
Дисмиелинизация это аномальное образование миелиновой оболочки. Это связано с несколькими лейкодистрофии, а также в шизофрения.[53][54][55]
Суровый травматическое повреждение мозга может привести к обширным поражениям нервных путей, повреждая аксоны в состоянии, известном как диффузное повреждение аксонов. Это может привести к стойкое вегетативное состояние.[56] Это было показано в исследованиях на крыса такое повреждение аксонов в результате единичного легкого черепно-мозгового повреждения может оставить восприимчивость к дальнейшему повреждению после повторных легких черепно-мозговых травм.[57]
А нервный проводник является искусственным средством направления роста аксонов, чтобы нейрорегенерация, и является одним из многих методов лечения, используемых для различных видов повреждение нерва.
История
Немецкий анатом Отто Фридрих Карл Дайтерс обычно приписывают открытие аксона, отличив его от дендритов.[5] Швейцарский Рюдольф Альберт фон Кёлликер и немецкий Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Келликер назвал аксон в 1896 году.[58] Луи-Антуан Ранвье был первым, кто описал бреши или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлы Ранвье. Сантьяго Рамон-и-Кахаль, испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции.[5] Джозеф Эрлангер и Герберт Гассер ранее разработали систему классификации периферических нервных волокон,[59] на основе скорости аксональной проводимости, миелинизация, размер волокна и т. д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), а к 1952 году они получили полное количественное описание ионной основы потенциал действия, что привело к формулировке Модель Ходжкина – Хаксли. Ходжкин и Хаксли были награждены совместно Нобелевская премия для этой работы в 1963 году. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхойзера-Хаксли. Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает много деталей об отдельных ионные каналы.
Другие животные
Аксоны в беспозвоночные были широко изучены. В Прибрежный кальмар, часто используется как модельный организм имеет самый длинный из известных аксонов.[60] В Гигантский кальмар имеет самый большой аксон известен. Его размер варьируется от половины (обычно) до одного миллиметра в диаметре и используется для управления его реактивный двигатель система. Самая быстрая зарегистрированная скорость проводимости 210 м / с обнаружена в заключенных в оболочку аксонах некоторых пелагических Креветки Penaeid[61] и обычный диапазон составляет от 90 до 200 м / с.[62] (ср 100–120 м / с для аксона самого быстрого миелинизированного позвоночного.)
В других случаях, как показали исследования на крысах, аксон происходит от дендрита; такие аксоны, как говорят, имеют «дендритное происхождение». Некоторые аксоны с дендритным происхождением аналогичным образом имеют «проксимальный» начальный сегмент, который начинается непосредственно в источнике аксона, в то время как другие имеют «дистальный» начальный сегмент, заметно отделенный от источника аксона.[63] У многих видов некоторые из нейронов имеют аксоны, которые исходят из дендрита, а не из тела клетки, и они известны как дендриты, несущие аксоны.[1] Во многих случаях аксон берет свое начало от бугорка аксона на соме; такие аксоны, как говорят, имеют «соматическое происхождение». Некоторые аксоны соматического происхождения имеют «проксимальный» начальный сегмент, прилегающий к бугорку аксона, в то время как другие имеют «дистальный» начальный сегмент, отделенный от сомы протяженным бугорком аксона.[63]
Смотрите также
- Электрофизиология
- Ганглионарное возвышение
- Гигантская аксональная нейропатия
- Нейрональная трассировка
- Пионерский аксон
Рекомендации
- ^ а б Triarhou LC (2014). «Аксоны, исходящие из дендритов: филогенетические последствия с оттенками Кахаля». Границы нейроанатомии. 8: 133. Дои:10.3389 / fnana.2014.00133. ЧВК 4235383. PMID 25477788.
- ^ Яу К.В. (декабрь 1976 г.). «Рецептивные поля, геометрия и проводящий блок сенсорных нейронов центральной нервной системы пиявки». Журнал физиологии. 263 (3): 513–38. Дои:10.1113 / jphysiol.1976.sp011643. ЧВК 1307715. PMID 1018277.
- ^ а б c Сквайр, Ларри (2013). Фундаментальная нейробиология (4-е изд.). Амстердам: Elsevier / Academic Press. С. 61–65. ISBN 978-0-12-385-870-2.
- ^ а б c Людерс Э., Томпсон П.М., Тога А.В. (август 2010 г.). «Развитие мозолистого тела в мозгу здорового человека». Журнал неврологии. 30 (33): 10985–90. Дои:10.1523 / JNEUROSCI.5122-09.2010. ЧВК 3197828. PMID 20720105.
- ^ а б c d е Дебанн Д., Кампанак Е., Бяловас А., Карлье Е., Алькарас Г. (апрель 2011 г.). «Аксонная физиология» (PDF). Физиологические обзоры. 91 (2): 555–602. Дои:10.1152 / Physrev.00048.2009. PMID 21527732. S2CID 13916255.
- ^ Нельсон А.Д., Дженкинс П.М. (2017). «Аксональные мембраны и их домены: сборка и функция начального сегмента аксона и узла Ранвье». Границы клеточной неврологии. 11: 136. Дои:10.3389 / fncel.2017.00136. ЧВК 5422562. PMID 28536506.
- ^ Летерье С., Клерк Н., Руэда-Борони Ф, Монтерсино А., Дарджент Б., Кастетс Ф (2017). «Мембранные партнеры Ankyrin G стимулируют создание и поддержание начального сегмента Axon». Границы клеточной неврологии. 11: 6. Дои:10.3389 / fncel.2017.00006. ЧВК 5266712. PMID 28184187.
- ^ Летерье С (февраль 2018 г.). «Начальный сегмент аксона: обновленная точка зрения». Журнал неврологии. 38 (9): 2135–2145. Дои:10.1523 / jneurosci.1922-17.2018. ЧВК 6596274. PMID 29378864.
- ^ Расбанд М.Н. (август 2010 г.). «Начальный сегмент аксона и поддержание полярности нейронов». Обзоры природы. Неврология. 11 (8): 552–62. Дои:10.1038 / nrn2852. PMID 20631711. S2CID 23996233.
- ^ а б c d Джонс С.Л., Свиткина Т.М. (2016). «Цитоскелет начального сегмента аксона: архитектура, развитие и роль в полярности нейронов». Нейронная пластичность. 2016: 6808293. Дои:10.1155/2016/6808293. ЧВК 4967436. PMID 27493806.
- ^ Кларк Б.Д., Голдберг Е.М., Руди Б. (декабрь 2009 г.). «Электрогенная настройка начального сегмента аксона». Нейробиолог. 15 (6): 651–68. Дои:10.1177/1073858409341973. ЧВК 2951114. PMID 20007821.
- ^ а б Ямада Р., Куба Х (2016). «Структурная и функциональная пластичность в начальном сегменте аксона». Границы клеточной неврологии. 10: 250. Дои:10.3389 / fncel.2016.00250. ЧВК 5078684. PMID 27826229.
- ^ а б Сусуки К., Куба Х (март 2016 г.). «Активно-зависимая регуляция возбудимых аксональных доменов». Журнал физиологических наук. 66 (2): 99–104. Дои:10.1007 / s12576-015-0413-4. PMID 26464228. S2CID 18862030.
- ^ а б c d е Альбертс Б. (2004). Essential Cell Biology: введение в молекулярную биологию клетки (2-е изд.). Нью-Йорк: Гарленд. стр.584–587. ISBN 978-0-8153-3481-1.
- ^ а б Альбертс Б. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Гарленд. С. 979–981. ISBN 978-0-8153-4072-0.
- ^ Озген, Н; Барон, W; Hoekstra, D; Кахья, Н. (сентябрь 2016 г.). «Динамика олигодендроглиальной мембраны в связи с биогенезом миелина». Клеточные и молекулярные науки о жизни. 73 (17): 3291–310. Дои:10.1007 / s00018-016-2228-8. ЧВК 4967101. PMID 27141942.
- ^ Сэдлер, Т. (2010). Медицинская эмбриология Лангмана (11-е изд.). Филадельфия: Липпинкотт Уильям и Уилкинс. п.300. ISBN 978-0-7817-9069-7.
- ^ Гесс А., Янг Дж. З. (ноябрь 1952 г.). «Узлы Ранвье». Труды Лондонского королевского общества. Серия B, Биологические науки. Серия Б. 140 (900): 301–20. Bibcode:1952RSPSB.140..301H. Дои:10.1098 / rspb.1952.0063. JSTOR 82721. PMID 13003931. S2CID 11963512.
- ^ Роббинс А.А., Фокс С.Е., Холмс Г.Л., Скотт Р.К., Барри Дж. М. (ноябрь 2013 г.). «Кратковременные сигналы, записанные внеклеточно у свободно движущихся крыс, представляют активность аксонов». Границы в нейронных цепях. 7 (181): 181. Дои:10.3389 / fncir.2013.00181. ЧВК 3831546. PMID 24348338.
- ^ Rongjing Ge, Hao Qian и Jin-Hui Wang * (2011) Molecular Brain 4 (19), 1 ~ 11
- ^ Rongjing Ge, Hao Qian, Na Chen и Jin-Hui Wang * (2014) Molecular Brain 7 (26): 1-16
- ^ Чен Н, Ю Дж, Цянь Х, Ге Р, Ван Дж Х (июль 2010 г.). «Аксоны усиливают соматические неполные спайки до однородных амплитуд в пирамидных нейронах коры головного мозга мышей». PLOS ONE. 5 (7): e11868. Bibcode:2010PLoSO … 511868C. Дои:10.1371 / journal.pone.0011868. ЧВК 2912328. PMID 20686619.
- ^ Вольперт, Льюис (2015). Принципы развития (5-е изд.). С. 520–524. ISBN 978-0-19-967814-3.
- ^ Флетчер Т.Л., Банкир Г.А. (декабрь 1989 г.). «Установление полярности нейронами гиппокампа: взаимосвязь между стадией развития клетки in situ и ее последующим развитием в культуре». Биология развития. 136 (2): 446–54. Дои:10.1016/0012-1606(89)90269-8. PMID 2583372.
- ^ Цзян Х., Рао Й. (май 2005 г.). «Формирование аксона: судьба против роста». Природа Неврология. 8 (5): 544–6. Дои:10.1038 / nn0505-544. PMID 15856056. S2CID 27728967.
