Рассмотрим более подробно строение
отростков нейрона и различия между
ними. Как уже было сказано, определяющее
отличие отростков — функциональное,
т.е. направление проведения нервного
импульса: по аксону он проводится от
тела клетки, по дендриту — к телу.
Существует и ряд анатомических различий,
однако они не абсолютны и возможен ряд
исключений из них. Тем не менее, для
типичных аксонов и дендритов характерны
следующие признаки:
-
Аксон один, а дендритов несколько (хотя
существуют нейроны и с одним дендритом). -
Дендрит короче аксона. Длина дендрита
обычно не более700 мкм, а аксон может
достигать длины 1 м. -
Дендрит плавно отходит от
тела нейрона и постепенно истончается.
Аксон, отходя от тела клетки, практически
не меняет диаметр на всем своем
протяжении. Диаметр различных аксонов
колеблется от 0,3 до 16 мкм. От их толщины
зависит скорость проведения нервного
импульса — чем аксон толще, тем скорость
больше. Участок, примыкающий к телу
нейрона (аксонный холмик), имеет большую
толщину, чем остальная часть аксона. -
Дендриты ветвятся на всем
своем протяжении под острым углом,
дихотомически (вильчато), ветвление
начинается от тела клетки. Аксон обычно
ветвится только на конце, образуя
контакты (синапсы) с другими клетками.
Конечные разветвления аксона называют
терминалами. В некоторых местах от
аксонов могут отходить под прямым углом
тонкие ответвления — коллатерали. -
Дендриты (по крайней мере,
в ЦНС) не имеют миелиновой оболочки,
аксоны часто окружены миелиновой
оболочкой (о миелиновой оболочке см.
ниже).
Кроме того, иногда на веточках
дендрита есть выросты
— шипики, являющиеся
характерной структурной особенностью
дендритов, особенно в коре больших
полушарий (рис. 6). Шипик состоит из двух
частей — тела и головки, размеры и форма
которых варьируют. Шипики значительно
увеличивают постсинаптическую поверхность
дендрита. Они являются лабильными
образованиями и при различных воздействиях
(или разных функциональных состояниях)
могут менять свою конфигурацию,
дегенерировать и вновь появляться. В
результате увеличивается либо уменьшается
число синапсов, меняется эффективность
передачи в них нервного сигнала и т.д.

Рис. 6. Шипик на
дендрите нейрона и контактирующие с
ним пресинаптические окончания. Стрелками
показано направление проведения
информации
Теперь, когда мы рассмотрели
строение дендритов и аксонов, следует
несколько детальнее изучить строение
синапса. Синапс, состоящий из одного
пре- и одного постсинаптического
окончаний, называют простым. Однако
большинство синапсов в ЦНС являются
сложными. В таких синапсах один аксон
может контактировать сразу с несколькими
дендритами благодаря нескольким
мембранным выростам на его окончании.
И наоборот, один дендрит за счет своих
шипиков может контактировать с несколькими
аксонами. Еще более сложную структуру
имеют синаптические гломерулы
(клубочки)— компактные
скопления окончаний нервных отростков
разных клеток, формирующие большое
количество взаимных синапсов. Обычно
гломерулы окружены оболочкой из глиальных
клеток. Особенно характерно присутствие
гломерул в тех зонах мозга, где происходит
наиболее сложная обработка сигналов —
в коре больших полушарий и мозжечка, в
таламусе.
Итак, нейрон состоит из тела
(сомы) и отростков. Как правило, один из
отростков существенно длиннее остальных.
Такой длинный отросток называют нервным
волокном. В ЦНС это
всегда аксон; в периферической нервной
системе это может быть как аксон, так и
дендрит. По волокнам проводятся нервные
импульсы, имеющие электрическую природу,
в связи с чем, каждое волокно нуждается
в изолирующей оболочке.
По типу такой оболочки все
волокна делятся на миелиновые
(мякотные) и безмиелиновые
(безмякотные).
Безмиелиновые нервные волокна покрыты
только оболочкой, образованной телом
шванновской (нейроглиальной) клетки.
Эти волокна имеют малый диаметр и
полностью либо частично погружены во
впячивание шванновской клетки. Одна
шванновская клетка может образовывать
оболочку вокруг нескольких аксонов
разного диаметра. Такие волокна называются
волокнами кабельного типа (рис. 7). Так
как длина аксона существенно больше
размеров шванновских клеток, оболочку
аксона образуют цепочки нейроглиальных
клеток. Скорость проведения нервного
импульса по таким волокнам — 0,5-2 м/с.
Многие нервные волокна
имеют миелиновую оболочку. Она также
образуется нейроглиальными клетками.
При формировании такой оболочки
олигодендроцит (в ЦНС) или шванновская
клетка (в периферической нервной системе)
обхватывает участок нервного волокна
(рис. 8). После этого образуется вырост
в виде язычка, который закручивается
вокруг волокна, образуя мембранные слои
(цитоплазма при этом из «язычка»
выдавливается). Таким образом, миелиновая
оболочка представляет собой двойные
слои клеточной мембраны и по своему
химическому составу является липопротеидом,
т.е. соединением липидов (жироподобных
веществ) и белков. Миелиновая оболочка
осуществляет электрическую
изоляцию нервного волокна наиболее
эффективно. Нервный импульс проводится
по такому волокну быстрее,
чем по лишенному миелина (скорость
проведения может достигать
120 м/с). Миелиновая оболочка начинается
немного
отступя от тела нейрона и заканчивается
примерно в 2
мкм от синапса. Она состоит из цилиндров
длиной 1,5-2
мм,
каждый из которых образован своей
глиальной клеткой.
Цилиндры разделяют перехваты Ранвье —
не покрытые
миелином участки волокна (их длина 0,5 —
2,5 мкм), играющие большую роль в быстром
проведении нервного импульса. В перехватах
от аксона могут отходить коллатерали.
Поверх миелиновой оболочки у мякотных
волокон есть еще
наружная оболочка — неврилемма,
образованная цитоплазмой и ядром
нейроглиальных клеток.

Рис. 7. Строение
нервных волокон:
А
—
миелиновое;
Б
—
безмиелиновая;
I
—
волокно;
2 —
миелиновый
слой; 3—
ядро
шванновской клетки; 4 —
микротрубочки;
5—Нейрофиламенты;
6 —
митохондрии;
7—соединительнотканная
оболочка
Рис.
8.
Строение миелиновой оболочки (А).
Образование
миелиновой оболочки шванновской клеткой
(Б): ‘
1
—
аксон;
2 —
слои
миелиновой оболочки; 3
— перехваты
Ранвье;
4
—
ядро
шванновской клетки. Стрелкой показано
направление
продвижения
выроста цитоплазматической мембраны
Миелин
имеет белый цвет. Именно это его свойство
позволило
разделить вещество нервной системы на
серое и белое. Тела
нейронов и их короткие отростки образуют
более темное серое
вещество, а
волокна — белое
вещество.
-
Классификация
нейронов
Нейроны очень
разнообразны по форме, величине,
количеству и способу отхождения от тела
отростков, химическому строению (имеется
в виду, в первую очередь, синтез тех или
иных нейромедиаторов) и т.д. (рис. 9). Тела
самых крупных нейронов достигают в
диаметре 100 — 120 мкм (гигантские пирамиды
Беца в коре больших полушарий), самых
мелких — 4-5 мкм (зернистые клетки коры
мозжечка). Приведем основные способы
классификации нервных клеток.



Рис.
9.
Различные типы нейронов:
А
—
псевдоуниполярный
нейрон спинномозгового ганглия;
Б
—
биполярный
нейрон сетчатки; В —
мотонейрон
спинного мозга;
Г
—
пирамидная
клетка коры больших полушарий (видно,
что дендриты покрыты
шипиками); Д —
клетка
Пуркинье мозжечка; I
—
тело
клетки;
2
—
дендрит;
3 —
аксон;
4 —
коллатерали
аксона
-
Функционально
нейроны подразделяются на чувствительные
(сенсорные), вставочные (переключательные,
интернейроны)
и исполнительные (двигательные или
мотонейроны и
др.). Сенсорные
нейроны —
это нервные клетки, воспринимающие
раздражения из внешней или внутренней
среды организма.
Интернейроны
(вставочные
нейроны) обеспечивают связь
между чувствительными и исполнительными
нейронами рефлекторных дугах. Общее
направление эволюции нервной системы
связано с увеличением числа интернейронов.
Из более
чем ста миллиардов нейронов человека
более 70% составляют
вставочные нейроны.
Исполнительные
нейроны,
управляющие сокращениями поперечно —
полосатых мышечных волокон, называют
двигательными
(мотонейронами). Они
образуют нервно-мышечные синапсы.
Исполнительные нейроны, называемые
вегетативными, управляют
работой внутренних органов, включая
гладкомышечные волокна, железистые
клетки и др.
2. По количеству отростков
нейроны делятся на униполярные,
псевдоуниполярные, биполярные и
мультиполярные.Большинство
нейронов нервной системы (и почти все
нейроны в ЦНС) — это мультиполярные
нейроны (см.
рис. 9, В — Д), они имеют
один аксон и несколько дендритов.
Биполярные
нейроны (см.
рис. 9, Б) имеют один аксон и один дендрит
и характерны для
периферических отделов анализаторных
систем. Униполярных
нейронов, имеющих только один отросток,
у человека практически
нет. Из тела псевдоуниполярного
нейрона (см.
рис.9,
А) выходит один отросток, который
практически сразу делится
на две ветви. Одна из них выполняет
функцию дендрита, а
другая — аксона. Такие нейроны находятся
в чувствительных спинномозговых
и черепных ганглиях. Их дендрит
морфологически
(по строению) похож на аксон: он гораздо
длиннее аксона и часто имеет миелиновую
оболочку.
3. По форме тела и характеру
ветвления отростков выделяют звездчатые,
пирамидные, веретеновидные, корзинчатые,
зернистые
и др. нейроны.
-
По длине аксона нейроны делят
на клетки типа Гольджи I и типа Гольджи
II (эта классификация разработана
итальянским ученым К.
Гольджи). Клетки Гольджи I
имеют длинный
аксон, выходящий за пределы области, в
которой находится
тело нейрона. Это, например, пирамидные
клетки коры
больших полушарий. У клеток Гольджи II
короткий и, как
правило, очень разветвленный аксон, не
выходящий за пределы
области, в которой находится тело
нейрона. Примером
таких нейронов могут быть корзинчатые
клетки коры мозжечка. -
Каждый нейрон синтезирует
только один основной нейромедиатор.
Для того чтобы определить нервную
клетку с этой
точки зрения к названию медиатора
добавляют окончание
«-ергический». Например, ацетилхолинергический
нейрон образует ацетилхолин,
глицинергический — глицин и т.д.
Соседние файлы в предмете Анатомия и физиология
- #
01.06.201515.58 Mб544ОТ НЕЙРОНА К МОЗГУ.doc
- #
- #
- #
- #
- #
Что такое дендрит — функции и морфология
Дендриты (dendrite) — многочисленные тонкие трубчатые или округлые выпячивания клеточного тела (перикариона) нервной клетки. Сам термин говорит о чрезвычайной разветвленности этих участков нейронов (от греч. δένδρον (dendron) — дерево).
В поверхностной структуре нейроцитов могут насчитываться от нуля до множества дендритов. Аксон чаще всего единственный. Поверхность дендритов не имеет миелиновой оболочки в отличие от аксонных отростков.

