Механизм возникновения и проведения нервного импульса
Возникновение
нервного импульса связано с состоянием
мембраны нейрона. Биологические мембраны,
будучи липидно-белковым комплексом,
непроницаемы для заряженных ионов.
Перемещение ионов через мембрану
осуществляется с помощью специальных
транспортных систем: натриевого насоса,
работающего с затратой энергии (Na+,
К+-АТФ-аза)
и ионпроводящих каналов – натриевых и
калиевых.
В состоянии
физиологического покоя по разные стороны
мембраны имеет место разность концентраций
ионов, главным образом Na+
и К+.
Концентрация ионов К+
выше в
клетке,
ионов Na+
– в межклеточной жидкости. Это влияет
и на распределение других ионов. Разность
концентраций ионов поддерживается с
помощью натриевого насоса, перекачивающего
ионы против градиента концентрации (в
противном случае диффузия ионов по
градиенту концентрации привела бы к
выравниванию концентраций ионов по обе
стороны мембраны и гибели клетки) Таким
образом, внутри аксона образуется
избыток (–) зарядов, снаружи – (+) зарядов,
то есть возникает разность электрических
потенциалов – потенциал покоя (ПП). Его
значение составляет – 60-70 мВ. ПП одинаков
по всей длине нервного волокна (рис. 4).
Раздражение
нервного волокна приводит сначала к
открытию Na+—
и К+—
каналов. Раньше открываются Na+-каналы,
и в клетку устремляется поток ионов
натрия. Это изменяет трансмембранный
потенциал: сначала он становится равным
0 (деполяризация мембраны), затем
происходит перезарядка мембраны,
внутренняя сторона мембраны приобретает
(+) заряд, наружная (-). Разность потенциалов
при этом достигает 40 мВ. Эта величина и
есть потенциал действия (ПД) (рис. 5).
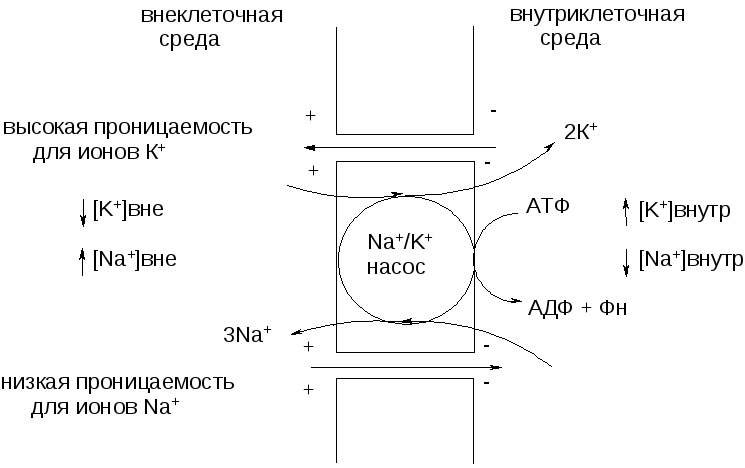
Рис. 4. Природа поляризации клеточной
мембраны
Натриевые каналы
закрываются, открываются калиевые,
начинается выход калия из клетки,
потенциал изменяется от – 40 мВ до – 70
мВ, то есть идет реполяризация мембраны.
После закрытия ионных каналов ионный
насос восстанавливает исходное
распределение ионов по сторонам мембраны.

Рис. 5. Потенциал действия
ПД развивается в
том случае, если под действием раздражителя
происходит изменение ПП до -50 мВ (пороговое
значение). Если действие раздражителя
не достигает этого значения, ПД не
генерируется (принцип «все или ничего»).
Движение пд по аксону
Потенциал действия,
возникая в одном участке аксона,
вследствие диффузии ионов вдоль нервного
волокна изменяет ПП в соседнем участке
и вызывает развитие ПД. Благодаря этому
ПД, возникнув в одном месте, проходит
весь аксон и достигает следующей клетки.
В таком качестве ПД называется нервным
импульсом.
В миелиновом
волокне Na+—
и К+—
каналы располагаются в перехватах
Ранвье, где мембрана аксона контактирует
с межклеточной жидкостью. Поэтому
распространение нервного импульса идет
скачками; скорость распространения
импульса в 5-6 раз выше, чем в безмиелиновом
волокне, где ПД распространяется плавно.
Вся деятельность
нервной системы строится на основе
рефлексов, материальной основой которых
являются рефлекторные дуги. Рефлекторной
дугой
называется путь, состоящий из цепей
нейронов, по которому нервный импульс
проходит от чувствительных нервных
клеток до рабочего органа. Рефлекторные
дуги могут быть простыми или сложными.
Простая рефлекторная дуга состоит из
двух нейронов – чувствительного и
двигательного (эффекторного), между
которыми один синапс.
Примерами простых
рефлекторных дуг являются дуги сухожильных
рефлексов (коленного, ахиллова,
подошвенного, сгибательного и
разгибательного локтевого, брюшного),
которые замыкаются на уровне спинного
мозга.