- ^ Гослин К., Банкир Г. (апрель 1989 г.). «Экспериментальные наблюдения за развитием полярности нейронов гиппокампа в культуре». Журнал клеточной биологии. 108 (4): 1507–16. Дои:10.1083 / jcb.108.4.1507. ЧВК 2115496. PMID 2925793.
- ^ Lamoureux P, Ruthel G, Buxbaum RE, Heidemann SR (ноябрь 2002 г.). «Механическое напряжение может определять судьбу аксонов в нейронах гиппокампа». Журнал клеточной биологии. 159 (3): 499–508. Дои:10.1083 / jcb.200207174. ЧВК 2173080. PMID 12417580.
- ^ а б Аримура Н., Кайбути К. (март 2007 г.). «Полярность нейронов: от внеклеточных сигналов к внутриклеточным механизмам». Обзоры природы. Неврология. 8 (3): 194–205. Дои:10.1038 / номер 2056. PMID 17311006. S2CID 15556921.
- ^ Нейроглия и пионерные нейроны express UNC-6 для предоставления глобальных и локальных сетевых сигналов для управления миграциями в C. elegans
- ^ Серафини Т., Кеннеди Т.Э., Галко М.Дж., Мирзаян С., Джессел Т.М., Тесье-Лавин М. (август 1994 г.). «Нетрины определяют семейство белков, способствующих отрастанию аксонов, гомологичных C. elegans UNC-6». Клетка. 78 (3): 409–24. Дои:10.1016/0092-8674(94)90420-0. PMID 8062384. S2CID 22666205.
- ^ Хонг К., Хинк Л., Нишияма М., Пу М.М., Тесье-Лавин М., Штейн Э. (июнь 1999 г.). «Управляемая лигандом ассоциация между цитоплазматическими доменами рецепторов семейства UNC5 и DCC превращает индуцированное нетрином притяжение конуса роста в отталкивание». Клетка. 97 (7): 927–41. Дои:10.1016 / S0092-8674 (00) 80804-1. PMID 10399920. S2CID 18043414.
- ^ Hedgecock EM, Culotti JG, Hall DH (январь 1990 г.). «Гены unc-5, unc-6 и unc-40 управляют периферической миграцией первичных аксонов и мезодермальных клеток по эпидермису C. elegans». Нейрон. 4 (1): 61–85. Дои:10.1016 / 0896-6273 (90) 90444-К. PMID 2310575. S2CID 23974242.
- ^ Хуанг Э.Дж., Райхардт Л.Ф. (2003). «Рецепторы Trk: роли в передаче сигнала нейронов». Ежегодный обзор биохимии. 72: 609–42. Дои:10.1146 / annurev.biochem.72.121801.161629. PMID 12676795. S2CID 10217268.
- ^ а б Да Силва Дж. С., Хасегава Т., Мияги Т., Дотти К. Г., Абад-Родригес Дж. (Май 2005 г.). «Активность асимметричной мембранной ганглиозидной сиалидазы определяет судьбу аксонов». Природа Неврология. 8 (5): 606–15. Дои:10.1038 / nn1442. PMID 15834419. S2CID 25227765.
- ^ Брадке Ф., Дотти К.Г. (март 1999 г.). «Роль локальной нестабильности актина в формировании аксонов». Наука. 283 (5409): 1931–4. Bibcode:1999Научный … 283.1931B. Дои:10.1126 / science.283.5409.1931. PMID 10082468.
- ^ Ферли А.Дж., Мортон С.Б., Манало Д., Карагогеос Д., Додд Дж., Джессел Т.М. (апрель 1990 г.). «Аксональный гликопротеин TAG-1 является членом суперсемейства иммуноглобулинов с активностью, способствующей росту нейритов». Клетка. 61 (1): 157–70. Дои:10.1016/0092-8674(90)90223-2. PMID 2317872. S2CID 28813676.
- ^ Альбертс, Брюс (2015). Молекулярная биология клетки (Шестое изд.). п. 947. ISBN 9780815344643.
- ^ Куник Д., Дион С., Одзаки Т., Левин Л.А., Константино С. (2011). «Лазерная пересечение одного аксона для исследований повреждения и регенерации аксонов с высоким содержанием аксонов». PLOS ONE. 6 (11): e26832. Bibcode:2011PLoSO … 626832K. Дои:10.1371 / journal.pone.0026832. ЧВК 3206876. PMID 22073205.
- ^ Schwab ME (февраль 2004 г.). «Регенерация ног и аксонов». Текущее мнение в нейробиологии. 14 (1): 118–24. Дои:10.1016 / j.conb.2004.01.004. PMID 15018947. S2CID 9672315.
- ^ Гензель Дж. К., Накамура С., Гуан З., ван Ройен Н., Анкени Д. П., Попович П. Г. (март 2009 г.). «Макрофаги способствуют регенерации аксонов с одновременной нейротоксичностью». Журнал неврологии. 29 (12): 3956–68. Дои:10.1523 / JNEUROSCI.3992-08.2009. ЧВК 2693768. PMID 19321792.
- ^ Майерс К.А., Баас П.В. (сентябрь 2007 г.). «Кинезин-5 регулирует рост аксона, действуя как тормоз на его массив микротрубочек». Журнал клеточной биологии. 178 (6): 1081–91. Дои:10.1083 / jcb.200702074. ЧВК 2064629. PMID 17846176.
- ^ Ришал И., Кам Н., Перри Р. Б., Шиндер В., Фишер Е. М., Скьяво Г., Файнзильбер М. (июнь 2012 г.). «Механизм с приводом от двигателя для измерения длины клетки». Отчеты по ячейкам. 1 (6): 608–16. Дои:10.1016 / j.celrep.2012.05.013. ЧВК 3389498. PMID 22773964.
- ^ Карамчед Б.Р., Бресслофф ПК (май 2015 г.). «Модель отсроченной обратной связи для измерения длины аксонов». Биофизический журнал. 108 (9): 2408–19. Bibcode:2015BpJ … 108.2408K. Дои:10.1016 / j.bpj.2015.03.055. ЧВК 4423051. PMID 25954897.
- ^ Бресслов П.С., Карамчед Б.Р. (2015). «Механизм частотно-зависимого декодирования для определения длины аксона». Границы клеточной неврологии. 9: 281. Дои:10.3389 / fncel.2015.00281. ЧВК 4508512. PMID 26257607.
- ^ Фольц Ф., Веттманн Л., Мориджи Г., Круз К. (май 2019 г.). «Звук роста аксона». Физический обзор E. 99 (5–1): 050401. arXiv:1807.04799. Bibcode:2019PhRvE..99e0401F. Дои:10.1103 / PhysRevE.99.050401. PMID 31212501.
- ^ Эндрю Б.Л., часть Нью-Джерси (апрель 1972 г.). «Свойства быстрых и медленных двигательных единиц в мышцах задних конечностей и хвоста крысы». Ежеквартальный журнал экспериментальной физиологии и когнитивных медицинских наук. 57 (2): 213–25. Дои:10.1113 / expphysiol.1972.sp002151. PMID 4482075.
- ^ Рассел Нью-Джерси (январь 1980 г.). «Скорость аксональной проводимости изменяется после тенотомии мышц или деафферентации во время развития у крысы». Журнал физиологии. 298: 347–60. Дои:10.1113 / jphysiol.1980.sp013085. ЧВК 1279120. PMID 7359413.
- ^ Покок Г., Ричардс С.Д. и др. (2004). Физиология человека (2-е изд.). Нью-Йорк: Издательство Оксфордского университета. С. 187–189. ISBN 978-0-19-858527-5.
- ^ Dawodu ST (16 августа 2017 г.). «Травматическая травма головного мозга (ЧМТ) — определение, эпидемиология, патофизиология». Medscape. В архиве с оригинала 12 июня 2018 г.. Получено 14 июля 2018.
- ^ Травма и валлеровская дегенерация В архиве 2 мая 2006 г. Wayback Machine, Калифорнийский университет в Сан-Франциско
- ^ Coleman MP, Freeman MR (1 июня 2010 г.). «Валлеровское вырождение, мир (а) и сущность». Ежегодный обзор нейробиологии. 33 (1): 245–67. Дои:10.1146 / annurev-neuro-060909-153248. ЧВК 5223592. PMID 20345246.
- ^ Джилли Дж., Член парламента Коулмана (январь 2010 г.). «Эндогенный Nmnat2 является важным фактором выживания для поддержания здоровья аксонов». PLOS Биология. 8 (1): e1000300. Дои:10.1371 / journal.pbio.1000300. ЧВК 2811159. PMID 20126265.
- ^ Кремер-Альберс Е.М., Гериг-Бургер К., Тиле С., Троттер Дж., Наве К.А. (ноябрь 2006 г.). «Нарушенные взаимодействия мутантного протеолипидного белка / DM20 с холестерином и липидными рафтами в олигодендроглии: последствия для дисмиелинизации при спастической параплегии». Журнал неврологии. 26 (45): 11743–52. Дои:10.1523 / JNEUROSCI.3581-06.2006. ЧВК 6674790. PMID 17093095.
- ^ Маталон Р., Михалс-Маталон К., Сурендран С., Тайринг С.К. (2006). «Болезнь Канавана: исследования на нокаутной мыши». N-ацетиласпартат. Adv. Exp. Med. Биол. Успехи экспериментальной медицины и биологии. 576. С. 77–93, обсуждение 361–3. Дои:10.1007/0-387-30172-0_6. ISBN 978-0-387-30171-6. PMID 16802706. S2CID 44405442.
- ^ Ткачев Д., Миммак М.Л., Хаффакер С.Дж., Райан М., Bahn S (август 2007 г.). «Дальнейшие доказательства изменения биосинтеза миелина и глутаматергической дисфункции при шизофрении». Международный журнал нейропсихофармакологии. 10 (4): 557–63. Дои:10.1017 / S1461145706007334. PMID 17291371.
- ^ «Травма головного мозга, травматическая». Медциклопедия. GE. Архивировано из оригинал 26 мая 2011 г.
- ^ Райт Д.К., Брэди Р.Д., Камнакш А., Трезизе Дж., Сан М., Макдональд С.Дж. и др. (Октябрь 2019 г.). «Повторные легкие черепно-мозговые травмы вызывают стойкие изменения белков плазмы и биомаркеров магнитно-резонансной томографии у крыс». Научные отчеты. 9 (1): 14626. Дои:10.1038 / с41598-019-51267-ш. ЧВК 6787341. PMID 31602002.