Цитоплазма содержит те же клеточные компоненты, что и само тело нервной клетки:
- эндоплазматический гранулярный ретикулум;
- скопления рибосом — полисомы (белоксинтезирующие органеллы);
- митохондрии (энергетические “станции” клетки, которые, используя глюкозу и кислород, синтезируют необходимые высокоэнергетические молекулы);
- аппарат Гольджи (отвечает за доставку внутренних секретов к внешнему слою клетки);
- нейротубулы (микротрубочки) и нейрофиламенты — главные компоненты цитоплазмы, тонкие опорные структуры, которые обеспечивают сохранение определенной формы.
Строение дендритных окончаний напрямую связано с их физиологическими функциями — получением информации от аксонов, дендритов, перикариона соседних нервных клеток посредством многочисленных межнейронных контактов на основе избирательной чувствительности к определенным сигналам.
Строение нейрона: аксоны и дендриты
Основные статьи: Дендрит,Аксон
Схема строения нейрона
Аксон— обычно длинный отросток нейрона, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу.Дендриты— как правило, короткие и сильно разветвлённые отростки нейрона, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновойоболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Основная статья: Синапс
Си́напс(греч.σύναψις, отσυνάπτειν— обнимать, обхватывать, пожимать руку) — место контакта между двумянейронамиили между нейроном и получающей сигналэффекторнойклеткой. Служит для передачинервного импульсамежду двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
- Общая информация
- Аксоны нейрона
- Дендриты нейрона
- Тело
Важнейший элемент в нервной системе – нейронная клетка, или простой нейрон. Это специфическая единица нервной ткани, задействованная в передаче и первичной обработке информации, а так же, являющаяся главным структурным образованием в ЦНС. Как правило, клетки имеют универсальные принципы строения и включают в себя помимо тела, еще аксоны нейронов и дендриты.
Структура и типы
Внешняя поверхность дендритов покрыта тонкими выпячиваниями в виде мельчайших шипиков размером 2-3 мкм. Количество таких формирований на поверхности может варьировать от нуля до десятка тысяч. Формы самих микрошипиков многообразны, но самой часто встречаемой формой считается грибовидный шипик.
Количество шипиков на поверхности и их размеры могут быстро меняться. От этого зависит реакция нейрона на сигналы от других клеток.
На образование выпячиваний-шипиков, их форму и развитие влияют внутренние и внешние обстоятельства: возраст организма, активность синаптических связей, информационная загруженность нейронных цепей, образ жизни организма и многое другое.
Целостность и стабильность структуры шипиков могут подвергаться влиянию негативных факторов:
- патофизиологические факторы (например, нейродегенеративные процессы в нервной ткани, опосредованные тяжелой наследственностью);
- токсикологические агенты (при употреблении наркотиков, алкоголя, ядов различной природы).
Под воздействием этих негативных факторов во внутреннем строении микрошипиков происходят серьезные деструктивные превращения: разрушение цистерн шипикового аппарата, накопление мультивезикулярных тел (пропорционально степени разрушительных влияний).
После серии испытаний, проведенной с подопытными мышами, было доказано, что не столько сами дендриты, сколько дендритные шипики являются элементарными единицами хранения памяти и формирования синаптической пластичности.

Ветвление
Дендритные структуры образуются вследствие древовидного разветвления отростков нейронов. Этот процесс называется арборизацией. Количество точек (или узлов) ветвления обуславливает степень разветвленности и сложность окончаний дендрита. В цитоплазме узлов ветвления обычно сконцентрированы митохондрии, так как ветвление – энергозатратный физиологический процесс.
Структура дендритного дерева обуславливает физическую восприимчивую площадь, то есть количество входных импульсов, которые суммарно сможет принять и провести нейроцит.
Одно из главных предназначений дендритов состоит в наращивании контактной поверхности для синапсов (увеличении рецепторного поля).
Это позволяет клетке принимать и перенаправлять больший объем информации, которая поступает к телу нейрона. Степень разветвленности определяет то, как нейрон в итоге суммирует электрические сигналы, полученные от других клеток: чем больше и сложнее ветвление, тем более плотно нейроны прилегают друг к другу.
За счет разветвленного строения поверхность рецепторной мембраны нервной клетки увеличивается в 1000 и более раз.

Диаметр и длина
Дендритные окончания имеют разные размеры, но всегда характеризуются постепенным уменьшением диаметра претерминальных веточек. Длина обычно от нескольких мкм до 1 мм. Но, например, у некоторых чувствительных нейронов спинномозговых ганглиев дендриты очень длинные – до метра и более.
Строение дендрита
На основании изучения микроскопических препаратов нервных клеток установили, что большинство отростков имеют цилиндрическую форму. Их диаметр в среднем составляет 0,9 мкм. Длина дендритов варьирует в широких пределах. Например, звездчатые нейроны серого вещества коры головного мозга имеют короткие (не более 200 мкм) ветви дендритного дерева, тогда как отростки двигательного нейрона, входящего в передние рога спинного мозга, составляют порядка 2 мм.
Специальные образования — шипики, формирующиеся на ветвях нейроцитов, приводят к появлению большого числа синапсов – щелевидных мест контакта с аксоном, дендритом или сомой другого нейрона. Синапсы могут располагаться на теле дендрита и называются стволовыми или же непосредственно на его шипиках. Как мы уже знаем, дендриты – это разветвленные отростки нейроцитов, способные принимать возбуждение. Передача же биопотенциалов происходит в них с помощью молекул химических соединений — медиаторов, например, ГАМК или ацетилхолина. В мембране, покрывающей дендрит, обнаружены ионные каналы, избирательно пропускающие катионы кальция, натрия и калия, участвующие в прохождении нервных импульсов через нейрон.

Проведение нервного импульса
Рецепторная мембрана поверхности дендритов (как и тела нервной клетки) покрыта многочисленными синаптическими бляшками, которые передают возбуждение на восприимчивый участок поверхностной мембраны нейрона, где генерируется биоэлектрический потенциал.
Информация, закодированная в виде электрических импульсов, передается на электровозбудимую проводящую мембрану аксона. Таким образом формируются нейронные сети организма.

На что влияет разветвленность нервных отростков
Тело нейрона является универсальным передающим и одновременно принимающим биологическим объектом. Объем (прежде всего поступающей информации) прямо пропорционален количеству входящих нервных импульсов. Они определяются по степени ветвления дендритного дерева. Поэтому дендриты – это структуры нейроцита, играющие интегративную функцию.