Рис. 6. Рефлекторная дуга кожного рефлекса
Рефлекторные дуги
большинства рефлексов включат помимо
чувствительного и эффекторного еще
несколько вставочных нейронов. Посредством
вставочных нейронов осуществляется
контроль адекватности ответа рабочего
органа полученному раздражению (рис.
6).
Вегетативные
рефлекторные дуги являются кольцевыми,
от рабочего органа есть обратная связь
в ЦНС для регуляции рефлекторной реакции.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Нервные импульсы распространяются при перемещении ионов через мембрану нервной клетки и передаются из одной нервной клетки в другую с помощью нейромедиаторов.
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
См. также:
Краткое описание:
Библиографическая ссылка для цитирования: Сазонов В.Ф. 2_3 Потенциал действия и нервный импульс [Электронный ресурс] // Кинезиолог, 2009-2021: [сайт]. Дата обновления: 21.12.2021. URL: http://kineziolog.su/content/23-potentsial-deistviya-i-nervnyi-impuls (дата обращения: __.__.20__).
__________________Понятие потенциала действия и нервного импульса. Описание графика потенциала действия и механизмов нервного импульса. «Нервный импульс — это волна изменений, движущаяся по мембране нейрона». © 2012-2021 Сазонов В.Ф. © 2012-2021 kineziolog.su
Нервный импульс
Разбираясь в нервных импульсах, мы будем иметь в виду нервное возбуждение, бегущее (=распространяющееся) по мебране нейрона. Строго говоря, движущееся по нейронам и нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия, хотя в физиологической литературе два этих понятия обычно используют как синонимы.
Для того чтобы произвести нервный импульс, нейрон сначала должен создать состояние готовности (потенциал покоя), затем предготовности (локальный потенциал), и, наконец, при достижении порогового уровня локального потенциала (критического уровня деполяризации) — породить бегущий по мембране нервный импульс.

Нервный импульс — это движущаяся волна изменений в состоянии мембраны. Волна включает в себя три вида изменений: структурные (открытие и закрытие мембранных ионных каналов); химические (трансмембранные потоки ионов); электрические (изменения электрического потенциала мембраны) Электрических изменений тоже три: деполяризация, позитивная поляризация (=инверсия, =овершут) и реполяризация. Электрические изменения (проявления) в состоянии мембраны — это потенциал действия. © 2012-2021 Сазонов В.Ф. © 2012-2021 kineziolog.su
Можно сказать короче:
«Нервный импульс — это волна изменений, движущаяся по мембране нейрона». © 2012-2021 Сазонов В.Ф. © 2012-2021 kineziolog.su
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин «потенциал действия». Хотя потенциал действия — это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Сравним два понятия: «нервный импульс» и «потенциал действия».
Нервный импульс — это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.

Потенциал действия — это только электрический компонент нервного импульса, характеризующий изменения электрического потенциала на локальном участке мембраны во время прохождения через него нервного импульса. Потенциал мембраны при этом изменяется от -70 до +30 мВ и обратно до -70 мВ — это и есть потенциал действия. (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик — главный породитель нервных импульсов
Аксонный холмик — это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) — и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим «начинаниям». Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они «перебиваются» потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала рецепторный потенциал, затем генераторный потенциал, а потом и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита)
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия — и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трёх сторон, на трёх уровнях:
-
Электрические явления – развитие потенциала действия.
-
Химические явления – движение ионных потоков.
-
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию, что означает уменьшение электроотрицательности на внутренней стороне мембраны. Сначала деполяризация иёет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением. Говоря образно, деполяризация так разгоняется, что в разгону перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация, т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:

Восходящая ветвь графика:
-
Потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ).
-
Нарастающий локальный потенциал – пропорциональная раздражителю деполяризация в интервале от –70 мВ до –50 мВ.
-
Критический уровень деполяризации (–50 мВ) запускает резкое ускорение деполяризации за счёт самораскрытия нового вида натриевых каналов (потенциал-управляемых), с этой точки начинается спайк – высокоамплитудная часть потенциала действия.
-
Самоусиливающаяся круто нарастающая деполяризация за счёт автоматического самораскрытия потенциал-управляемых натриевых каналов.
-
Переход нулевой отметки (0 мВ) – смена полярности мембраны. Но новых процессов это событие не вызывает.
-
«Овершут» – положительная поляризация (=инверсия, или =реверсия потенциала мембраны). Электроотрицательность переходит в электроположительность.
-
Пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия. Открытые потенциал-управляемые натриевые каналы к этому моменту самостоятельно автоматически закрываются изнутри специальными белковыми «пробками», и поступление положительно заряженных ионов натрия в клетку прекращается. Поэтому прекращается нарастание потенциала действия.
Нисходящая ветвь графика:
-
Реполяризация – восстановление прежней исходной электроотрицательности мембраны. Потенциал от +30 мВ опускается вниз. Это происходит благодаря утечке ионов калия из клетки через множество открытых калиевых каналов. Эти ионы выходят из клетки под действием химической силы, из-за разности их концентрации внутри и снаружи клетки. Внутри их много, а снаружи мало, вот они и перемещаются туда, где их мало.
-
Переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную. Но ничего принципиально нового здесь не происходит. Разве что с этого момента начинает действовать электрическая сила, затягивающая положительные ионы внутрь клетки и удерживающая те из них, которые уже находятся в клетке. Ионам калия с этого момента становится всё труднее выходить из клетки.
-
Переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости нейрона.
-
Следовые процессы (следовая деполяризация или следовая гиперполяризация).
-
Восстановление потенциала покоя – возврат мембраны к своему нормальному состоянию: –70 мВ.
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Образно можно сказать, что заряженные ионы – это и есть создатели электрических потенциалов в нервных клетках. Для многих людей звучит странно утверждение, что вода не проводит электрический ток. Но на самом деле это так. Сама по себе вода является диэлектриком, а не проводником. В воде электрический ток обеспечивают не электроны, как в металлических проводах, а заряженные ионы: положительные катионы и отрицательные анионы. В живых клетках основную «электрическую работу» выполняют катионы, так как они более подвижны. Электрические токи в клетках – это потоки ионов.
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками. Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы.
На молекулярно-структурном уровне происходит открытие мембранных натриевых каналов. Сначала этот процесс идет пропорционально силе внешнего воздействия, а затем становится просто «неудержимым» и массовым. Открытие каналов обеспечивает вход натрия в клетку и вызывает деполяризацию. Затем, примерно через 2-5 миллисекунд, происходит их автоматическое закрытие. Это закрытие каналов резко обрывает движение ионов натрия внутрь клетки, и, следовательно, обрывает нарастание электрического потенциала. Рост потенциала прекращается, и на графике мы видим спайк. Это вершина кривой на графике, дальше процесс пойдет уже в обратном направлении. Конечно, очень интересно разобраться в том, что натриевые каналы имеют двое ворот, и открываются они активационными воротами, а закрываются инактивационными, но это следует обсуждать ранее, в теме «Возбуждение». Мы на этом останавливаться не будем.
Параллельно в открытием натриевых каналов с небольшим отставанием во времени идет нарастающее открытие калиевых каналов. Они медлительные по сравнению с натриевыми. Открытие дополнительных калиевых каналов усиливает выход положительных ионов калия из клетки. Выход калия противодействует «натриевой» деполяризации и вызывает восстановление полярности (восстановление электроотрицательности). Но натриевые каналы опережают калиевые, они срабатывают примерно в 10 раз быстрее. Поэтому входящий поток положительных ионов натрия в клетку опережает компенсирующий выход ионов калия. И поэтому деполяризация развивается опережающими темпами по сравнению с противодействующей ей поляризацией, вызванной утечкой ионов калия. Вот почему, пока натриевые каналы не закроются, восстановление поляризации не начнется.
Пожар как метафора распространяющегося возбуждения
Для того чтобы перейти к пониманию смысла динамичного процесса возбуждения, т.е. к пониманию его распространения вдоль мембраны, надо представить себе, что описанные нами выше процессы захватывают сначала ближайшие, а затем все новые, все более и более отдаленные участки мембраны, пока не пробегут по всей мембране полностью. Если вы видели «живую волну», которую устраивают болельщики на стадионе за счет вставания и приседания, то вам легко будет представить себе мембранную волну возбуждения, которая образуется за счет последовательного протекания в соседних участках трансмембранных ионных токов.
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
Можно ли «погасить» волну распространяющегося возбуждения?
Вряд ли нервная клетка способна «погасить» этот начавшийся «пожар» возбуждения. Поэтому первый способ подходит только для искусственного вмешательства в работу нейронов (например, в лечебных целях). Но вот «залить водичкой» некоторые участки и поставить блок распространению возбуждения, оказывается, вполне возможно.
© Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии — деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью — невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Источник: Антонов В.Ф., 1996.
Смотрите также: Потенциал действия мотонейрона
Видео: Потенциал действия (Action potential)
© 2012-2019 Сазонов В.Ф. © 2016-2019 kineziolog.su.