- ^ Палец S (1994). Истоки нейробиологии: история исследований функций мозга. Издательство Оксфордского университета. п. 47. ISBN 9780195146943. OCLC 27151391.
Келликер дал название аксону в 1896 году.
- ^ Грант G (декабрь 2006 г.). «Нобелевские премии 1932 и 1944 годов по физиологии и медицине: награды за новаторские исследования в области нейрофизиологии». Журнал истории неврологии. 15 (4): 341–57. Дои:10.1080/09647040600638981. PMID 16997762. S2CID 37676544.
- ^ Хелье, Дженнифер Л. (16 декабря 2014 г.). Мозг, нервная система и их болезни [3 тома]. ABC-CLIO. ISBN 9781610693387. В архиве из оригинала 14 марта 2018 г.
- ^ Хсу К., Теракава С. (июль 1996 г.). «Фенестрация в миелиновой оболочке нервных волокон креветки: новый узел возбуждения для скачкообразной проводимости». Журнал нейробиологии. 30 (3): 397–409. Дои:10.1002 / (SICI) 1097-4695 (199607) 30: 3 <397 :: AID-NEU8> 3.0.CO; 2- #. PMID 8807532.
- ^ Зальцер Дж. Л., Залц Б. (октябрь 2016 г.). «Миелинизация». Текущая биология. 26 (20): R971 – R975. Дои:10.1016 / j.cub.2016.07.074. PMID 27780071.
- ^ а б Хёффлин Ф., Джек А., Ридель С., Мак-Бухер Дж., Роос Дж., Корчелли С. и др. (2017). «Неоднородность начального сегмента аксона в интернейронах и пирамидных клетках зрительной коры грызунов». Границы клеточной неврологии. 11: 332. Дои:10.3389 / fncel.2017.00332. ЧВК 5684645. PMID 29170630.
внешняя ссылка
- Гистологическое изображение: 3_09 в Центре медицинских наук Университета Оклахомы — «Слайд 3 Спинной мозг «
Ещё со времён изучения анатомии в средней школе большинство из нас знает, что наш мозг состоит из нейронов, а основная мыслительная деятельность происходит в коре головного мозга. При этом нам что-то рассказывали про большие полушария, мозжечок и прочие составляющие. Но, что интересно, нам при этом не рассказывали весьма важных вещей о тех процессах, на самом деле происходящих в головном мозге и нервной системе человека, которые весьма важны для понимания того, что и зачем мы делаем, в том числе в процессе обучения и различных тренировок.

Надеюсь, что если вы найдёте немного времени на то, чтобы изучить данную статью, это поможет вам более рационально и эффективно построить свою жизнь и использовать с пользой для себя возможности вашего организма.
В организме человека выделяют центральную и периферическую нервные системы. В центральную нервную систему входит головной и спиной мозг. К периферической нервной системе относят остальные нейроны, которые пронизывают все ткани человека, собирая информацию о состоянии этих тканей и передавая им управляющие сигналы от центральной нервной системы. Именно за счёт нейронов периферической нервной системы мы чувствуем боль, которая сообщает нам о том, что с теми или иными органами что-то не в порядке.
На элементарном уровне нервная система человека состоит из нейронов (нервных клеток) и вспомогательных нейроглиальных клеток, которые помогают нейронам в выполнении их функций.

Нейрон состоит из тела клетки (2), или сомы, одного длинного мало ветвящегося отростка, называемого аксоном (4), а также множества (от 1 до 1000) коротких сильно ветвящихся отростков — дендритов (1). Также на схеме обозначены ядро клетки (3), ветви аксона (6), миелиновое волокно (5), перехват (7) и неврилемма (8).
Длина аксона достигает метра и более, его диаметр колеблется от сотых долей микрона до 10 мкм. Длина дендрита может достигать 300 мкм, а его диаметр 5 мкм.
Нейроны соединяются между собой, образуя так называемые нейронные сети. При этом дендриты нейронов, являющиеся входными линиями сигналов, присоединяются к аксонам других нейронов, по которым от нейрона передаются так называемые «нервные импульсы». Место соединения одного нейрона с другим получило название «синапса» (от греческого слова «синапта» — контактировать). Количество синаптических контактов неодинаково на теле и отростках нейрона и весьма отличается в различных частях нервной системы. Тело нейрона на 38% покрыто синапсами и их насчитывают до 1200-1800 на одном нейроне. Все нейроны центральной нервной системы соединяются между собой в основном в одном направлении: разветвления аксона одного нейрона контактируют с телом или дендритами других нейронов.
У нейронов из периферической нервной системы аксоны контактируют с тканями управляемых ими органов или клеткам мышечной ткани. То есть, импульс, передаваемый по аксону, воздействует не на другие нейроны, а заставляет, например, сокращаться клетки мышечной ткани.
При этом хочу особо обратить ваше внимание на тот факт, что на самом деле то, что во многих источниках называют «нервные импульсы», на самом деле является импульсами электрического тока, что очень хорошо демонстрируется на старом школьном опыте, когда мышцы на лягушачьей лапке начинают сокращаться под действием электрического тока. То есть в основе деятельности головного мозга лежат электромагнитные импульсы, которые распространяются по нейронной сети, образованной связями между нейронами.
Изначально нейрон находится в так называемом невозбуждённом состоянии. Через синапсы к нему поступают электрические импульсы от других нейронов, и когда общее количество этих импульсов достигает некоего порогового значения, нейрон переходит в возбуждённое состояние и по его аксону пробегает импульс электрического тока, передавая сигнал другим нейронам или заставляя мышечную ткань сокращаться.
Таким образом управление различными физиологическими процессами и наше мышление происходят за счёт распространения электрических импульсов в нейронной сети центральной и периферической нервных систем.
Распространяются эти импульсы не так уж быстро. Скорость распространения импульса через один синапс измерена и составляет порядка 3 миллисекунд. Это означает, что максимальная частота сигнала, которую вы сможете передать через такой контакт, составляет всего около 333 Гц. Нам, привыкшим к частотам процессоров в несколько гигагерц, скорость работы нервных клеток может показаться слишком низкой, но на самом деле это представление сильно ошибочно, поскольку нейронная сеть нашего головного мозга на самом деле обладает просто огромной вычислительной мощностью.
Летом 2013 года японские учёные провели моделирование работы нейронной сети, которая состояла из 1.73 млрд. нейронов, между которыми было установлено 10.4 трлн. синапсов (соединений). Для моделирования использовался суперкомпьютер Fujitsu K computer, который в ноябре 2013 года занимал 4-ое место в мире про общей производительности http://www.top500.org/list/2013/11/
Так вот на моделирование одной секунды работы данной нейронной сети у суперкомпьютера имеющего 705 024 ядер и потребляющего 12.6 КВт электроэнергии ушло целых 40 минут! http://www.telegraph.co.uk/technology/10567942/Supercomputer-models-one-second-of-human-brain-activity.html Считается, что в среднем в мозгу человека содержится порядка 86 млрд. нейронов. http://globalscience.ru/article/read/20324/ Это примерно в 50 раз больше, чем смоделированная нейронная сеть. При этом разница во времени составила 2400 раз (столько секунд в 40 минутах). Итого общая разница в скорости составляет порядка 120 000 раз. Добавьте к этому также тот объём, который занимает данный суперкомпьютер, а также то количество энергии, которое было на данные вычисления потрачены.
Другими словами, нашим ЭВМ пока ещё очень далеко до той эффективности и быстродействия, которые реализованы Природой в нашем головном мозге!
Но вернёмся к рассмотрению того, какие процессы происходят в нашем мозгу и всей нервной системе в целом. Есть три важных составляющих, которые обеспечивают её функционирование. Первый, о котором я уже говорил, это распространение электрических импульсов по нейронной сети. Это, если так можно выразиться, главный вычислительный процесс, который происходит постоянно. И именно он определяет нашу мыслительную деятельность и двигательную активность. Второй процесс основан на действии так называемых нейромедиаторов, которые формируют химический уровень регулирования нервной деятельности. В зависимости от того, какие нейромедиаторы выделяются организмом, скорость работы нейронов и всей нервной сети может либо повышаться, особенно в критических ситуациях, либо, наоборот, понижаться, когда состояние перевозбуждения требуется погасить и успокоиться, поскольку работа нейронов в ускоренном перевозбуждённом состоянии приводит к их преждевременному разрушению и отмиранию. А вот про третью важную составляющую в медицинской литературе вы не найдёте практически ничего! При том, что данная третья составляющая как раз и является одной из важнейших, поскольку именно она определяет качество работы всей нейронной сети, её функциональные возможности. Этой важнейшей составляющей является та структура связей, которая сформирована между нейронами, поскольку именно она определяет то, как и какие процессы происходят в этой нейронной сети во время её работы.

Главной особенностью нейронной сети, которую образуют наши нейроны, состоит в том, что она не является постоянной. Нейроны обладают способностью перестраивать связи между собой, меняя структуру нейронной сети. И в этом одно из её принципиальных отличий от наших современных компьютеров, которые в основном имеют фиксированную структуру вычислительных модулей.
Уникальность нашей нервной системы состоит в том, что она постоянно меняет свою структуру, оптимизируя её под решение тех или иных задач. При этом формирование связей между нейронами, в том числе и в головном мозге, начинается задолго до рождения ребёнка. Детерминация клеток плода, при которой уже можно выделить те клетки, из которых будут в дальнейшем сформированы лобные доли головного мозга, наблюдается уже на 25 день после зачатия. На сроке 100 дней уже сформированы основные отделы мозга и начинает формироваться его структура.

Это означает, что уже с этого момента всё, что происходит вокруг ребёнка, находящегося в утробе матери, будет оказывать влияние на ту структуру нейронной сети, которая в конечном итоге будет сформирована! Другими словами, способности и возможности будущего ребёнка начинают закладываться задолго до его рождения. Именно поэтому беременным девушкам и женщинам необходимо создавать более комфортные условия практически сразу после зачатия, а не на 6-7 месяце. Причём комфортные не столько в физическом смысле, сколько в психологическом, поскольку все эмоциональные переживания матери в конечном итоге передаются и будущему ребёнку.