Более того, отростки расширяют площадь контакта нервных клеток между собой. Дополнительное же образование синапсов в разы повышает эффективность работы всех отделов, как головного и спинного мозга, так и нервной системы в целом.
Дендритная кристаллизация
В случае увеличения температуры в сторону жидкой фазы, то есть при положительном градиенте температуры , образуются относительно плоские грани кристаллов. Пока кристаллы окружены жидкой фазой, они растут свободно и имеют почти правильную геометрическую форму, которая определяется анизотропией скорости роста. В результате взаимного столкновения граней соседних кристаллов на завершающей стадии кристаллизации их правильная форма искажается. Так образуются кристаллиты произвольной формы.
В металлах и сплавах вследствие интенсивного выделения скрытой теплоты кристаллизации на границе кристалла и сильно переохлажденного расплава возникает отрицательный градиент температуры, который меняет механизм роста кристаллов. Чем дальше от границы, тем больше является переохлаждение расплава. Поэтому любая выпуклость на поверхности зародыша попадает в зону большего переохлаждения и растет вглубь расплава быстрее, чем плоские участки поверхности, образуя вытянутую первичную ось кристалла.
Направление роста осей кристалла совпадает с направлением оси пирамиды, которую образуют плоскости густого упаковки атомов. Поэтому в кристаллах с кубической или тетрагональной решеткой оси кристаллитов перпендикулярны между собой, а с гексагональной — размещены под углом 60 °. Наконец, когда температуры осей кристалла и расплава между ними выравниваются, начинается медленнее кристаллизация в межосевому пространстве. Так из зародышей вырастают древовидные кристаллиты — дендриты (с греч. Δένδρον — дерево). Механизм такого роста называют дендритным.
Дендриты растут до момента столкновения с соседями на завершающей стадии кристаллизации, приобретая при этом нерегулярной формы и определенных размеров. Размер дендритных кристаллитов является важным фактором, определяющим свойства литых сплавов. Мелкодисперсная структура обеспечивает лучшие механические и технологические свойства по сравнению с грубодисперсной. В практике литейного производства для большинства отливок стремятся обеспечить мелкозернистую структуру, реализующих преимущественно модифицирования или из-за изменения температуры расплава или скорости охлаждения при кристаллизации.
( 1 оценка, среднее 5 из 5 )
https://ru.wikipedia.org/wiki/%D0%9D%D0%B5%D0%B9%D1%80%D0%BE%D0%BD
Пирамидальные ячейки нейронов в коре головного мозга мыши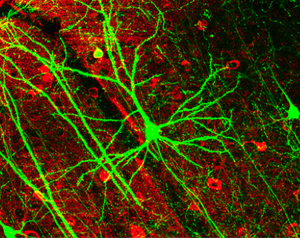
Нейрон (от греч. néuron — нерв) (неврон) — это структурно-функциональная единица нервной системы, принимающая сигналы, поступающие от экстерорецепторов и др.. Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов.[1]
Обзор
Сложность и многообразие нервной системы зависит от взаимодействия между нейронами, которые, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
Эфир и нелокальность
Поскольку наблюдается дальнодействие нейронов [1]-[2], природа передачи сигналов в нейросистемах остается открытым вопросом. Этот вопрос часть общей проблемы отношения эфир-интеллект, нелокальность.
- [1] Cell Intelligence. Guenter Albrecht-Buehler
- [2] Ultimate Computing. Stuart R. Hameroff
Строение
Нейрон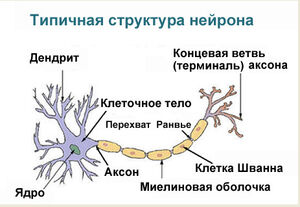
Тело клетки
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), снаружи ограничена мембраной из двойного слоя липидов(билипидный слой). Липиды состоят из гидрофильных головок и гидрофобных хвостов, расположены гидрофобными хвостами друг к другу, образуя гидрофобный слой, который пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: на поверхности (в форме глобул), на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в них находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро (с большим количеством ядерных пор) и другие органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), и отростков. Выделяют два вида отростков: дендриты и аксон.
Нейрон имеет развитый цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Схема строения нейрона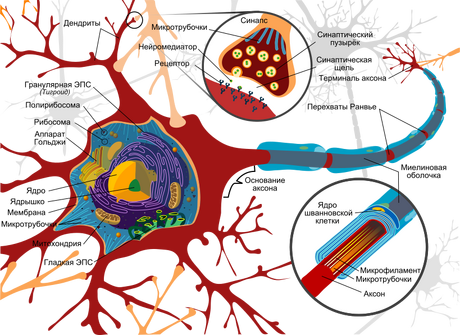
Аксон — обычно длинный отросток, приспособленный для проведения возбуждения от тела нейрона.
Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Си́напс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Визуализации нейроструктур
На каждой обложке журнала The Journal of Neuroscience печатаются (доступные on-line) высококачественные визуализации нейросистем.
Классификация
Структурная классификация
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
Мультиполярные нейроны — Нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — неультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — эта группа нейронов осуществляет связь между эфферентными и афферентными, их делят на комиссуральные и проекционные (головной мозг).
Морфологическая классификация
Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона,
- количество и характер ветвления отростков,
- длину нейрона и наличие специализированные оболочки.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов. Длина нейрона у человека составляет от 150 мкм до 120 см.
По количеству отростков выделяют следующие морфологические типы нейронов (Puc.[1]):
— униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге ;
— псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях ;
— биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях ;
— мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС .
Развитие и рост нейрона
Конус роста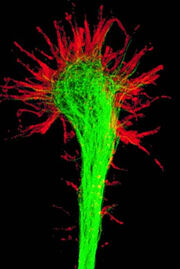
Нейрон развивается из небольшой клетки — предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным. [2] (рус.)) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
См. также
- Нервные волокна
- Аксон
- Дендрит
- Синапс
- Компьютер
- Искусственный нейрон
- Нейронная сеть
Примечания
- ↑ http://bse.sci-lib.com/article080889.html
Гистология: Нервная ткань |
|
|---|---|
| Нейроны (Серое вещество) |
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты) Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит, Базальный дендрит) типы: Биполярные нейроны · Псевдополярные нейроны · Мультиполярные нейроны · Пирамидальный нейрон · Клетка Пуркинье · Гранулярная клетка |
| Афферентный нерв/ Сенсорный нерв/ Сенсорный нейрон |
GSA · GVA · SSA · SVA · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено, II or Aβ, Aδ-волокна, C-волокна) |
| Эфферентный нерв/ Моторный нерв/ Моторный нейрон |
GSE · GVE · SVE · Верхний моторный нейрон · Нижний моторный нейрон (α мотонейроны, γ мотонейроны) |
| Синапс |
Нейропиль · Синаптический пузырек · Нервно-мышечный синапс · Электрический синапс · Интернейрон (Клетки Реншоу) |
| Сенсорный рецептор |
Чувствительное тельце Мейснера · Нервное окончание Меркеля · Тельца Пачини · Окончание Руффини · Нервномышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица |
| Нейроглия |
Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия |
| Миелин (Белое вещество) |
CNS: Олигодендроцит PNS: Клетки Шванна · Невролемма · Перехват Ранвье/Межузловой сегмент · Насечка миелина |
| Соединительная ткань |
Эпиневрий · Периневрий · Эндоневрий · Нервные пучки · Оболочки мозга |
Дендрит – это короткий, дихотомически (раздваивающийся) ветвящийся отросток, отходящий от тела нейрона. Согласно определению, отросток образует синапсы (электрические, химические) с другими элементами нервной ткани, через которые получает сигналы от периферических рецепторов и клеток организма в гистологической структуре органов и систем. Слово «дендрит» также имеет другое значение, если используется не в медицинской сфере, а в области материаловедения, в данном случае подразумевается кристаллическая структура материала независимо от его состава. Термин применим к камням, металлам, сплавам, искусственным соединениям, обладающим кристаллической, древовидной структурой.

Синаптическая щель
Сигнал от других клеток поступает к телу нейрона по одному из его дендритов. Дендрит в нервной системе человека получает обычно химический сигнал (нейромедиатор) от аксона. Место соединения дендрита и аксона называется синапсом.
Синапсы позволяют передавать точные сообщения от нейрона к нейрону. Благодаря синапсам существует нейропластичность и возможность тонкой настройки функций и поведения организма.

На дендрите находятся рецепторы, которые принимают нейромедиатор. Рецепторы — это специализированные белки, которые захватывают молекулу нейромедиатора и в зависимости от своего типа запускают дальнейшие реакции в клетке.
Общие сведения
Нейрон – основная структурно-функциональная единица нервной ткани. Нервная клетка – образование с многочисленными отростками размером 4-130 мкм. От нейрона отходят несколько (реже один) дендритов и единственный аксон. Дендрит в биологии – это такой отросток, который передает возбуждение от периферических рецепторов к телу нейрона, что обуславливает его ведущую роль в восприятии внешних стимулов. Особенности дендритных ответвлений, которые наблюдаются в ходе микроскопических исследований:
- Система микротрубочек.
- Наличие шипиков, отходящих от общей оси.
- Присутствие узлов ветвления.
- Наличие эндоплазматического ретикулума (внутриклеточный органоид, представленный системой канальцев, полостей, пузырьков, окруженных мембраной).
Отростки, находящиеся рядом с сомой (телом), утолщенные, образуют большое число синаптических контактов. Мембрана отростка наподобие мембраны самого нейрона состоит из большого количества белковых молекул, которые играют роль химических рецепторов. Рецепторные образования наделены специфической чувствительностью к определенным химическим соединениям.
Обозначенные химические вещества – нейромедиаторы торможения и возбуждения, активно участвуют в процессе, когда импульсы распространяются по нервной ткани и поступают к соме. Строение дендрита предполагает наличие шипиков, которые образуют синаптические контакты с терминалями – концевыми участками сотен тысяч нервных клеток. Огромное количество шипиков располагается на отростках нервных клеток, образующих корковый слой больших полушарий.
Дендритный шипик сформирован из тела и головки. Размеры и форма структурных компонентов шипика существенно варьируются. Благодаря шипикам значительно увеличивается площадь постсинаптической мембраны. Шипики в дендритной структуре – лабильные образования, которые под воздействием внешних стимулов изменяют конфигурацию, дегенерируют (разрушаются), регенерируют (появляются вновь).
Количество синапсов определяет качество передачи импульсов и скорость, с которой они распространяются. Несколько дендритных ответвлений образуют единую ветку. Совокупность всех дендритов является дендритным деревом – поверхностью, воспринимающей сторонние раздражители. Исследования показывают, дендритные деревья составляют 90% мозгового вещества.

Дендритные шипики
На дендритах образуются маленькие наросты — шипики. Последние могут принимать множество форм, но наиболее устойчивая — это форма грибка.
Количество дендритных шипиков колеблется от 20 до 50 на 10 мкм длины дендрита. Шипики очень изменчивы по форме и объему.
В мозге 86 миллиардов нейронов. Аксоны, дендриты и тела нейронов образуют огромные нейронные сети.
Дендриты отвечают за обучение и память, а также контролируют равновесие в системе. Когда происходит локальное усиление связей между определенными нейронами, именно в дендритах возрастает производство белка, регулирующего снижение активности других синапсов.

Отличия от аксонов
Аксон служит для передачи нервных импульсов от тела нервной клетки, которое по-другому называется сома, к исполнительным органам. Окончание аксона является элементом синапса, через который осуществляется синаптическая передача сигналов между отдельными клетками нервной ткани.
Дендритные клетки обладают разветвленной структурой. Дендритные отростки ветвятся на всем протяжении в отличие от аксона, который разветвляется только в конечном сегменте, образуя терминали. В отличие от аксона, длина которого может превышать 1 метр, дендрит – короткий отросток (около 700 мкм). Другие различия между дендритом и аксоном:
- Разнонаправленное проведение импульсов (дендриты – к телу нейрона, аксон – от тела нейрона).
- Разная толщина (дендрит истончается по мере удаления от тела, аксон сохраняет одинаковые значения диаметра сечения по всей протяженности). Диаметр аксонов разных клеток составляет около 0,3-16 мкм. Чем толще аксон, тем выше скорость распространения по нему импульсов.
- Наличие миелиновой оболочки (у дендритов ЦНС миелиновая оболочка отсутствует, у аксонов – присутствует).
Дендритный транспорт предусматривает движение по стволу отростка белковых веществ и ферментов от сомы к конечным сегментам. В отличие от дендритного транспорта, аксональный транспорт предполагает непрерывный ток аксоплазмы в обоих направлениях. Механизм транспорта поддерживается благодаря микротрубочкам и белкам (кинезин – движение внутри микротрубочек, динеин – движение по поверхности микротрубочек).
Движение веществ по стволу осуществляется посредством затрат АТФ. Размеры дендритных отростков коррелируют с активностью нейронов. Стимулы, поступающие из внешней среды, преобразуются в биоэлектрические сигналы. Нервный импульс представляет собой волну возбуждения, распространяющуюся по отростку. Процесс образования энергии, необходимой для поддержания дендритного транспорта, происходит в митохондриях.

Обучение и шипики
Дендритные шипики отвечают за возможность обучения и формирования памяти. Благодаря шипикам и их пластичности, нейрон легко может подключаться к тем или другим соседям и быстро от них отсоединяться, контролируя возможность получения сигнала.
Логично было бы предположить, что если синаптические связи ответственны за воспоминания, то их пластичность — проблема для сохранения памяти о прошлом. В 2009 году в Nature вышла публикация, в которой авторы исследовали, как опыт обучения влияет на синаптические связи мышей.

В работе показано, что большое количество новых шипиков, образующихся от нового опыта, исчезало со временем, если опыт не повторялся периодически. Но те, что сохранялись, скорее всего, и отвечали за приобретенные навыки.
При этом если тренировка повторялась в течение длительного времени, происходило удаление шипиков, по-видимому, удаленные отвечали за неверные действия. Обучение и ежедневный сенсорный опыт оставляют постоянные пометы в виде немногочисленной группы шипиков, сформированных на разных этапах обучения.
Что такое дендриты, если не огромная библиотека воспоминаний? Но основная проблема дендритных шипиков в том, что они очень чувствительны к любым механическим и химическим воздействиям. Поэтому травмы мозга, даже если и локализованы в одном месте, обычно оказывают влияние на всю нейронную сеть.
Сон и обучение
В исследовании (Z.G. Yang) 2014 года было показано, как после обучения и сна, спустя 24 часа, появляются новые дендритные шипики у мышей, а некоторые из существующих исчезают. Авторы отмечают, что скорость образования новых шипиков у мышей, прошедших обучение новому поведению, была значительно выше в течение 6 часов после обучения по сравнению с нетренированными мышами.