Активный процесс формирования связей между нейронами, то есть программирование нейронной сети, продолжается и после рождения. На самом деле именно в формировании нужных связей и оптимизации их структуры и состоит смысл обучения. Только что родившийся ребёнок толком не умеет управлять своим телом. И не только потому, что у него ещё не окрепли кости и мышцы, но и потому, что в нервной системе не сформированы связи необходимые для управления движениями. Встроенные программы имеются только для обеспечения деятельности главных органов и систем, таких как сердце, лёгкие, печень, почки и т. п. Это формируется ещё на этапе развития плода в утробе матери по тем программам, которые прописаны в ДНК. А вот всё то, что связано с двигательной активностью, приобретается уже после рождения в процессе обучения.
Первые движения, например когда ребёнок учится ходить, делаются под полным контролем головного мозга, а потому происходят они медленно. В том числе и потому, что импульсы через синапсы распространяются достаточно медленно, как уже было сказано выше, около 3 мс на одно соединение. Если в этот процесс включён головной мозг, то количество соединений, которые участвуют в обработке информации, принятии решения и передачи управляющего сигнала мышцам, будет исчисляться десятками и сотнями. Но когда ребёнок многократно повторяет те или иные движения, в его нервной системе нейроны постепенно будут образовывать новые связи, за счёт которых время выполнения часто повторяющихся задач будет существенно сокращаться. И в какой-то момент головной мозг будет исключён из процесса обработки данного движения и оно начинает происходить рефлекторно, то есть только за счёт тех импульсов, которые проходят по периферической нервной системе. С этого момента человеку достаточно только подумать, что он хочет сделать, а как это сделать организм, точнее периферическая нервная система уже знает сама. В ней уже прошита соответствующая программа, которая и реализует требуемое движение, зачастую достаточно сложное.
Вспомните, как вы когда-то учились каким нибудь новым сложным движениям, например езде на велосипеде, катании на лыжах или катании, или тому же плаванию. В начале у вас ничего толком не получалось. Вам приходилось с помощью своего сознания контролировать все свои движения, куда повернуть руль велосипеда или как поставить ноги, чтобы затормозить на лыжах. Но если вы были настойчивы, то через некоторое время у вас начинало получаться всё лучше и лучше, и в какой-то момент вы вдруг начинали просто ехать на велосипеде не думая о том, куда крутить руль, чтобы не упасть или начинали гонятся с клюшкой за шайбой, не думая как правильно ставить коньки, чтобы повернуть и не упасть. В вашей нервной системе сформировались необходимы нервные связи, которые разгрузили ваш мозг, а ваше тело приобрело соответствующие навыки.
На самом деле один из смыслов тренировок при занятии каким-либо спортом состоит как раз в формировании необходимых навыков, то бишь в создании и последующей оптимизации связей между нейронами, которые обеспечивают наиболее оптимальные движения для данного вида спорта. То, что обычно называется спортивной техникой. При этом чем раньше человек начинает заниматься тем или иным видом спорта, тем легче его нервной системе сформировать необходимые связи, поскольку она ещё не наполнена программами, как у взрослого человека. Именно поэтому сейчас наблюдается тенденция, что чем раньше ребёнок начинает заниматься тем или иным видом спорта, тем больше у него шансов добиться выдающихся результатов. К этому нужно также добавить, что при занятиях той или иной деятельностью нервная система будет не только перестраивать свои нервные связи, но также будет запускать процессы адаптации всего организма к данным условиям.
Процесс формирования связей и оптимизации структуры нейронной сети происходит не только для выполнения движений, но и вообще для любой деятельности, которую осуществляет нервная система и наш мозг. Если вы занимаетесь математикой и решаете много задач, то у вас также будет вырабатываться соответствующие навыки, ваша нейронная сеть перестроится и с какого-то времени вы будете решать задачи быстрее других. Часто вы даже будете знать ответ только посмотрев на условие задачи, ещё толком не успев его аналитически обосновать (это проверено мной на личном опыте). Аналогично формирование навыков, то есть необходимых связей в нейронной сети, происходит и при занятии музыкой, и при обучении рисованию, и вообще при любой деятельности. Обучаясь чему либо, мы постоянно программируем сами себя, меняя связи между нейронами.
Если проводить аналогию с современными компьютерами, то в начале мы решаем какую-либо задачу программно, используя ресурсы головного мозга, а если та или иная задача повторяется достаточно часто, то соответствующая программа переносится на аппаратный уровень, что резко сокращает время её выполнения.
При этом перестройка связей между нейронами происходит не в любой момент времени. Поскольку процесс этот весьма не быстрый, для того, чтобы перестраивать связи между нейронами, нам необходим регулярный сон. И именно в этом и состоит главная функция сна, о которой вы не прочитаете ни в одном учебнике или книге по медицине!
Та информация, которую наш мозг воспринимает во время бодрствования, принимается и сохраняется в виде набора электрических импульсов, которые распространяются в среде нейронов головного мозга. Это, если там можно выразиться, наша оперативная память. И хотя количество нейронов в головном мозге очень большое, наша оперативная память всё равно оказывается достаточно ограниченной и её необходимо периодически очищать. Именно этот процесс на самом деле и происходит во время сна. Бытует ошибочное мнение, что есть две фазы сна, медленная и быстрая. Это не совсем так. Согласно последним исследованиям различают четыре фазы медленного сна и одну фазу так называемого быстрого сна. Названия «медленная» и «быстрая» данные фазы получили из-за частоты основных мозговых волн, которые фиксируются у коры головного мозга во время той или иной фазы сна.
Общая суть происходящих во время сна процессов следующая. После засыпания происходит первичный анализ накопленной за день информации, во время которого принимается решение, какую информацию нужно сохранить надолго, какую нужно оставить на некоторое время, а какую можно забыть, как несущественную. Та информация, которую мы решили сохранить на некоторое время, так и останется в «оперативной памяти», то бишь в виде набора импульсов, которые распространяются между нейронами. Информация, которую решено забыть, просто стирается, а соответствующие нейроны освобождаются и переходят в режим ожидания. А с той информацией, которую было решено сохранить в долговременную память как важную, начинается дальнейшая работа.
На следующей фазе происходит составление плана перестройки связей между нейронами для того, чтобы запомнить необходимую информацию или навыки. При этом если информация запоминается в коре головного мозга, то навыки переносятся на уровень спинного мозга или даже периферической нервной системы, где и будут формироваться новые связи между нейронами. Когда программа перестройки готова, то наступает так называемая «четвёртая фаза» или глубокий медленный дельта-сон. Именно в этот момент одни связи между нейронами разрушаются, а другие формируются. То есть, ставшие ненужными или содержащие ошибки программы могут быть стёрты или скорректированы, а необходимые новые будут дополнительно добавлены.
Именно тем, что во время этой фазы нейронная сеть находится в состоянии глубокой перестройки связей, объясняется тот факт, что во время дельта-сна человека очень сложно разбудить. А если это удаётся, то он будет чувствовать себя плохо, не выспавшимся, рассеянным, с пониженными показателями мозговой деятельности. При этом чтобы прийти в нормальное состояние ему нужно ещё поспать от пяти до пятнадцати минут. После этого он уже просыпается окончательно и при этом чувствует себя очень бодрым и выспавшимся. Почему? Да потому, что когда его разбудили, часть связей была ещё не сформирована, поэтому нейронная сеть не могла функционировать нормально. А когда он ещё немного поспал, то процесс формирования связей завершился и нервная система смогла переключиться в нормальный режим работы.
Подобные циклы анализа, формирования программы перестройки связей и собственно их перестройка за время сна повторяются циклически 4-5 раз. Соответственно, человека сравнительно легко и без особых последствий для него можно разбудить во время стадии анализа и подготовки программы, но нежелательно будить во время фазы перестройки связей.
А вот быстрая фаза сна служит уже для других целей. Именно во время этой фазы мы видим наиболее яркие и красочные сны. Данная фаза нужна для анализа накопленной информации или решения тех задач, на которые нам не хватает ресурсов во время бодрствования, в том числе для моделирования различных ситуаций, включая предсказание возможного развития событий в будущем. Именно по этому у нас на Руси есть поговорка: «утро вечера мудренее».
Дело в том, что во время бодрствования большая часть ресурсов нервной системы расходуется на обработку сигналов, поступающих от наших органов чувств. До 80% мы расходуем только на анализ зрительной информации. Именно поэтому многие люди, когда заняты решением сложной задачи, обдумыванием какой-то важной проблемы или пытаются вспомнить нужную им информацию, прикрывают на время глаза. Это позволяет им часть ресурсов нервной системы направить на решение этой задачи. Во время сна наши органы чувств находятся в пассивном состоянии, реагируя только на самые сильные раздражители, что позволяет высвободить основную часть мозга для анализа имеющейся информации и решения важных для нас задач. Именно поэтому там много рассказов о «вещих снах» и о том, что именно во сне человек вспомнил, куда он положил ту вещь, которую никак не мог найти днём, либо о том, что во сне ему удалось наконец-то решить ту или иную задачу, над которой он безуспешно бился днём. Одна из самых известных историй на эту тему о том, как Дмитрий Иванович Менделеев именно во сне увидел как должна выглядеть периодическая система химических элементов (и которую, кстати, нам сегодня рисуют совсем в другом искажённом виде).
В вещих снах, в которых человек видит некие события, которые потом происходят на яву, на самом деле тоже нет никакой мистики. То, что будущее можно в определённых пределах предсказать, на самом деле является очевидным фактом. Практически каждый, кто водит автомобиль, вынужден постоянно предугадывать будущее на основании той информации об окружающем мире, которую он воспринимает через свои органы чувств, а также своего предыдущего опыта, который им накоплен и сохранён в виде нейронных связей в коре его головного мозга. Невозможно двигаться на автомобиле, не попадая при этом в аварии, если ты не можешь предугадать, что должно произойти на дороге в ближайший момент времени. Появится ли на перекрестке другой автомобиль поперёк вашего пути или нет? Ведь от момента нажатия на педаль и до того, как ваш автомобиль проедет перекрёсток, проходит довольно много времени. То есть, при подъезде к перекрёстку ваш мозг через органы чувств, в первую очередь зрение, собирает информацию о поведении окружающих объектов, анализирует её и предсказывает будущее, то есть то, где они окажутся в тот момент, когда ваш автомобиль через несколько секунд будет находиться на перекрёстке.