Кроме того, авторы показали, что шипики при лишении мышей сна формируются намного медленнее. И ситуацию не может исправить ни новая тренировка навыка, ни поздний сон.

Дендрит как самостоятельная единица
Что такое дендриты, выясняют до сих пор. Дело в том, что сложно изучать поведение и функции дендритов на живых объектах.
Если размер нейрона около десяти микрон, то длина дендрита может доходить до тысячи. Обычно под дендритами понимают не очень активных участников процесса.
В 2020 году в журнале Science было опубликовано исследование, которое позволяет пересмотреть классический взгляд на дендриты. Оказалось, дендриты генерируют сигналы в несколько раз чаще, чем это делает тело нейрона, что наводит на предположение о кодировании информации и на уровне дендритов.

Ранее уже было обнаружено, что если во время переживания опыта тела нейронов активировались, а дендриты молчали, то долговременная память не формировалась относительно этого опыта. Было высказано предположение, что активность нейронов связана в большей степени с реальным временем, с актуальными переживаниями, а дендритов — с тем, что от этого останется в памяти.
Что такое дендриты, учитывая новые данные? Это удивительные конструкции, которые составляя 90% нервной ткани и, возможно, берут на себя большую часть работы по сохранению и преобразованию опыта.
Особенности строения и функции аксонов и дендритов.
Аксон (греч. ἀξον — ось) — нейрит, осевой цилиндр, отросток нервной клетки, по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам. В месте соединения аксона с телом нейрона находится аксонный холмик. Регистрация электрических потенциалов выявила, что нервный импульс генерируется в самом аксоне, а именно в начальном сегменте. Питание и рост аксона зависят от тела нейрона: при перерезке аксона его периферическая часть отмирает, а центральная сохраняет жизнеспособность. При диаметре в несколько микронов длина аксона может достигать у крупных животных 1 метра и более (например, аксоны, идущие от нейронов спинного мозга в конечности). У многих животных встречаются гигантские аксоны толщиной в сотни мкм (у кальмаров — до 2—3 мм). Обычно такие аксоны отвечают за проведение сигналов к мышцам.обеспечивающим «реакцию бегства» (втягивание в норку, быстрое плавание и др.). При прочих равных условиях с увеличением диаметра аксона увеличивается скорость проведения по нему нервных импульсов.
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна. Миелиновая оболочка аксонов имеется только у позвоночных. Её образуют «накручивающиеся» на аксон специальные шванновские клетки, между которыми остаются свободные от миелиновой оболочки участки — перехваты Ранвье. Только на перехватах присутствуют потенциал-зависимые натриевые каналы и заново возникает потенциал действия. При этом нервный импульс распространяется по миелинизированным волокнам ступенчато, что в несколько раз повышает скорость его распространения. Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью.
Дендрит.
Основные характерные черты дендрита, которые выделяют его на электронно-микроскопических срезах:
§ отсутствие миелиновой оболочки
§ наличие правильной системы микротрубочек
§ наличие на них активных зон синапсов с ясно выраженной электронной плотностью цитоплазмы дендрита
§ отхождение от общего ствола дендрита шипиков,
§ специально организованные зоны узлов ветвлений,
§ вкрапление рибосом
§ наличие в проксимальных участках гранулированного и не гранулированного ЭПР.
Наиболее примечательной особенностью цитоплазмы дендритов является наличие многочисленных микротрубочек. Они хорошо выявляются как на поперечных срезах, так и на продольных. Микротрубочки следуют в дендрите параллельно друг другу, не соединяясь и не пересекаясь между собой. Шипик является производным дендрита, в нем отсутствуют нейрофиламенты и дендритические трубочки, его цитоплазма содержит грубо или тонко гранулированный матрикс. Еще одной характерной чертой шипика в коре мозга является обязательное присутствие на них синаптических контактов с аксонными окончаниями. Можно отметить своеобразную триаду в цитоплазме шипика: субсинаптическая специализация активных зон — шипиковый аппарат — митохондрии. Можно говорить о том, что цитоплазма дендритического шипика и шипиковый аппарат могут иметь непосредственное отношение к синаптической функции.
Дендритические шипики и концы дендритов также очень чувствительны к действию экстремальных факторов. При любом виде отравления (например, алкогольном, гипоксическом, тяжелыми металлами — свинцом, ртутью и т.д.) меняется количество выявленных шипиков на дендритах клеток коры больших полушарий. По всей вероятности, шипики при этом не исчезают, но у них нарушаются цитоплазматические компоненты, и они хуже импрегнируются солями тяжелых металлов. Так как шипики — один из структурных компонентов обеспечения межнейрональных контактов, то неполадки в них приводят к серьезным нарушениям функции мозга.
Для всех дендритов ЦНС характерно увеличение поверхности за счет многократного дихотомического деления. При этом образуются в зонах деления особые расширительные площадки или узлы ветвления.
Функции дендритов.
· трофическая функция
· передача нервного импульса от одной клетки к другой, также отвечает за частоту и амплитуду сигнала.
 Когда с нами что-то происходит наш мозг фиксирует это, создавая воспоминания. Изменения, которые при этом происходят с мозгом, принято называть энграммами или следами памяти.
Когда с нами что-то происходит наш мозг фиксирует это, создавая воспоминания. Изменения, которые при этом происходят с мозгом, принято называть энграммами или следами памяти.
Вполне естественно, что понимание того, как выглядят следы памяти – основной вопрос изучения мозга. Без этого невозможно построить никакую биологически достоверную модель его работы. Понимание строения памяти непосредственно связано с пониманием того, как мозг кодирует информацию и как он ей оперирует. Все это, пока, — неразгаданная загадка.
Еще большую интригу в загадку памяти вносят исследования по локализации воспоминаний. Еще в первой половине двадцатого века Карл Лэшли поставил очень интересные опыты. Сначала он обучал крыс находить выход в лабиринте, а затем удалял им различные части мозга и снова запускал в тот же лабиринт. Так он пытался найти ту часть мозга, которая отвечает за память о полученном навыке. Но оказалось, что память каждый раз сохранялась, несмотря на временами значительные нарушения моторики. Крысы всегда помнили где искать выход и упорно стремились к нему.

Эти опыты вдохновили Карла Прибрама сформулировать ставшую популярной теорию голографической памяти. В соответствии с ней, аналогично оптической голограмме, каждое конкретное воспоминание не находится в каком-то одном месте коры, а присутствует в каждом ее месте и, соответственно, каждое место коры хранит сразу все воспоминания.
Одно время, очень большие надежды в поисках энграмм связывались с синаптической пластичностью. Способность синапсов менять свою чувствительность давала надежду, что через это можно описать все механизмы памяти. Представление о пластичности синапсов привело к созданию искусственных нейронных сетей. Эти сети показали, как нейрон может обучиться узнавать, что-то общее для набора воспоминаний. Но узнавать общее — это совсем не то же, что хранить отдельные воспоминания.
Если вы не занимайтесь непосредственно нейронауками, то скорее всего у вас сложилось впечатление, что у нейробиологов есть множество теорий относительно памяти, но, видимо, нет уверенности, какая из них верна. И так как, скорее всего, эти теории очень сложны, то о них не особо рассказывают в популярной литературе. Так вот, до сих пор, как ни удивительно это звучит, нет ни одной теории памяти. То есть, есть разные предположения о том, что может быть связано с памятью. Но нет никаких моделей, которые бы хоть как-то объясняли, как выглядят энграммы и как они работают.
При этом накоплены огромные знания о биологии нейронов, о проявлениях памяти, о молекулярных процессах, сопутствующих формированию воспоминаний и тому подобное. Но углубление знаний не упрощает ситуацию, а только усложняет ее. Пока о предмете исследований известно не особо много, удобно фантазировать. Полет фантазии не сильно ограничен рамками знаний. Но по мере того, как становятся известны все новые и новые факты, многие гипотезы отпадают сами собой. Придумать же новые, которые находились бы в согласии с фактами становится все труднее.
Когда в науке наступает такая ситуация – это верный признак того, что где-то в самое начало рассуждений закралась фатальная ошибка. В свое время Аристотель сформулировал законы движения. Он, исходил из того, что видел перед собой. Аристотель сказал, что есть два вида движения: естественное движение и принудительное движение. Естественное движение, по Аристотелю, присуще только небесному веществу и только небесные тела могут двигаться без прикладывания силы. Все остальные «земные» тела для движения требуют приложения силы, иначе любое движение рано или поздно должно прекратиться. Почти две тысячи лет это считалось очевидной истиной, так как все остальные видели перед собой то же самое. Но при этом, почему-то, никому за все эти два тысячелетия не удалось построить ни одной работоспособной теории, которая пошла бы дальше утверждений Аристотеля. И только, когда Галилей и Ньютон указали на досадную ошибку Аристотеля, что, дескать, он забыл про силу трения, оказалось возможным сформулировать известные нам законы механики. Затем, правда, был Эйнштейн, но это уже другая история.
Мне кажется, что таким «принудительным движением», сейчас, в нейронауке выступает «нейрон бабушки». По сути, все основные трудности с построением теории памяти связаны с тем, что оказывается очень трудной задачей увязать конкретный нейрон, если приписывать ему функции детектора какого-либо свойства, и память, которая в силу многих причин, не должна быть жестко привязана к определенному нейрону.
Далее я покажу, как могут выглядеть энграммы для случая, когда нейроны лишаются своих «бабушкиных» наклонностей.
В предыдущих частях был описан клеточный автомат, состоящий из однородных элементов. Когда в любом месте этого автомата создается какой-либо узор активности от этого места расходится фронт волны. В каждом месте этого фронта возникает уникальный специфичный, только для этой волны узор.

Если запомнить какой рисунок создает волна, проходя через какое-либо место, то затем можно в том же месте воспроизвести этот же рисунок и запустить из этого места новую волну. В каждом месте, которое на своем пути пройдет эта новая волна, она будет повторять узор исходной волны.
Если составить словарь, состоящий из конечного числа понятий, то каждому понятию можно сопоставить свою уникальную волну. Тогда, в любом месте автомата по узору проходящей волны можно будет определить, какое понятие распространяет эта волна. И из любого места можно будет запустить волну любого понятия если воспроизвести в этом месте фрагмент узора нужной нам волны.
Плоскому авомату можно придать объем.

Прохождение волны в небольшом цилиндрическом объеме тогда будет выглядеть, как показано на рисунке ниже.
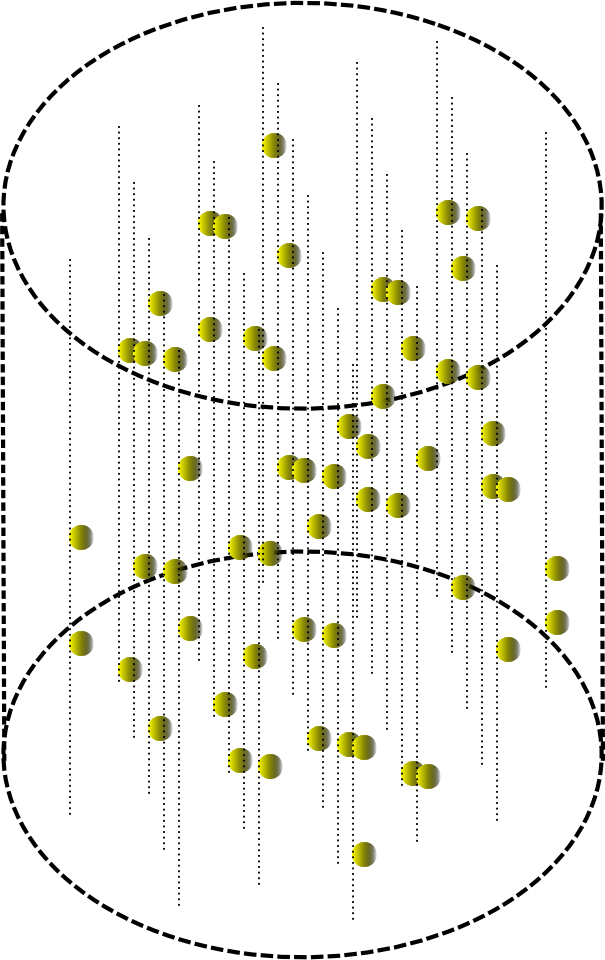
Если запустить в автомате информационную волну, а затем запустить волну идентификатора. То можно запомнить картину условной “интерференции” этих волн. Для этого в каждом месте автомата элементами, по которым прошла информационная волна, надо запомнить окружающий их узор волны идентификатора. Такая процедура позволяет запомнить пару «ключ – значение». Если впоследствии запустить в автомат волну идентификатора воспоминания, то элементы автомата воспроизведут узор информационной волны самого воспоминания.
Запомнить информационную пару «ключ – значение» можно, как избирательно в любой малой области автомата, так и глобально во всем пространстве автомата. При глобальном запоминании информация оказывается многократно продублирована по всей площади автомата.
Когда информационное описание состоит не из одного понятия, а из нескольких, то передать такое описание по автомату можно, последовательно распространив информационные волны этих понятий.
В каждом фиксированном объеме автомата прохождение серии волн вызовет смену узоров, каждый из которых можно записать бинарным вектором. Если в описании неважна последовательность понятий, то для одного места автомата бинарные вектора, создаваемые разными волнами можно побитно логически сложить и получить суммарный вектор описания. Этот суммарный вектор при достаточной разрядности сохраняет всю информацию о входящих в него понятиях.
Суммарный вектор имеет высокую разрядность и содержать большое количество единиц. Уменьшить количество единиц и понизить разрядность суммарного вектора можно, рассчитав для него хеш-функцию.
Идентификатор воспоминания, как и само описание, может состоять из нескольких понятий. Тогда и для него можно вычислить соответствующий хеш. При запоминании могут использоваться не исходные коды, а полученные хеши.
Ранее было показано, что для реального мозга возможными кандидатами на роль элементов клеточного автомата могут являться веточки дендритных деревьев нейронов.
Вычисление нейронами хеш-функции дендритных сигналов
Дендритные веточки образуют дендритное дерево (рисунок ниже). Веточки имеют только парные ветвления и не образуют замкнутых циклов.

Модели реальных нейронов (проект EyeWire)
Спайк нейрона возникает, когда деполяризация мембраны на его теле в районе аксонного холмика достигает критического значения. Деполяризация сомы, то есть тела нейрона, происходит, в основном, за счет сигналов от дендритных веточек. Потенциально такими сигналами могут быть токи, возникающие в дендритных веточках и дендритные спайки.
За счет того, что на пути к соме сигналы разных веточек в местах ветвления взаимодействуют между собой, сигналы, доходящие то тела нейрона, оказывается некими функциями сигналов дендритных веточек. Сам мембранный потенциал тела нейрона оказывается функцией сигналов всех веточек его дендритного дерева. Это, кстати, не особо противоречит классическому представлению о формальном нейроне. С той оговоркой, что классический формальный нейрон является простым пороговым сумматором сигналов на синапсах, а мы говорим о достаточно хитрой функции сигналов дендритных веточек.
В описанной трактовке спайк одного нейрона можно смело назвать бинарным результатом хеш-преобразования над сигналами его дендритных веточек. Таким образом, можно говорить о том, что вся картина активности нейронов может быть истолкована, как результат хеш-преобразования активности дендритных сегментах.
Для активации нейрона вся активность, возникающая на дендритном дереве должна уложиться в небольшой временной интервал, составляющий несколько миллисекунд. Если предположить, что за такой интервал и формируется суммарная картина дендритной активности, возникающая после прохождения всех волн сложного описания, то спайки нейронов идеально подходят на роль хеша, связанного с картиной, возникшей на дендритных сегментах.
Для запоминания через интерференцию паттернов необходимо две волны: волна идентификатора и волна значения, то есть сохраняемой информации. В реальной коре эти волны могут распространяться одновременно. При этом, идентификатор сам может быть достаточно сложным описанием. Можно предположить, что в коре хеш для информации и хеш для идентификатора могут формироваться одновременно, но разными нейронами. В принципе, эти нейроны могут быть нейронами разных типов. Самые распространенные нейроны коры – это пирамидальные и звездчатые нейроны. Может так оказаться, что, например, активность пирамидальных нейронов кодирует хеш-функцию для информации, а активность звездчатых хеш-функцию для идентификаторов воспоминаний.
Избранные точки дендрита
Мы пришли к тому, что в каждом месте коры текущая информация может кодироваться сочетанием активности расположенных в этом месте нейронов. Совокупную мгновенную картину их спайков при этом можно воспринимать, как хеш-функцию от соответствующей этим нейронам дендритной активности.
В клеточном автомате для запоминания требовалось, чтобы каждый элемент автомата видел и мог запомнить фрагмент хеш-кода, достаточной длины. Волна идентификатора указывала, какие элементы должны запоминать, а серия информационных волн формировала суммарную картину активности элементов из которой получался тот самый хеш-код, который они должны были запоминать.
В аналогии с мозгом это означает, что каждая дендритная веточка должна видеть активность окружающих ее нейронов и должна иметь возможность ее избирательно запомнить.
Если учесть, что мы хотим, чтобы одна веточка могла запомнить не одну и не две, а тысячи или миллионы различных комбинаций активности нейронов, то задача становится очень интересной.
До сих пор мы, в основном, говорили о дендритных деревьях нейронов, теперь посмотрим на их аксоны. Итак, основной процент нейронов коры приходится на пирамидальные и звездчатые нейроны. Для аксонов этих нейронов характерны сильно ветвящиеся коллатерали. Большая часть синаптических контактов аксона приходится на объем, размеры которого сопоставимы с размером дендритного дерева (рисунок ниже). Такая геометрия аксона обеспечивает то, что сигнал об активности нейрона становится доступен практически всем дендритным веткам этого и других нейронов, находящимся в некоторой окрестности (радиусом порядка 50-70 мкм) этого нейрона.
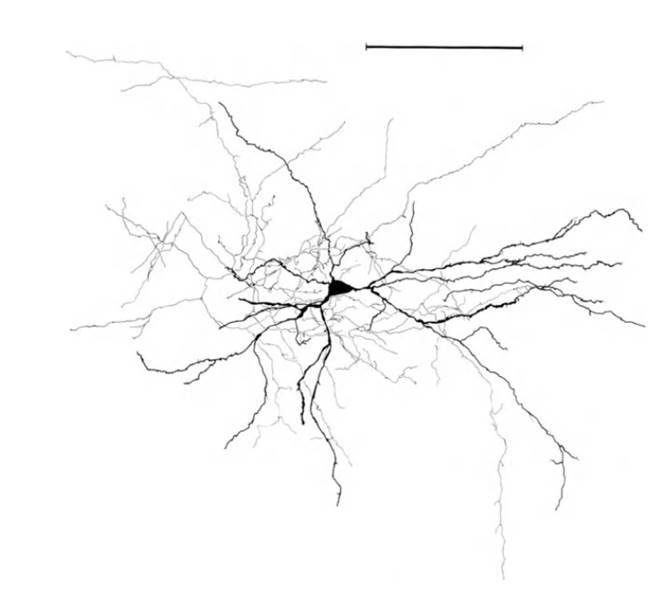
Структура звездчатого нейрона, линейка – 0.1 мм (Braitenberg, 1978)
Доступность сигнала следует понимать в том смысле, что для каждой дендритной ветки поблизости он нейрона будет место где вблизи от нее пройдет аксон этого нейрона. Соответственно, в момент активности нейрона по его аксону распространяется спайк и из всех синапсов, образуемых аксоном, высвобождаются нейромедиаторы. Часть этих нейромедиаторов за счет спиловера, то есть выброса за пределы синапса, может достичь требуемой дендритной ветки.
Вообще, аксоны могут распрстраняться далеко по коре или за ее пределы. Но основное ветвление аксона в большинстве случаев приходится на пространство окружающее сам нейрон. Среднее расстояние между синапсами на дендрите 0.5 мкм. Среднее расстояние между синапсами на аксоне 5 мкм. Число контактов на дендритах равно числу контактов на синапсах. Соответственно, суммарная длина аксона в 10 раз больше суммарной длины дендрита. На ближайшее окружающее нейрон пространство приходится около 6000 его синапсов. Это соответствует длине аксона в 3 сантиметра. Теперь, представьте, что эти 3 сантиметра уложены в сферу радиусом менее одной десятой миллиметра и вы получите представление о характере ветвления аксона. Рядом с любым сегментом дендрита проход множество аксонов соседних нейронов, причем некоторые приближаются к нему не по одному разу.
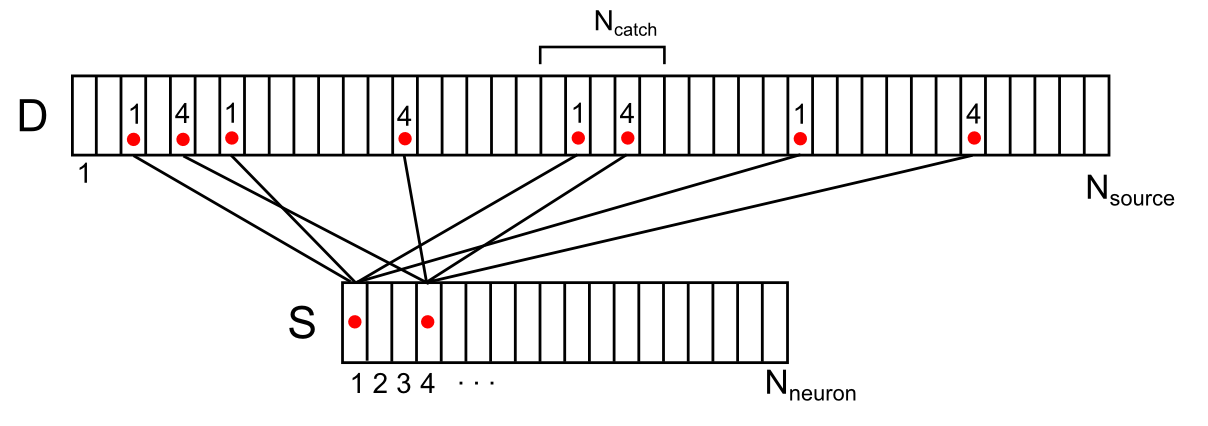
Синапсы непосредственно окружающие дендритную ветку, как собственные, так и просто, находящиеся рядом, являются источниками внесинаптических нейромедиаторов для этой ветки. Изобразим расположение этих источников вдоль условного сегмента дендрита (рисунок ниже). Для этого совместим источники с дендритом, приблизительно соблюдая их положение по длине дендрита. Занумеруем окружающие дендрит нейроны. Тогда для каждого источника на этой ветке можно указать номер нейрона из окружающего пространства, управляющего этим источником. Каждый из нейронов окружения может иметь сразу несколько контролируемых им источников, случайным образом распределенных по дендриту. Обозначим соотнесение нейронов и источников на дендритной ветке вектором D с элементами di, где каждый элемент – номер нейрона, управляющего источником.