Если ваш мозг ошибся или получил неполную информацию, то прогноз будет ошибочным, что может привести к аварии или только к аварийной ситуации, если предсказание мозга водителя другого автомобиля окажутся лучше ваших, поскольку он был более внимателен или более опытен, что позволило ему избежать столкновения. И то, что во время управления автомобилем водителя ничто не должно отвлекать, в том числе разговор по сотовому телефону, объясняется как раз тем, что любой дополнительный мыслительный процесс так или иначе забирает на себя часть ресурсов мозга, а это значит, что он начинает хуже воспринимать поступающую информацию или делает менее качественные предсказания будущего.
Также мы регулярно делаем предсказания и на более длительный срок, хотя и более простые, которые часто называем «планирование». Если вы всё хорошо спланировали и учли все факторы, которые могут повлиять на результат, то с очень большой вероятностью запланированное событие происходит.
На самом деле в вещих снах нет ничего удивительного. Мы постоянно получаем информацию об окружающем нас мире, в том числе такую, которую днём просто не успеваем до конца проанализировать. Но во сне, когда основная часть ресурсов мозга как раз направлена на анализ собранной информации, наше сознание может сделать глубокий качественный анализ и сформировать более качественное предсказание, которое мы и увидим во сне как «вещее».
Но видим сны, тем более вещие, мы далеко не всегда. Фаза быстрого сна наступает только после как минимум одного полного цикла медленного сна. Для того, чтобы мозг мог приступить к анализу собранной информации и формированию сновидений, он должен хотя бы частично освободиться от накопленной за день информации. При этом экспериментально установлено, что чем дальше, тем продолжительность фазы быстрого сна становится больше. И это совершенно логично, поскольку чем больше циклов переноса информации с оперативной в долговременную память успело пройти, тем больше ресурсов освободилось у мозга для обработки информации и формирования сновидений. Но если вы постоянно не высыпаетесь, то ваш мозг постепенно переполняется, не успевая полностью очиститься во время слишком короткого сна. В этом случае у вас либо вообще не будет фаз быстрого сна, либо они будут очень короткими, при этом вы не будете запоминать те сновидения, которые в это время будут возникать, поскольку ваша память ещё не освободилась от накопленной информации. Другими словами, если вы не видите или не можете вспомнить свои сны, то это означает, что вы спите не достаточно и ваш мозг не успевает восстановиться.
Представьте, что мозг это сосуд, а поступающая за день информация, это вода, которую мы в этот сосуд постепенно наливаем. Переработка во время сна накопленной за день информации аналогична опустошению этого сосуда от накопленной за день воды. Ну а дальше у нас получается известная нам ещё со школы задачка про то, сколько в сосуд воды втекает, и сколько вытекает. Если общая емкость сосуда 5 литров и вы каждый день будете вливать по 1.5 литра воды, а выливаться за время короткого сна будет только 1 литр, то каждый день у вас будет оставаться по 0.5 литра воды. Соответственно, на восьмой день ваш сосуд окажется заполненным на 4 литра и очередных полтора литра воды вы в него просто не сможете влить. Остальная вода просто не поместится в сосуд, а будет проливаться мимо него. И если ничего не менять, то этот процесс переполнения может продолжаться достаточно долго. До тех пор, пока вы не увеличите время слива воды, слив всю лишнюю накопившуюся воду, то бишь не выспитесь как следует, дав возможность вашему мозгу разгрести наконец Авгиевы конюшни излишков накопленной информации.

Считается, что человеку для сна требуется около 8 часов. Это цифра весьма примерная, поскольку на практике это зависит от характера деятельности, которой занят человек в течение дня. Если эта деятельность связана с однообразной физической деятельностью, при которой накопление информации идёт медленнее, то для сна может требоваться меньше времени. Если же человек занят активной умственной деятельностью, то ему может потребоваться и больше 8 часов. Но если вы регулярно не высыпаетесь, то ваши интеллектуальные возможности начнут постепенно ухудшаться. Вам будет сложнее воспринимать и запоминать информацию, вы будете хуже решать задачи, ваше внимание будет более рассеянным.
Вообще без сна средний человек может находится 3-4 дня. Рекорд максимального пребывания без сна, без использования стимуляторов любого рода, был установлен в 1965 году американским школьником Рэнди Гарднером из города Сан-Диего, штат Калифорния, который бодрствовал в течении 264.3 часов (одиннадцать дней). При этом в некоторых источниках даже говорится о том, что продолжительное лишение сна имеет весьма небольшой эффект. Но если поднять более подробный отчёт о данном эксперименте, то выясняется, что это далеко не так. Подполковник Джон Росс, который контролировал состояние здоровья Гарднера, сообщил о серьёзном изменении умственных способностей и поведения в период лишения сна, включающее в себя удручённость, проблемы с концентрацией и краткосрочной памятью, паранойю и галлюцинации. На четвёртый день Гарднер представлял себя Полем Лоуи, играющим на стадионе Rose Bowl, и принимал уличный знак за человека. В последний день, когда его попросили последовательно вычитать 7 из 100-а, он остановился на 65-ти. Когда его спросили, почему он остановил счёт, он заявил, что забыл что сейчас делает.
Таким образом одна из полезных рекомендаций, которую можно дать в свете вышеизложенной информации, состоит в том, что если вы не можете по каким-то причинам постоянно спать необходимое вам время, то желательно хотя бы раз в неделю как следует выспаться, чтобы дать вашему организму время компенсировать тот недостаток сна, который у вас накопился. При этом показателем того, что сна достаточно, будет не подъём по будильнику, а пробуждение тогда, когда это произойдёт естественным путём и вы почувствуете, что наконец-то выспались. Если для этого потребуется проспать 12 часов, значит нужно спать 12 часов.
Но для нормального восстановления ресурсов мозга во время сна необходимо не только время, но и энергия. Наш мозг потребляет очень много энергии. Составляя всего 5% массы тела, в зависимости от рода деятельности мозг потребляет от 30% до 50% получаемой организмом энергии. При этом большую часть энергии мозг получает за счёт процесса катаболизма глюкозы, то есть медленного окисления глюкозы до СО2 и Н20 (углекислый газ и вода). Глюкозу мы получаем с пищей, которая током крови переносится к клеткам головного мозга. Но одной глюкозы для этого процесса недостаточно, для окисления каждой молекулы глюкозы С6Н12О6 необходимо ещё 6 молекул кислорода О2, который мы постоянно получаем из окружающего воздуха во время дыхания. Это означает, что если вы хотите хорошо высыпаться или занимаетесь активной умственной деятельностью, помещение, где вы находитесь, должно достаточно хорошо проветриваться. В противном случае, если в воздухе имеется недостаток кислорода или, что бывает намного чаще, избыток углекислого газа, ваш мозг будет получать недостаточно энергии для всех протекающих в нём процессов. Так что даже если вы проспите 8 или даже 10 часов в плохо проветриваемом помещении, этого будет недостаточно, чтобы хорошо выспаться, что многократно проверено мной на личном опыте. По этой же причине рекомендуется обеспечить вентиляцию помещения, где вы занимаетесь активной умственной деятельностью, в том числе где проходит обучение. Наверное многие из вас замечали, что когда много людей собираются в небольшом помещении, например, чтобы послушать какой нибудь доклад или лекцию, то через некоторое время людей начинает клонить в сон. Это происходит как раз потому, что из-за скопления большого количества людей в помещении резко повысилась концентрация углекислого газа и что уменьшает приток кислорода в кровь и наш мозг переходит в энергосберегающий режим, понижая свою активность и переставая воспринимать информацию, особенно если лекция скучная. То есть, делает примерно тоже самое, что и процессор ноутбука, который замедляет свою работу при переходе на питание от аккумулятора. И чтобы сохранять внимание, нам нужно в подобной ситуации будет прикладывать дополнительные усилия, не давая себе заснуть.
В свете повальной моды на установку пластиковых окон, которые, несомненно, гораздо лучше изолируют помещение от улицы, проблема вентиляции помещений становится ещё более актуальной, поскольку существующая в зданиях система естественной вентиляции справляется далеко не всегда, а зачастую и вообще не работает, поскольку соседи выше этажом во время очередного евроремонта умудрились засыпать мусором ваш вентиляционный канал. Так что если вы хотите хорошо высыпаться, особенно если времени для сна у вас оказывается недостаточно, то уделите особое внимание тому, чтобы ваше помещение для сна хорошо проветривалось. Лучше чуть приоткрыть ваше пластиковое окно, но при этом включит обогреватель, чем спать с плотно задраенными окнами в плохо проветриваемом помещении. По этой же причине в спальных помещениях желательно устанавливать пластиковые окна обязательно с системой микропроветривания, которые позволяют это окно чуть-чуть приоткрывать, либо приобрести и установить на ваше окно дополнительные внешние специальные приспособления, которые позволяют делать тоже самое, если у вас уже установлено такое окно без подобной системы.
Есть у сна и ещё одна важная функция, о которой большинство людей мало что знает. Последние исследования показали, что у людей с недостатком сна наблюдается не только снижение качества работы мозга, но и понижение иммунитета. Происходит это потому, что именно во время сна запускаются процессы регенерации и восстановления повреждённых тканей, а также формирования необходимых антител для борьбы с вирусами и бактериями. Во всех этих процессах участвуют спинной мозг и периферическая нервная системы. Во время бодрствования они загружены обеспечением двигательной активности человека, а во время сна их ресурсы высвобождаются и могут быть использованы для анализа что, где и как следует в организме починить. Именно поэтому когда мы болеем, нам хочется прилечь и поспать. По этой же причине, если вы не достаточно высыпаетесь, то болеть вы будете чаще, а организм ваш будет стареть и разрушаться быстрее.
Отдельная тема, это использование различных нейростимуляторов, особенно всяческих энергетиков, которые, как уверяет реклама, позволяют сократить время сна и оставаться бодрым и весёлым длительное время. На коротких промежутках времени это действительно так. С помощью химического воздействия вы можете заставить свой мозг активно работать ещё несколько часов. Но при этом нужно понимать, что это происходит далеко не бесплатно.
Во-первых, применение нейростимуляторов, будь то чай, кофе, или более агрессивные энергетики, на самом деле не увеличивает емкости вашего мозга, его оперативной памяти, того гипотетического сосуда, в который мы можем наливать воду поступающей из окружающего мира информации. Они лишь позволяют вам вместо 1.5 литров за один раз налить туда 2 литра. Но это означает, что ваш сосуд переполниться гораздо быстрее. Поэтому критическое состояние переполнения, после которого мозг уже перестаёт нормально функционировать, наступает гораздо быстрее, после чего уже никакие нейростимуляторы вам толком помогать не будут. Соответственно, после подобного экстремального режима работы вашему мозгу потребуется более длительный отдых (нужно слить больше воды).
Во-вторых, все нейростимуляторы переводят нейроны в предельный или даже запредельный режим работы, что резко понижает срок их жизни. Весьма популярный миф о том, что нейроны в организме не восстанавливаются, давно уже опровергнут. Возник он потому, что нейроны являются наиболее долгоживущими клетками в организме, потому что заменять их в составе нейронной сети весьма не простая задача, поэтому организм пытается оттянуть этот процесс на как можно более поздний срок. По этой же причине новые нейроны появляются намного медленнее обычных клеток. Так что в данном случае вопрос не в том, что новые нейроны в организме вообще не появляются, а в балансе гибели существующих и появлении новых нервных клеток. Если нейроны гибнут быстрее, чем организм производит новые, то происходит процесс деградации нервной системы и сознания. И если вы начинаете злоупотреблять теми же энергетиками, то тем самым вы повышаете скорость гибели нейронов, делая этот баланс отрицательным.
Аналогичный, но гораздо более сильный эффект происходит и при употреблении различных наркотических веществ, в особенности алкоголя. О том, как воздействует на организм и нервную систему алкоголь, я расскажу в следующей части.
Продолжение http://mylnikovdm.livejournal.com/6080.html
Аксон – это волокнистая ось, отходящая от тела нейрона, покрытая миелиновым слоем, обеспечивающая связь с другими нейронами и клетками рабочих органов. Представляет собой удлиненный осевой отросток, по которому передаются потенциалы действия (возбуждения), что делает его важнейшим структурным элементом ЦНС.
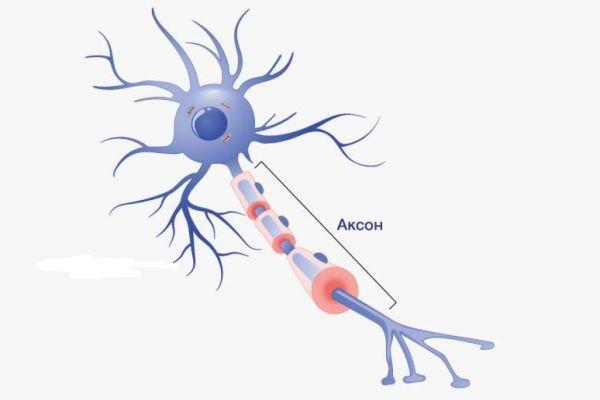
Определение
Мозговое вещество – высокоорганизованная структура, образованная нервными клетками, от которых отходят аксоны. Из нервных клеток состоит мозговая ткань. Аксон в переводе с греческого означает «ось» – это такой отросток, элемент мозгового вещества, который обеспечивает взаимодействие между клетками разного типа (нейроны, клетки иннервируемых органов), что ассоциируется с тонким, четким управлением работой органов и систем. Функции ткани ЦНС:
- Воспринимает раздражения, преобразуя их в импульсы.
- Поддерживает передачу импульсов от управляющих отделов мозга к исполнительным органам.
- Формирует ответную реакцию на раздражающее воздействие.
- Обеспечивает взаимодействие в работе систем и органов, поддерживает интеграцию структурных единиц организма.
- Обеспечивает взаимосвязь организма с внешней средой.
Согласно определению в биологии, аксон (англ. axon) – удлиненный отросток, по которому идут импульсы от тела нейрона к другим нервным клеткам и структурным элементам всех тканей организма. Мозговая ткань в период внутриутробного развития образуется из нервной пластины. Края пластинки прогибаются, что приводит к формированию валиков и желобка. В результате смыкания краев валиков возникает нервная трубка – основа ЦНС.
Дифференциация клеток, образующих трубку, приводит к появлению нейробластов и спонгиобластов. Первые служат основой для формирования нейронов, вторые – для образования нейроглии. Нейроны (анат.) – основные структурные элементы мозгового вещества. Они характеризуются отсутствием функции деления, что приводит к постепенному уменьшению их численности. Тело нейрона состоит из ядра и цитоплазмы. В зависимости от типа нейронов меняется геометрическая форма тела, которая бывает круглая, овальная, пирамидальная и другая.
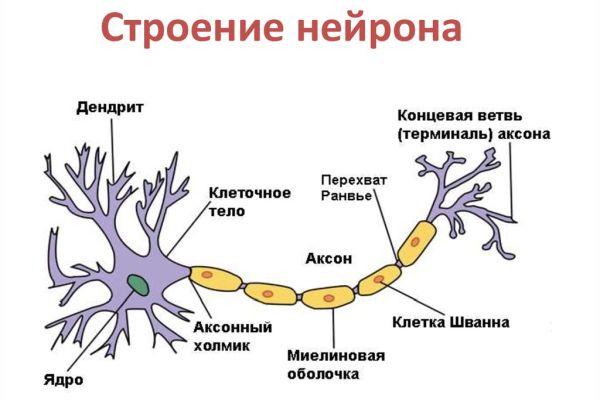
Цитоскелет, состоящий из микротрубочек и нейрофибриллов, обеспечивает опорную и трофическую функцию. Цитоскелет поддерживает форму нейрона, обеспечивает транспорт веществ и органелл. От тела ответвляются отростки – единичный аксон и множественные дендриты. Аксон нейрона почти не ветвится, иногда образует коллатеральные (обходные) сегменты. Концевые сегменты (окончания) разветвляются, называются терминали.
Терминали взаимосвязаны с окончаниями других нейронов и с клетками, образующими паренхиму (ткань) рабочих органов – мышц, желез. Количество дендритов варьируется от 1 до нескольких. Тонкие ответвления дендритов оканчиваются небольшими шипами, где сосредоточены терминали аксональных отростков многих тысяч других клеток. Дендриты воспринимают раздражения или потенциалы действия от других клеток и передают их по волокнам к телу своего нейрона.
Рост аксона зависит от особенностей строения и жизнедеятельности нейрона, который поддерживает функцию питания отростка. К примеру, если перерезать аксональный ствол, сегмент, связанный с телом, остается жизнеспособным и продолжает деятельность, участок, утративший связь с телом, отмирает. Аксоны образуют нервы, что предполагает сложную структурно-морфологическую организацию ЦНС.
Строение
Аксон – это длинный отросток нейрона, который обеспечивает взаимодействие между нервными клетками. Согласно анатомии, аксон ответвляется от холмика, находящегося на теле. Холмик аксона представляет собой структуру, где постсинаптический потенциал преобразуется в биоэлектрический сигнал. Чтобы в холмике происходила генерация биоэлектрических сигналов, необходима согласованная деятельность каналов –натриевых, кальциевых, нескольких типов калиевых.
Длина аксона у человека существенно варьируется в зависимости от вида нейрона, от которого отходит аксональная ось. Минимальная длина – около 1 миллиметра, максимальная – около 1,5 метров. Длина более 1 метра наблюдается в случаях, когда отросток отходит от спинного мозга в область конечностей. Диаметр аксональной оси также неодинаковый у разных типов клеток, равен около 1-20 микрон. Импульсы проходят быстрее по аксональным осям большего диаметра.
Размеры аксонального отростка нередко достигают 99% от общего объема нервной клетки, в структуру которой он входит. Аксон состоит из протоплазмы (аксоплазмы), где находятся тончайшие волокна, белковые нити – нейрофибриллы, из чего образован ствол аксонального ответвления. Согласно одной из теорий, нейрофибриллы – проводники питательных веществ. Аксональная протоплазма также содержит митохондрии и микротрубочки, которые представляют собой самые крупные элементы цитоскелета.
Диаметр микротрубочек составляет около 24 нанометров. Они обеспечивают внутриклеточный транспорт веществ, в том числе поддерживают трофику аксональных отростков. Тело (перикарион) – источник протеинов и нейромедиаторов, распространяющихся по аксональной оси посредством микротрубочек, которые у аксона имеют направленную полярную ориентацию (в отличие от микротрубочек дендритов).
Положительно заряженные концы микротрубочек направлены к сегменту терминали, отрицательно заряженные концы – к телу. Строение аксона предполагает наличие оболочки. Аксон покрыт глиальным (миелиновым) слоем по всей длине, чем защищен от разрушающих внешних воздействий. Миелиновый слой в аксональных отростках периферического отдела сформирован клетками Шванна.
Миелиновая оболочка, покрывающая нервную ось, обеспечивает ее механическую прочность, электрохимическую изоляцию, трофику (питание). Миелиновый слой ускоряет проведение биоэлектрических сигналов. Нервы – пучки объединенных аксональных отростков, которые покрыты оболочкой из соединительной ткани и снабжены кровеносными сосудами.
Функции
Основная задача нейронов – переработка данных. С их помощью осуществляется получение, обработка, передача информации отделам нервной и других систем организма.
Если дендриты проводят сигналы по направлению к телу нервной клетки (перикариону), то аксональный отросток передает импульсы от перикариона к другим клеткам.
Основная функция аксонов – проведение импульсов в пределах нейрональной сети и к исполнительным органам. Аксональные ответвления относятся к первичным проводниковым путям в нервной системе. Вспомогательная функция – транспорт веществ. При помощи аксонального транспорта осуществляется движение белков, синтезированных в теле, нейромедиаторов, органелл. Многие вещества способны двигаться в обоих направлениях.

В периферических сегментах аксона в него могут проникать вирусы и токсичные вещества, которые перемещаясь к телу нервной клетки, повреждают ее. Аксональный транспорт зависит от количества энергии АТФ. Если энергетический уровень АТФ понижается больше, чем в 2 раза, происходит блокировка аксонального транспорта.
Функции аксона заключаются в передаче импульсов. При взаимодействии аксона с телом другого нейрона образуется аксосоматический контакт. Если аксон взаимодействует с дендритами других клеток возникает аксодендритический контакт. Взаимодействие с аксоном другой клетки приводит к образованию аксо-аксонального контакта, который редко происходит в нервной системе, поддерживает тормозные рефлекторные реакции.
Особенности регенерации нервной ткани
Нервные клетки почти полностью лишены способности к регенерации. Однако нервные клетки способны восстанавливать поврежденные или утраченные ответвления. Процесс регенерации аксона возможен, если тело сохраняет жизнеспособность, и на пути роста аксонального отростка отсутствуют препятствия. В ходе процесса регенерации отросток вновь прорастает к органу-мишени.