Соотнесение окружающих нейронов и их контактов на дендрите
Обозначим Nneuron – количество нейронов окружения и Nsource – количество источников для одного сегмента дендрита.
Если мы зададимся расстоянием, на которое распространяются нейромедиаторы после спиловера, то сможем определить какие синапсы способны влиять на выбранное место дендрита. Обозначим через Ncatch количество источников, способных влиять на выбранное место дендрита. Для этих источников такое место можно назвать «ловушкой».
Теперь предположим, что несколько нейронов из окружения дендрита дали спайки. Это можно трактовать как сигнал, доступный для наблюдения нашему дендритному сегменту. Обозначим Nsig – количество активных нейронов, создающих информационный сигнал. Запишем этот сигнал бинарным вектором S размерности Nsource.
Для всех позиций на дендрите, кроме самых крайних, можно определить количество активных источников (плотность сигнала), попадающих в ловушку, по формуле

Например, для сигнала, показанного на рисунке ниже, плотность сигнала в отмеченной синаптической ловушке составит 2 (сумма сигналов от 1-го и 4-го нейронов).
Отображение активности двух нейронов окружения на дендритный сегмент (показана только часть связей и нумерации)
Для любого произвольного сигнала можно рассчитать какое распределение плотности он создаст на дендрите. Эта плотность будет колебаться в диапазоне от 0 до Ncatch. Максимальное значение будет достигаться, когда будут активны все источники, образующие соответствующую ловушку.
Воспользуемся значениями, характерными для реальной коры крысы (Braitenberg V., Schuz A., 1998) и, исходя из них, выберем ориентировочные параметры модели:
Длина секции = 150 мкм (300 синапсов, в среднем 0.5 мкм между синапсами)
Радиус плотного ветвления дендрита = 70 мкм
Плотность нейронов в коре = 9×104/мм3
Количество нейронов в окружении дендрита (Nneuron) = 100
Количество источников для дендритного сегмента (Nsource) = 3000
Размеры ловушки (Ncatch) = 15
Будем полагать, что сигнал кодируется активностью, например, 10% нейронов, тогда
Nsig = 10
Можно рассчитать вероятность того, что для произвольного сигнала, состоящего из Nsig единиц, найдется хотя бы одно место на дендритном сегменте, где сигнальная плотность составит ровно K единиц. Для приведенных параметров вероятность принимает следующие значения:
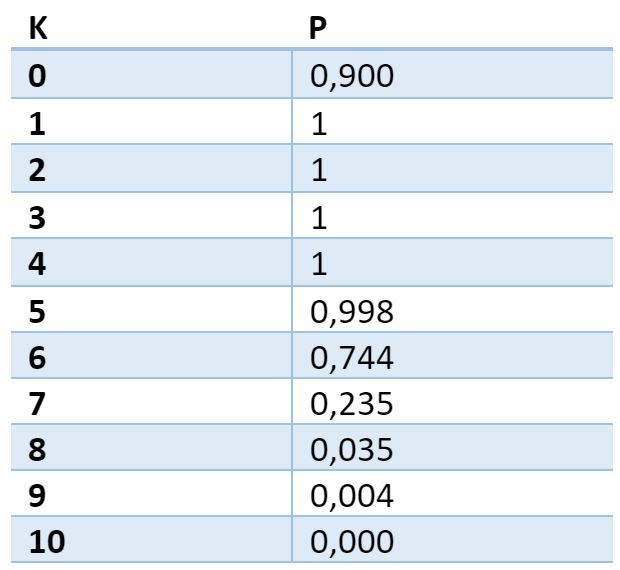
Таблица вероятности нахождения хотя бы одной ловушки с заданной плотностью. Первый столбец – требуемое количество активных источников в ловушке. Второй – вероятность нахождения на дендрите хотя бы одного места, где будет именно такое количество активных источников
Из таблицы видно, что с вероятностью близкой к 1 на любом дендритном сегменте для любого выбранного объемного сигнала найдется место, в котором сойдутся как минимум 5 аксонов активных нейронов. Это место дендрита можно считать избранным по отношению к выбранному сигналу. Если в этом месте запомнить какие именно аксоны (синапсы) были активны, то это позволит в последствии с высокой точностью детектировать повторение того-же сигнала.
Точность детектирования определяется вероятностью коллизий, то есть вероятностью того, что в каком-либо другом объемном сигнале окажутся активны те же нейроны, аксоны которых сошлись в избранном месте. То есть если, например, сигнал определяется активностью 10 нейронов из 100 и в избранном месте было зафиксировано сочетание 5 из этих 10 нейронов, то для коллизии достаточно, чтобы в каком-либо другом сигнале именно эти 5 нейронов также оказались активны.
Обозначим количество нейронов «пойманных» в ловушку, то есть активность которых совпала в избранном месте, через K. Чтобы другой сигнал, состоящий из Nsig активных нейронов, вызвал ошибочное узнавание надо, чтобы К его нейронов совпали с нейронами ловушки. Число таких совпадающих сигналов –  . Общее количество возможных сигналов —
. Общее количество возможных сигналов — 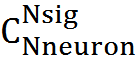 , вероятность ошибки
, вероятность ошибки