Восстановление нервной проводимости в мышцах с нарушенной иннервацией – один из критериев успешного лечения невропатий разного генеза. При невропатиях травматического генеза восстановление функций мышц происходит за счет регенерации ствола прерванного аксона и ремиелинизации отростка (восстановление миелиновой оболочки). Периферический отдел нервной системы обладает более высоким потенциалом регенерационных возможностей в сравнении с центральным отделом.
Восстановление иннервации в мышечной или кожной ткани происходит благодаря сохранившимся аксонам, которые начинают ускоренно разрастаться и ветвиться. Процесс ветвления аксонов в зоне перехватов Ранвье (периодические разрывы миелинового слоя) получил название «спрутинг». В результате происходит частичное или полное возобновление первичной иннервации.
В ходе экспериментов установлено, что близлежащие интактные (не вовлеченные в патологический процесс) аксоны выпускают нервные волокна, которые иннервируют участок мускулатуры или кожных покровов с нарушенной проводимостью нервных импульсов. Различают виды спрутинга – коллатеральный (обходной) и регенераторный (терминальный).
Регенераторный спрутинг начинается после устранения в нейронах ретроградных изменений, обусловленных аксонотомией (повреждением, рассечением нервной оси). Это связано с потребностью в продукции аппарата ядра нервной клетки, производящего протеины. Материал, необходимый для регенераторного спрутинга, продуцируется в теле и транспортируется по микротрубочкам по всей длине оси. Параллельно происходит процесс ремиелинизации осевого ствола.
Аксон – удлиненный отросток нервной клетки, обеспечивающий взаимодействие между структурными элементами мозговой ткани и связь ЦНС с исполнительными органами.
Просмотров: 2 334

Содержание
- Обзор
- Анатомия
- Начальный сегмент
- Узлы Ранвье
- Потенциалы действия
- Рост и развитие
- История
- Травма, повреждение
- Классификация
- Мотор
- Сенсорный
- Автономный
| Аксон |
|---|
 Узел ядра аксона дендрита сомы Узел ядра аксона дендрита сомыRanvier Axon Terminal Шванновская клетка Миелиновая оболочка |
| Структура типичного нейрона |
An аксон представляет собой тонкий, похожий на руку (или похожий на кабель) выступ, который выходит из тела нейрона (нервной клетки) и проводит нервные импульсы по всей его длине. Обычно, но не всегда, аксоны проводят нервные импульсы от тела клетки, вызывая на их конечном конце выброс нейротрансмиттеров во внеклеточное пространство, где они могут возбуждать или подавлять другие нейроны. В некоторых сенсорных нейронах нервные импульсы проходят по аксону от периферии к телу клетки.
Во многих случаях аксон нейрона может быть очень длинным и поэтому известен как нервное волокно. У жирафов есть одиночные аксоны длиной в несколько метров, проходящие по всей длине шеи, а человеческий мотонейрон может быть более метра в длину, простираясь от поясничной области позвоночника до пальцев ног. Однако у некоторых нейронов аксоны очень короткие и даже отсутствуют. Хотя нейрон имеет не более одного аксона, некоторые аксоны могут иметь ветви, и такие ветви могут быть значительными ближе к концу длины аксона, в том числе с 10 000 или более концевыми ветвями.
Аксон — это один из двух типов отростков, которые исходят от тела нейронной клетки, другой — дендриты. Дендриты представляют собой разветвленные (не похожие на руки) выступы, которые обычно получают сигналы от других нейронов и передают сигналы к телу клетки, обычно используя потенциалы действия на короткие расстояния, а не потенциалы действия (нервные импульсы) аксонов. Аксоны имеют большинство таких же органелл, как дендриты и тело клетки, но не имеют аппарата Гольджи и тельцов Ниссля.
Аксоны — это основные линии передачи нервной системы. Координация между многими сложными частями и процессами аксона — узлами Ранвье, потенциалом действия «все или ничего», ионными каналами кальция, пузырьками, заполненными нейротрансмиттером, рецепторами и т. Д. — отражает удивительную гармонию в природе.
Обзор
Аксон — это проекция нейрон. Нейрон или нервная клетка — это высокоспециализированная электрически возбудимая клетка нервной системы, которая проводит нервные импульсы между различными частями тела. Нейроны могут обрабатывать и передавать информацию как из внутренней, так и из внешней среды, передавая эту информацию посредством химического или электронного импульса через синапс (соединение между клетками) и используя потенциал действия — электрический сигнал, который генерируется посредством электрически возбудимой мембраны нейрон. У позвоночных животных нейроны являются основными компонентами головного, спинного мозга и периферических нервов.
Три основных типа нейронов: сенсорные нейроны (у которых есть специализированные рецепторы для преобразования разнообразных стимулов из окружающей среды в электрические сигналы, а затем передачи этой информации в более центральные точки нервной системы, такие как спинной или головной мозг); двигательные нейроны (которые передают импульсы от центральной области нервной системы к эффектору, например, к мышце); а также интернейроны или ретрансляционные нейроны (которые преобразуют химическую информацию обратно в электрические сигналы).
Три основных структурных области типичного нейрона: A сома, или тело клетки, которое содержит ядро; одно или несколько дендритных деревьев, которые обычно получают входные данные; и аксон несущий электрический импульс. Также можно выделить из аксона область, обозначенную как аксонный терминал, который относится к небольшим ветвям аксона, которые образуют синапсы или соединения с другими клетками и часто служат для передачи сигналов другим клеткам.
В сома или перикарион — луковичный конец нейрона, от которого отходят дендриты и аксон. Сома содержит множество органелл, гранул, называемых гранулами Ниссля, и ее ключевой особенностью является наличие ядра клетки.

Дендриты являются одним из двух типов протоплазматических выступов, которые выступают из тела клетки нейрона. Это клеточные отростки с множеством ответвлений, и именно здесь происходит большая часть ввода в нейрон. Общая форма и структура дендритов нейрона называется его дендритным деревом. Большинство нейронов имеют несколько дендритов, которые выходят наружу от сомы и специализируются на получении химических сигналов от концов аксонов других нейронов. Дендриты преобразуют эти сигналы в небольшие электрические импульсы и передают их соме.
Аксоны являются вторым из двух типов протоплазматических выступов, отходящих от клеточных тел нейронов. Аксон представляет собой тонкий, похожий на кабель выступ, который может увеличиваться в длину в десятки, сотни или даже десятки тысяч раз больше диаметра сомы и обычно проводит электрические импульсы от тела клетки нейрона. Функция аксона — передавать информацию различным нейронам, мышцам и железам. В некоторых сенсорных нейронах (псевдоуниполярных нейронах), таких как нейроны прикосновения и тепла, электрический импульс проходит по аксону от периферии к телу клетки и от тела клетки к спинному мозгу по другой ветви того же аксона.
Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно имеют постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты обычно получают сигналы, в то время как аксоны обычно их передают). Однако из всех этих правил есть исключения. Например, хотя аксон и бугорок аксона обычно участвуют в оттоке информации, эта область также может получать входные данные от других нейронов. Также может происходить отток информации от дендритов к другим нейронам. А аксоны могут быть очень короткими (или даже отсутствовать) в некоторых типах нейронов. Те типы нейронов, у которых отсутствует аксон, передают сигналы от своих дендритов. И дендриты, и аксоны имеют тенденцию разделять те же органеллы, что и сома, хотя у обоих отсутствует ядро, а у аксонов отсутствует аппарат Гольджи и тельца Ниссля.
Различие между дендритами и аксонами не всегда ясно. Например, нейроны, классифицируемые как униполярные (или псевдоуниполярные, поскольку они происходят как биполярные нейроны), имеют один отросток, который идет от тела клетки и образует два конца (центральный отросток и периферический отросток, оба с ветвями на концах, где есть сенсорные окончания / рецептивные окончания). В основном это сенсорные нейроны периферической нервной системы. Некоторые классифицируют это расширение как дендрит, используя старое определение дендритов как процессов, передающих импульсы к телу клетки. Однако функциональные определения, основанные на генерации и передаче импульса, классифицируют его как аксон (Marieb and Hoehn 2010).
Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга (Yau 1976).
Аксон специализируется на проведении электрического импульса, называемого потенциал действия, который движется от тела клетки вниз по аксону. Место соединения аксона и тела клетки называется аксонный бугорок («горка»). Это область нейрона, которая имеет наибольшую плотность зависимых от напряжения натриевых каналов, что делает ее наиболее легко возбуждаемой частью нейрона. Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсы. В синапсе мембрана аксона плотно прилегает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Большинство аксонов разветвляются, в некоторых случаях широко, обеспечивая связь со многими клетками-мишенями. Некоторые синаптические соединения появляются на полпути вдоль аксона по мере его расширения — это называется мимоходом («попутно») синапсы. Другие синапсы выглядят как терминалы на концах аксональных ветвей. Один аксон со всеми его ветвями, вместе взятыми, может иннервировать несколько частей мозга и генерировать тысячи синаптических окончаний.
Анатомия

Аксоны — это первичные линии передачи нервной системы, и в виде пучков они образуют нервы. Некоторые аксоны могут достигать одного метра и более, а другие — всего лишь один миллиметр. Самые длинные аксоны в человеческом теле — это аксоны седалищного нерва, которые проходят от основания спинного мозга до большого пальца каждой ноги. Диаметр аксонов также варьируется. Большинство отдельных аксонов имеют микроскопический диаметр (обычно около 1 микрона в диаметре). Самые большие аксоны млекопитающих могут достигать в диаметре до 20 микрон. Гигантский аксон кальмара, который специализируется на очень быстрой передаче сигналов, имеет диаметр около 1 миллиметра, размер небольшого грифеля карандаша. Аксональное ветвление (разветвленная структура на конце нервного волокна) также отличается от одного нервного волокна к другому. Аксоны в центральной нервной системе обычно представляют собой сложные деревья с множеством точек ветвления. Для сравнения, аксон гранулярных клеток мозжечка характеризуется одним Т-образным узлом ответвления, от которого отходят два параллельных волокна. Продуманная ветвь позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
Есть два типа аксонов, встречающихся в периферической системе и центральной нервной системе: немиелинизированные и миелинизированные аксоны. Миелин представляет собой слой изолирующего жирового вещества, а миелиновые оболочки вокруг аксонов защищают и электрически изолируют аксон (Marieb and Hoehn 2010). Миелин образован двумя типами глиальных клеток: шванновскими клетками, покрывающими периферические нейроны, и олигодендроцитами, изолирующими нейроны центральной нервной системы. Вдоль миелинизированных нервных волокон через равные промежутки времени возникают промежутки в миелиновой оболочке, известные как узлы Ранвье. Миелинизация аксонов (миелинизированные волокна с миелиновой оболочкой) обеспечивает особенно быстрый режим распространения электрических импульсов, называемый скачкообразной проводимостью. Немиелинизированные волокна передают нервные импульсы довольно медленно (Marieb and Hoehn, 2010). Демиелинизация аксонов вызывает множество неврологических симптомов, обнаруживаемых при заболевании «Рассеянный склероз».

Если головной или спинной мозг позвоночного животного извлекается и разрезается на тонкие срезы, некоторые части каждой части кажутся темными, а другие — более светлыми. Темные части известны как серое вещество, а более светлые части — как белое вещество. Белое вещество получает свой светлый цвет из миелиновых оболочек аксонов: части белого вещества мозга характеризуются высокой плотностью миелинизированных аксонов, проходящих через них, и низкой плотностью клеточных тел нейронов. Белое вещество спинного и головного мозга не содержит дендритов, которые встречаются только в сером веществе. Серое вещество содержит дендриты, тела нервных клеток и более короткие немилинированные аксоны. Кора головного мозга имеет на поверхности толстый слой серого вещества; под ним находится большой объем белого вещества: это означает, что большая часть поверхности заполнена телами нейронных клеток, тогда как большая часть области внизу заполнена миелинизированными аксонами, которые соединяют эти нейроны друг с другом. Как правило, белое вещество можно понимать как части головного и спинного мозга, ответственные за передачу информации (аксоны); тогда как серое вещество в основном отвечает за обработку информации (тела нейронов). В спинном мозге человека аксоны, покрытые миелином, находятся на поверхности, а сети аксон-дендриты находятся внутри, тогда как в головном мозге это наоборот (т. Е. В спинном мозге белое вещество находится снаружи, в то время как оно находится преимущественно внутри мозга (Чемберлин и Наринс 2005; Кэмпбелл и др. 2008; Мариеб и Хоэн 2010).
Начальный сегмент
Начальный сегмент аксона — толстая немиелинизированная часть аксона, которая соединяется непосредственно с телом клетки — состоит из специализированного комплекса белков. Его длина составляет примерно 25 мкм, и он функционирует как место инициации потенциала действия (Clark et al. 2009). Плотность потенциалзависимых натриевых каналов намного выше в начальном сегменте, чем в остальной части аксона или в прилегающем теле клетки, за исключением бугорка аксона (Wollner and Catterall 1986).
Как известно, потенциалзависимые ионные каналы находятся в определенных областях аксональной мембраны и инициируют потенциал действия, проводимость и синаптическую передачу (Debanne et al. 2011).
Узлы Ранвье
Узлы Ранвье (также известные как щели миелиновой оболочки) представляют собой короткие немиелинизированные сегменты миелинизированного аксона, которые периодически встречаются между сегментами миелиновой оболочки. Следовательно, в точке узла Ранвье аксон уменьшается в диаметре (Hess, Young 1952). Эти узлы представляют собой области, где могут быть созданы потенциалы действия. При скачкообразной проводимости электрические токи, возникающие в каждом узле Ранвье, передаются с небольшим затуханием к следующему узлу в линии, где они остаются достаточно сильными, чтобы генерировать другой потенциал действия. Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Потенциалы действия
| Структура типичного химического синапса |
|---|
 Постсинаптический ПостсинаптическийПлотность Напряжение- закрытый Ca++ канал Synaptic обратный захват пузырька помпа Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит |
Большинство аксонов несут сигналы в виде потенциалов действия, которые представляют собой дискретные электрохимические импульсы, которые быстро проходят по аксону, начиная с тела клетки и заканчиваясь в точках, где аксон устанавливает синаптический контакт с клетками-мишенями. Определяющей характеристикой потенциалов действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Когда потенциал действия достигает пресинаптического терминала, он активирует процесс синаптической передачи. Первый шаг — быстрое открытие каналов для ионов кальция в мембране аксона, позволяя ионам кальция проходить внутрь через мембрану. Возникающее в результате увеличение внутриклеточной концентрации кальция приводит к тому, что везикулы (крошечные контейнеры, окруженные липидной мембраной), заполненные химическим нейромедиатором, сливаются с мембраной аксона и выводят свое содержимое во внеклеточное пространство. Нейромедиатор высвобождается из пресинаптического нерва посредством экзоцитоза. Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или изменении ее метаболизма каким-либо образом. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор пузырьков перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона (Debanne et al. 2011).
Рост и развитие

Растущие аксоны перемещаются в окружающей среде через конус роста, который находится на кончике аксона. Конус роста имеет широкое пластинчатое расширение, называемое ламеллиподиями, которое содержит выступы, называемые филоподиями. Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует окружающую среду. Актин играет важную роль в подвижности этой системы.
Среда с высоким уровнем молекул клеточной адгезии или CAM создает идеальную среду для роста аксонов. Похоже, это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нервных систем, включают N-CAM, нейроглиальный CAM или NgCAM, TAG-1 и MAG, все из которых являются частью суперсемейства иммуноглобулинов. Другой набор молекул, называемых молекулами адгезии внеклеточного матрикса, также обеспечивает липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин, фибронектин, тенасцин и перлекан. Некоторые из них поверхностно связаны с клетками и, таким образом, действуют как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь эффекты дальнего действия.
Клетки, называемые направляющими клетками, помогают управлять ростом аксонов нейронов. Эти клетки обычно представляют собой другие, иногда незрелые нейроны.
В ходе исследований также было обнаружено, что если аксоны нейрона были повреждены, до тех пор, пока сома (тело клетки нейрона) не повреждена, аксоны будут регенерировать и воссоздавать синаптические связи с нейронами с помощью направляющего столба. клетки. Это также называется нейрорегенерацией (Kunik et al. 2011).
Nogo-A — это тип компонента, ингибирующего рост нейритов, который присутствует в миелиновых мембранах центральной нервной системы (обнаружен в аксоне). Он играет решающую роль в ограничении регенерации аксонов в центральной нервной системе взрослых млекопитающих. В недавних исследованиях, если Nogo-A блокирован и нейтрализован, можно вызвать регенерацию аксонов на большом расстоянии, что приводит к усилению функционального восстановления у крыс и спинного мозга мыши. Этого еще предстоит сделать на людях (Schwab 2004). Недавнее исследование также показало, что макрофаги, активируемые специфическим воспалительным путем, активируемым рецептором Dectin-1, способны способствовать восстановлению аксонов, однако также вызывают нейротоксичность в нейроне (Gensel et al. 2009).
История
Некоторые из первых внутриклеточных записей в нервной системе были сделаны в конце 1930-х годов К. Коулом и Х. Кертисом. Немецкому анатому Отто Фридриху Карлу Дейтерсу обычно приписывают открытие аксона, отличив его от дендритов (Debanne et al. 2011). Швейцарский Рюдольф Альберт фон Келликер и немец Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), и к 1952 году они получили полное количественное описание ионной основы потенциала действия, что привело к формулировке модели Ходжкина-Хаксли.
В 1963 году Ходжкин и Хаксли были совместно удостоены Нобелевской премии за эту работу. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхейзера-Хаксли. Луи-Антуан Ранвье был первым, кто описал промежутки или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлами Ранвье. Сантьяго Рамон-и-Кахаль, испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции (Debanne et al. 2011). Эрлангер и Гассер ранее разработали систему классификации периферических нервных волокон, основанную на скорости аксональной проводимости, миелинизации, размере волокна и т. Д. Даже недавно наше понимание биохимической основы распространения потенциала действия продвинулось и теперь включает много деталей об отдельных ионных каналах.
Травма, повреждение
По степени тяжести повреждение нерва может быть описано как нейропраксия, аксонотмезис или нейротмезис. Сотрясение мозга считается легкой формой диффузного повреждения аксонов (Dawodu 2013). Дисфункция аксонов в нервной системе является одной из основных причин многих наследственных неврологических расстройств, которые затрагивают как периферические, так и центральные нейроны (Debanne et al. 2011).
Классификация
Аксоны, из которых состоят нервы периферической нервной системы человека, можно классифицировать на основе их физических характеристик и свойств проводимости сигнала.
Мотор
Нижние двигательные нейроны имеют два типа волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр | Миелин | Скорость проводимости | Связанные мышечные волокна |
|---|---|---|---|---|---|
| α | Aα | 13-20 мкм | да | 80–120 м / с | Экстрафузионные мышечные волокна |
| γ | Aγ | 5-8 мкм | да | 4–24 м / с[1][2] | Внутрифузионные мышечные волокна |
Сенсорный
Различные сенсорные рецепторы иннервируются разными типами нервных волокон. Проприоцепторы иннервируются сенсорными волокнами типа Ia, Ib и II, механорецепторы — сенсорными волокнами типа II и III, а ноцицепторы и терморецепторы — сенсорными волокнами типа III и IV.
| Тип | Эрлангер-Гассер Классификация |
Диаметр | Миелин | Скорость проводимости | Связанные сенсорные рецепторы |
|---|---|---|---|---|---|
| Я | Aα | 13-20 мкм | да | 80–120 м / с | Первичные рецепторы мышечного веретена |
| Ib | Aα | 13-20 мкм | да | 80–120 м / с | Орган сухожилия Гольджи |
| II | Aβ | 6-12 мкм | да | 33–75 м / с | Вторичные рецепторы мышечного веретена Все кожные механорецепторы |
| III | Aδ | 1-5 мкм | Тонкий | 3–30 м / с | Свободные нервные окончания от прикосновения и давления Ноцицепторы неоспиноталамического тракта Холодные терморецепторы |
| IV | C | 0,2-1,5 мкм | Нет | 0,5-2,0 м / с | Ноцицепторы палеоспиноталамического тракта Рецепторы тепла |
Автономный
Вегетативная нервная система имеет два типа периферических волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр | Миелин[3] | Скорость проводимости |
|---|---|---|---|---|
| преганглионарные волокна | B | 1-5 мкм | да | 3–15 м / с |
| постганглионарные волокна | C | 0,2-1,5 мкм | Нет | 0,5-2,0 м / с |