Для нашей модели при K=5 вероятность ошибки детектирования составит 3,34×10-6, при K=6, соответственно, вероятность ниже — 1,76×10-7.
Кодирование сигнала в избранном месте сочетанием нейромедиаторов
Каждое место дендрита окружено синапсами, как собственными, так и соседних дендритов. Эти синапсы являются источниками внесинаптических нейромедиаторов. Те из синапсов, что способны влиять на выбранное место дендрита образуют ловушку. Среднее количество таких синапсов для произвольного места дендрита — Ncatch. Зададимся значением K, определяющим то, сколько синапсов должно быть активно, чтобы считать место избранным по отношению к объемному сигналу. Видно, что для каждого места дендрита будет существовать достаточно много сигналов, создающих в этом месте минимум K пересечений. Чтобы мы могли с высокой точностью отследить повторение искомого сигнала необходимо не просто зафиксировать факт того, что в избранном по отношению к этому сигналу месте сработало К источников, а надо еще убедиться, что это именно те источники, что соответствуют сигналу. То есть надо по выбросу нейромедиаторов понять не просто сколько синапсов активировалось, а определить какие именно синапсы сработали на этот раз.
Как мы уже говорили, для большинства синапсов, в момент активности выделяется один «основной» нейромедиатор и в дополнение к нему один нейропептид или более ( Lundberg, J. M. 1996. Pharmacol. Rev. 48: 113-178.) (Bondy, C.A., et al. 1989. Cell. Mol. Neurobiol. 9: 427-446). Факт наличия у нейронов мозга большого количества нейромедиаторов и нейромодуляторов позволяет предположить, что основная функция такого многообразия – это создание в момент синхронной активности нейронов в каждом месте пространства уникальных сочетаний медиаторов и модуляторов. Можно предположить, что дополнительные вещества в синаптических везикулах распределены по синапсам так, чтобы обеспечивать в каждом месте пространства максимально разнообразие распределение. Если это так, то детектирование определенного сочетания активности синапсов сводится к определению соответствующего этим синапсам уникального набора испускаемых веществ.
Таким образом, если в избранном по отношению к определенному сигналу месте дендрита поместить детектор, чувствительный к сочетанию веществ, характерному для этого сигнала, то срабатывание этого детектора будет с очень высокой вероятностью говорить о повторении исходного сигнала.
Теперь мы знаем, что на каждой дендритной веточке всегда найдется место, которое окажется избранным по отношению к любому сигналу окружающих нейронов. Нам осталось понять, как веточка дендрита может запомнить, что это ее избранное место должно реагировать на определенное сочетание нейромедиаторов.
Рецепторы нейронов, как элементы памяти
Описывая работу нейрона, мы говорили, что окружающие вещества влияют на работу нейрона через его рецепторы. Рецепторы бывают ионотропные и метаботропные. Ионотропные рецепторы связываются с нейромедиаторами, высвобождающимися в синапсе, что вызывает изменение их конформации. Конформация молекулы – это изменение ее пространственной структуры без изменения самого состава молекулы.
Ионотропные рецепторы одновременно являются ионными каналами. Изменение конформации открывает ионный канал рецептора, что приводит к перемещению ионов и, соответственно, изменению мембранного потенциала.
Метаботропные рецепторы не имеют ионных каналов и действуют иначе. Той частью, что находится внутри нейрона они связаны с, так называемыми, G-белками. Когда эти рецепторы взаимодействуют со своими сигнальными веществами их конформация меняется и G-белок отпускается. Это ведет к разным возможным последствиям. Одно из возможных последствий – это открытие G-белками соседних ионных каналов, которые достаточно быстро меняют локальный мембранный потенциал нейрона. Что, в свою очередь, вызывает токи в дендрите и может стать причиной дендритного спайка.
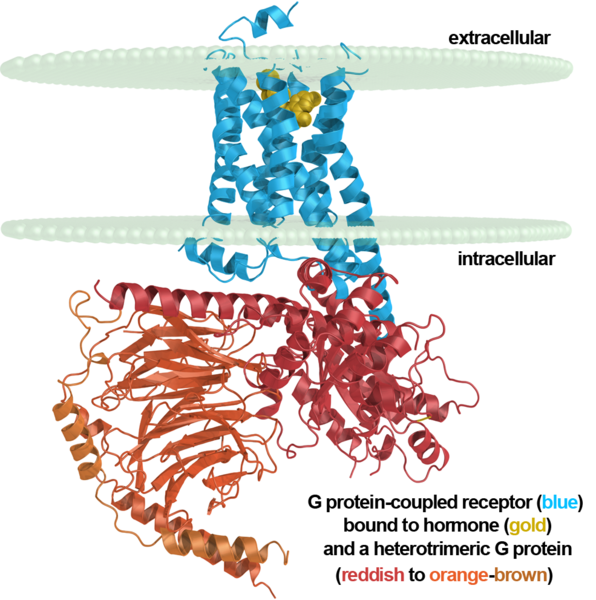
Метаботропный рецептор, мембрана нейрона и G-белок
Метаботропные рецепторы, в основном, располагаются вне синапсов и являются мишенями именно для внесинаптических нейромедиаторов. Рецепторы, в основном, объединяются в кластеры и действуют совместно. Кластеры метаботропных рецепторов, по сути, являются разнообразными детекторами, настроенными на определенные сочетания нейромедиаторов.
Кластеры метаботропных рецепторов очень подходят на роль элементов энграммы для нашей модели. Рядом с каждым синапсом могут находиться сотни таких кластеров. В них заранее случайным образом составлены сочетания рецепторов, потенциально чувствительные ко многим возможным в этом месте комбинациям внесинаптических нейромедиаторов, то есть таких нейромедиаторов, которые могут выделиться во внешнюю среду из соседних синапсов.
То есть расположенные в большом количестве в каждом месте дендрита метаботропные рецепторы могут являться «заготовками» для будущих энграмм. Переход «заготовок» в энграммы можно описать так. Предположим, что кластеры рецепторов, относящиеся к какой-либо дендритной ветке, изначально неактивны и никак не влияют на ее работу. Когда этой ветке требует произвести запоминание он дает знать об этом всем метаботропным рецепторам. Таким сигналом, например, может быть небольшая общая деполяризация мембраны этой ветки. Как мы говорили выше, где-то на этой ветке обязательно найдется избранное место. То есть место, где рядом с этой веточкой окажется сразу несколько активных синапсов. Если в этом месте веточки найдется кластер с рецепторами, чувствительность которого совпадет с коктейлем нейромедиаторов, то этот кластер должен будет перейти в активное состояние и впоследствии всегда реагировать на появление своего коктейля. Если до этого кластер не влиял на работу веточки дендрита, то теперь он должен будет при появлении своего коктейля создавать возбуждающий постсинаптический потенциал.
Оказывается, что метаботропные рецепторы, действительно умеют так себя вести. Определенные обстоятельства могут погружать наружные и внутренние части рецептора встречно в мембрану, лишая рецептор чувствительности. Или, наоборот, чувствительные части рецептора могут быть вытолкнуты из мембраны. В таком чувствительном состоянии рецептор может пребывать какое-то время, затем он может вернуться в исходное состояние – это соответствует краткосрочной памяти. Рецептор может и надолго зафиксировать такое чувствительное состояние. Если складываются подходящие условия, то начинаются процессы адгезии и полимеризации, которые могут оставить рецептор в чувствительном состоянии на дни и недели. Если процесс фиксации, который длится, предположительно, около месяца не прервется, то состояние рецептора зафиксируется навечно или правильнее сказать пожизненно. Все это соответствует различным стадиям консолидации долгосрочной памяти.
Механизмы, которые управляют таким поведением метаботропных рецепторов подробно изучил и описал А. Н. Радченко (Информационные механизмы мозга, 2007). Кстати, Радченко первый высказал предположение, что именно кластеры метаботропных рецепторов с их конформационными переходами и есть элементы энграмм.
Для памяти, основанной на пластичности синапсов, емкость считается достаточно просто. Пример такого расчета приведен на заглавной картинке. Заметим, что в нашей модели емкость памяти на дендритах, где-то в 1000 раз больше. И это еще не все.
В той пространственной структуре, что создается переплетением аксонов и дендритов работает идеология «избранных мест». То есть, чтобы рецепторы “были в деле” им необязательно относиться к тому дендриту, которому принадлежит синапс, через который передается сигнал. За счет того, что нейромедиаторы распространяются в межклеточном пространстве, “в деле” могут быть любые рецепторы, просто, геометрически находящиеся поблизости. Причем это, вообще, не обязательно должны быть рецепторы, принадлежащие именно нейронам.
Так, глиальные клетки коры, плазматические астроциты (рисунок ниже), имеют наборы тех же рецепторов, что и нейроны, а значит могут быть участниками механизмов памяти.

Плазматический астроцит
В коре количество астроцитов превышает количество нейронов. Астроциты коры имеют короткие ветвящиеся отростки. Этими отростками они охватывают находящиеся поблизости синапсы (рисунок ниже).
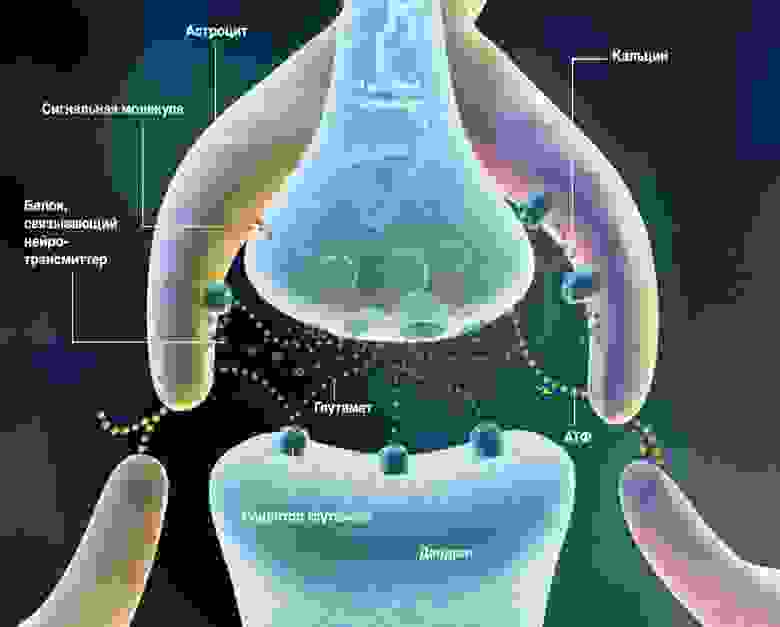
Трехсторонний синапс (R. D. Fields, B. Stevens-Graham, 2002)
Астроциты способны как усиливать реакцию синапса за счет выброса соответствующего медиатора, так и ослаблять ее за счет его поглощения или выброса связывающих нейромедиатор белков. Кроме того, астроциты способны выделять сигнальные молекулы, регулирующие выброс нейромедиатора аксоном. Концепция передачи сигналов между нейронами, учитывающая влияние астроцитов, называется трехсторонним синапсом (R. D. Fields, B. Stevens-Graham, 2002). Возможно, что трехсторонний синапс – это основной элемент, реализующий механизмы взаимной работы различных систем памяти.
Роль гиппокампа. Информация в идентификаторах. Кольцевые идентификаторы
В описываемой модели памяти чтобы на какой-либо зоне коры могли формироваться воспоминания кроме самой информационной картины на нее должны подаваться сигналы идентификаторов воспоминаний. Так как зоны коры выполняют различные функции, то уместно предположить, что и идентификаторы воспоминаний различаются для разных зон или групп зон.
Некоторые известные участки мозга в силу своей специфики хорошо подходят на роль источников идентификаторов. Так, верхние бугорки четверохолмия могут быть источником идентификаторов для зрительных зон. Нижние бугорки четверохолмия подходят на роль генераторов идентификаторов для слуховых зон коры. Наиболее показательный орган, связанный с памятью, – это гиппокамп, который хорошо подходит на роль генератора ключей воспоминаний для зон префронтальной коры.
В 1953 году пациенту, которого принято называть H.M. (Henry Molaison), при попытке вылечить эпилепсию было произведено двустороннее удаление гиппокампа (W. Scoviille, B. Milner, 1957). В результате у H.M. полностью исчезла способность что-либо запоминать. Он помнил все, что было с ним до операции, но что-либо новое вылетало у него из головы, как только переключалось его внимание. Кто смотрел фильм «Помни» («Memento») Кристофера Нолана, тот хорошо поймет, о чем разговор.

Henry Molaison
Случай H.M. достаточно уникален. В других случаях, связанных с удалением гиппокампа, где не было такого полного двустороннего разрушения, как у H.M., нарушения памяти либо были не столь ярко выражены, либо отсутствовали вовсе (W. Scoviille, B. Milner, 1957).
Полное удаление гиппокампа делает невозможным формирование новых воспоминаний. Нарушения в работе гиппокампа могут привести к синдрому Корсакова, который так же сводится к невозможности фиксировать текущие события, при сохранении старой памяти.
Достаточно распространенное представление о роли гиппокампа сводится к тому, что гиппокамп – это место хранение текущих воспоминаний, которые впоследствии перераспределяются по пространству коры. В описываемой модели роль гиппокампа иная, он создает уникальные ключи воспоминаний.
Созданные гиппокампом ключи через систему проекций распространяются на соответствующие зоны коры. Интерференция идентификаторов гиппокампа и информационных описаний создает память. При этом память сразу образуется «на своем месте» и не перемещается между гиппокампом и корой. Такое представление хорошо согласуется с экспериментальными данными. Действительно, удаление гиппокампа делает невозможным новое запоминание, так как исчезает генератор ключей воспоминаний. Старые воспоминания остаются нетронутыми, так как уже не зависят от гиппокампа. Их идентификаторы могут быть извлечены и использованы без задействования гиппокампа.
Но главные доводы в пользу предлагаемой роли гиппокампа связаны с функциями, которые обнаружены в гиппокампе и на первый взгляд не имеют прямого отношения к механизму памяти.
В 1971 году Джон О‘Киф открыл в гиппокампе клетки места (O’Keefe J., Dostrovsky J., 1971). Эти клетки реагируют подобно внутреннему навигатору. Если крысу поместить в длинный коридор, то по активности определенных клеток можно будет точно сказать, в каком его месте она находится. Причем реакция этих клеток не будет зависеть от того, как она попала в это место.
В 2005 году в гиппокампе были обнаружены нейроны, кодирующие положение в пространстве, образующие нечто наподобие координатной сетки (Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I., 2005).
В 2011 году оказалось, что в гиппокампе есть клетки, которые определенным образом кодируют временные интервалы. Их активность образует ритмические узоры, даже если ничего другого вокруг не происходит (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011).
Хранение данных в виде пар «ключ-значение» создает ассоциативный массив. В ассоциативном массиве ключ имеет двойную функцию. С одной стороны, он является уникальным идентификатором, который позволяет отличить одну пару от другой, с другой стороны, сам ключ может нести информацию сильно облегчающую поиск. Например, файловая система компьютера может рассматриваться как ассоциативный массив. Значение – это информация, хранящаяся в файле, ключ – информация о файле. Информация о файле – это путь, указывающий на место хранения, имя файла, дата создания. Для фотографий дополнительна информация – геотеги, координаты места, где сделан снимок. Для музыкальных файлов – название альбома и имя исполнителя. Все эти данные о файлах образуют составные сложные ключи, которые не только однозначно идентифицируют файлы, но и позволяют вести поиск по любому из полей ключа или любому их сочетанию. Чем подробнее составлен ключ, тем гибче оказываются возможности поиска.
Так как мозг реализует те же информационные задачи, что и компьютерные системы, то логично предположить, что хранение мозгом данных в виде пар «ключ-значение» будет сопровождаться создание ключей наиболее удобных для поиска. Для воспоминаний, с которыми имеет дело человек разумный набор дескрипторов ключа должен включать в себя:
- Указание места действия;
- Указание положения в пространстве;
- Указание времени события;
- Набор понятий, советующих основному смыслу происходящего. Некий аналог ключевых слов, описывающих содержание статьи.
Очень похоже, что гиппокамп не просто работает с местом, положением в пространстве и временем, а использует эти данные именно для составления сложных информационных ключей воспоминаний. По крайней мере, это очень хорошо объясняет, почему столь разнообразные функции сошлись в одном месте. Причем месте, непосредственно отвечающем, за формирование памяти.
Особый интерес представляет временное кодирование. Человеческая память не просто позволяет вспоминать статические картины, она способна воспроизводить последовательность сцен с сохранением их хронологии. Соответственно, в систему кодирования памяти должна быть заложена такая возможность. Было показано, что в гиппокампе есть клетки времени, создающие ритмические узоры (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011). Цикличность узоров наводит на мысль, что гиппокамп может использовать для создания временных полей идентификаторов событий те же приемы, что использует и человек для измерения времени.
Чтобы описать ход времени мы используем часы и календарь. В основе и того и другого лежат кольцевые идентификаторы. Минута состоит из 60 секунд. Это значит, что 60 идентификаторов последовательно сменяют друг друга, причем после 60 секунды опять следует первая. Аналогично с минутами в часе, часами в сутках, днями в месяцах, днями в неделях, месяцами в годах, годах в столетиях. То есть несколько кольцевых идентификаторов с разной периодичностью позволяют идентифицировать любой момент времени.

Похоже, что гиппокамп использует схожую систему временных кольцевых идентификаторов, что, собственно, и наблюдалось в опытах. Но, что особенно интересно, что такая система позволяет не только идентифицировать воспоминания, но и воспроизводить их последовательность. Если мы знаем правила следования идентификаторов друг за другом, то имея идентификатор одного события мы легко можем получить идентификатор события, следующего хронологически за ним и так далее.
Надо отметить, что кольцевые идентификаторы удобны не только для описания времени, но и для многих других ситуаций.
То, что мы описали в этой части, очень важно для понимания работы мозга, но не содержит, пока, никакой «магии». Да, это показывает где может находиться память и в каком виде она может храниться, но это не говорит ничего о том, как же мозг с легкостью решает задачи, непосильные компьютеру. Компьютер, конечно, может считать быстрее человека, но зато человек способен понимать смысл происходящего. В следующей части я расскажу, что вообще есть «смысл» и постепенно, станет понятно зачем нам так долго пришлось описывать работу нейронов и механизмы голографической памяти.
Алексей Редозубов
Логика сознания. Вступление
Логика сознания. Часть 1. Волны в клеточном автомате
Логика сознания. Часть 2. Дендритные волны
Логика сознания. Часть 3. Голографическая память в клеточном автомате
Логика сознания. Часть 4. Секрет памяти мозга
Логика сознания. Часть 5. Смысловой подход к анализу информации
Логика сознания. Часть 6. Кора мозга как пространство вычисления смыслов
Логика сознания. Часть 7. Самоорганизация пространства контекстов
Логика сознания. Пояснение «на пальцах»
Логика сознания. Часть 8. Пространственные карты коры мозга
Логика сознания. Часть 9. Искусственные нейронные сети и миниколонки реальной коры
Логика сознания. Часть 10. Задача обобщения
Логика сознания. Часть 11. Естественное кодирование зрительной и звуковой информации
Логика сознания. Часть 12. Поиск закономерностей. Комбинаторное пространство
Логика сознания. Часть 13. Мозг, смысл и конец света
![]()
Пирамидный нейрон коры головного мозга мыши, экспрессивный зеленый флуоресцентный белок (GFP)
Нейрон (от др.-греч. νεῦρον — волокно, нерв) — это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высокоспециализированная и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов.
Содержание
- 1 Обзор
- 2 Строение
- 2.1 Тело клетки
- 2.2 Дендриты и аксон
- 2.3 Синапс
- 3 Классификация
- 3.1 Структурная классификация
- 3.2 Функциональная классификация
- 3.3 Морфологическая классификация
- 4 Развитие и рост нейрона
- 5 Литература
- 6 См. также
- 7 Ссылки
- 8 Примечания
Обзор
Сложность и многообразие функций нервной системы определяются взаимодействием между нейронами, которое, в свою очередь, представляет собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд (потенциал действия), который движется по телу нейрона.
Строение
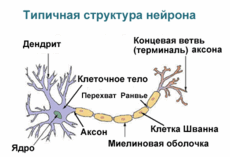
Тело клетки
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), снаружи ограничена мембраной из двойного слоя липидов (билипидный слой). Липиды состоят из гидрофильных головок и гидрофобных хвостов, расположены гидрофобными хвостами друг к другу, образуя гидрофобный слой, который пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: на поверхности (в форме глобул), на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый и сложный цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20-30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии. В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные(двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. эффектус — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон

![]()
Схема строения нейрона
Аксон — обычно длинный отросток, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
Классификация
Структурная классификация
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация
Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона;
- количество и характер ветвления отростков;
- длину нейрона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов. Длина нейрона у человека составляет около 150 мкм.
По количеству отростков выделяют следующие морфологические типы нейронов[1]:
- униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
- псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
- биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
- мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Развитие и рост нейрона
Нейрон развивается из небольшой клетки-предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным.) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Литература
| Нейрон на Викискладе? |
- Поляков Г. И., О принципах нейронной организации мозга, М: МГУ, 1965
- Косицын Н. С. Микроструктура дендритов и аксодендритических связей в центральной нервной системе. М.: Наука, 1976, 197 с.
- Немечек С. и др. Введение в нейробиологию, Avicennum: Прага, 1978, 400 c.
- Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение
- Мозг (сбоpник статей: Д. Хьюбел, Ч. Стивенс, Э. Кэндел и дp. — выпуск журнала Scientific American (сентябрь 1979)). М. :Миp, 1980
- Савельева-Новосёлова Н. А., Савельев А. В. Устройство для моделирования нейрона. А. с. № 1436720, 1988
- Савельев А. В. Источники вариаций динамических свойств нервной системы на синаптическом уровне // журнал “Искусственный интеллект”, НАН Украины. — Донецк, Украина, 2006. — № 4. — С. 323-338.
| |
|
|---|---|
| Нейроны (Серое вещество) |
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты) Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит, Базальный дендрит) типы: Биполярные нейроны · Псевдополярные нейроны · Мультиполярные нейроны · Пирамидальный нейрон · Клетка Пуркинье · Гранулярная клетка |
| Афферентный нерв/ Сенсорный нерв/ Сенсорный нейрон |
GSA · GVA · SSA · SVA · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено, II or Aβ, Aδ-волокна, C-волокна) |
| Эфферентный нерв/ Моторный нерв/ Моторный нейрон |
GSE · GVE · SVE · Верхний моторный нейрон · Нижний моторный нейрон (α мотонейроны, γ мотонейроны) |
| Синапс | Нейропиль · Синаптический пузырек · Нервно-мышечный синапс · Электрический синапс · Химический синапс · Интернейрон (Клетки Реншоу) |
| Сенсорный рецептор | Чувствительное тельце Мейснера · Нервное окончание Меркеля · Тельца Пачини · Окончание Руффини · Нервномышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица |
| Нейроглия | Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия |
| Миелин (Белое вещество) |
CNS: Олигодендроцит PNS: Клетки Шванна · Невролемма · Перехват Ранвье/Межузловой сегмент · Насечка миелина |
| Соединительная ткань | Эпиневрий · Периневрий · Эндоневрий · Нервные пучки · Мозговые оболочки: твёрдая, паутинная, мягкая |
См. также
- Компьютер
- Искусственный нейрон
- Модель биологического нейрона
- Нейронная сеть
- Нейрофизиология
- Зеркальные нейроны
Ссылки
- Научно-популярный фильм «Нервная клетка»
Примечания
- ↑ см. рuc.
|
|
В этой статье не хватает ссылок на источники информации.
Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена. |
Введение в нейробиологию
7. Нейроны
Основными структурными особенностями нейронов являются перикария, дендриты и аксоны Нейроны содержат те же самые внутриклеточные компоненты, что и другие клетки Молекулярные маркеры могут использоваться для идентификации нейронов
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 85—86 миллиардов нейронов[1][2]. Нейроны могут соединяться один с другим, формируя биологические нейронные сети. Нейроны разделяют на рецепторные, эффекторные и вставочные.
Сложность и многообразие функций нервной системы определяются взаимодействием между нейронами. Это взаимодействие представляет собой набор различных сигналов, передаваемых между нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов. Ионы генерируют электрический заряд (потенциал действия), который движется по телу нейрона.
Важное значение для науки имело изобретение метода Гольджи в 1873 году, позволявшего окрашивать отдельные нейроны[3][4]. Термин «нейрон» (нем. Neuron) для обозначения нервных клеток введён Г. В. Вальдейером в 1891 году[5][6].

Строение нейронов
Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20—30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии.(Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в среднем в 10—50 раз больше, чем нейронов).
В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Аксоны и дендриты

Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу. Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторнойклеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г.