Какими характеристиками обладают аксоны? Выберите все правильные ответы. Имеют короткие отростки
скопление отростков формирует белое вещество нервной системы
проводят нервный импульс со скоростью 120 м/с
скопление отростков формирует серое вещество нервной системы
имеют миелиновую оболочку
импульс по отростку всегда направляют к телу клетки
Предыдущий
вопрос
Следующий
вопрос
Длинная проекция на нейрон, который отводит сигналы
| Аксон | |
|---|---|
 Аксон многополярного нейрона Аксон многополярного нейрона |
|
| Идентификаторы | |
| MeSH | D001369 |
| Анатомическая терминология [редактировать в Викиданных ] |
аксон (от греческого ἄξων áxōn, ось) или нервное волокно (или нерв волокно : см. орфографические различия ), представляет собой длинный тонкий выступ нервной клетки или нейрона у позвоночных, который обычно проводит электрические импульсы, известные как потенциалы действия, от тела нервной клетки. Функция аксона — передавать информацию различным нейронам, мышцам и железам. В некоторых сенсорных нейронах (псевдоуниполярных нейронах ), таких как нейроны прикосновения и тепла, аксоны называются афферентными нервными волокнами, и электрический импульс проходит по ним от периферия к телу клетки и от тела клетки к спинному мозгу вдоль другой ветви того же аксона. Дисфункция аксонов является причиной многих наследственных и приобретенных неврологических расстройств, которые могут поражать как периферические, так и центральные нейроны. Нервные волокна классифицируются на три типа — нервные волокна группы A, нервные волокна группы B и нервные волокна группы C. Группы A и B являются миелинизированными, а группа C немиелинизированными. Эти группы включают как сенсорные волокна, так и двигательные волокна. Другая классификация группирует только сенсорные волокна как Тип I, Тип II, Тип III и Тип IV.
Аксон — это один из двух типов цитоплазматических выступов из тела клетки нейрона; другой тип — дендрит . Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно имеют постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты получают сигналы, тогда как аксоны передают их). Некоторые типы нейронов не имеют аксона и передают сигналы от своих дендритов. У некоторых видов аксоны могут исходить из дендритов, известных как дендриты, несущие аксоны. Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга.
Аксоны покрыты мембраной, известной как аксолемма ; цитоплазма аксона называется аксоплазмой. Большинство аксонов разветвляются, в некоторых случаях очень обильно. Концевые ветви аксона называются телодендриями. Набухший конец телодендрона известен как окончание аксона, которое соединяется с дендроном или телом клетки другого нейрона, образуя синаптическое соединение. Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсами. В некоторых случаях аксон одного нейрона может образовывать синапс с дендритами того же нейрона, что приводит к аутапсу. В синапсе мембрана аксона плотно прилегает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Некоторые синаптические соединения появляются по всей длине аксона по мере его расширения — они называются проходящими («проходящими») синапсами и могут быть сотнями или даже тысячами вдоль одного аксона. Другие синапсы выглядят как терминалы на концах аксональных ветвей.
Один аксон со всеми его ветвями, взятыми вместе, может иннервировать несколько частей мозга и генерировать тысячи синаптических окончаний. Пучок аксонов образует нервный тракт в центральной нервной системе и пучок в периферической нервной системе. У плацентарных млекопитающих самым большим белым веществом трактом в головном мозге является мозолистое тело, образованное примерно из 200 миллионов аксонов в человеческом мозге.
Содержание
- 1 Анатомия
- 1.1 Аксональная область
- 1.1.1 Аксональный бугор
- 1.1.2 Начальный сегмент
- 1.2 Аксональный транспорт
- 1.3 Миелинизация
- 1.4 Узлы Ранвье
- 1.5 Терминалы аксонов
- 1.1 Аксональная область
- 2 Потенциалы действия
- 3 Развитие и рост
- 3.1 Развитие
- 3.1.1 Внеклеточная передача сигналов
- 3.1.2 Внутриклеточная передача сигналов
- 3.1.3 Динамика цитоскелета
- 3.2 Рост
- 3.3 Регулирование длины
- 3.1 Развитие
- 4 Классификация
- 4.1 Двигательная
- 4.2 Сенсорная
- 4.3 Вегетативная
- 5 Клиническая значимость
- 6 Анамнез
- 7 Другие животные
- 8 См. Также
- 9 Ссылки
- 10 Внешние ссылки
Анатомия
 Типичный миелинизированный аксон
Типичный миелинизированный аксон  Рассеченный мозг человека, показывающий серое вещество и белое вещество
Рассеченный мозг человека, показывающий серое вещество и белое вещество
Аксоны являются основными линии передачи нервной системы, и в виде пучков они образуют нервы. Некоторые аксоны могут достигать одного метра и более, а другие — всего лишь один миллиметр. Самые длинные аксоны в человеческом теле — это аксоны седалищного нерва, которые проходят от основания спинного мозга до большого пальца каждой стопы. Диаметр аксонов также варьируется. Большинство отдельных аксонов имеют микроскопический диаметр (обычно около одного микрометра (мкм) в поперечнике). Самые большие аксоны млекопитающих могут достигать в диаметре до 20 мкм. Гигантский аксон кальмара, который специализируется на очень быстрой передаче сигналов, имеет диаметр, близкий к 1 миллиметру, то есть размер небольшого грифеля карандаша. Количество аксональных телодендрий (разветвляющихся структур на конце аксона) также может различаться от одного нервного волокна к другому. Аксоны в центральной нервной системе (ЦНС) обычно показывают множественные телодендрии с множеством синаптических конечных точек. Для сравнения, аксон гранулярных клеток мозжечка характеризуется одним Т-образным узлом ответвления, от которого отходят два параллельных волокна. Продуманное ветвление позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
В нервной системе есть два типа аксонов: миелинизированные и немиелинизированные аксоны. Миелин представляет собой слой жирового изолирующего вещества, которое образовано двумя типами глиальных клеток шванновских клеток и олигодендроцитов. В периферической нервной системе шванновские клетки образуют миелиновую оболочку миелинизированного аксона. В центральной нервной системе олигодендроциты образуют изолирующий миелин. Вдоль миелинизированных нервных волокон через равные промежутки времени возникают промежутки в миелиновой оболочке, известные как узлы Ранвье. Миелинизация обеспечивает особенно быстрый режим распространения электрического импульса, называемый скачкообразной проводимостью.
Миелинизированные аксоны от кортикальных нейронов образуют основную часть нервной ткани, называемую белым веществом в мозг. Миелин придает белый цвет ткани в отличие от серого вещества коры головного мозга, которое содержит тела нейрональных клеток. Подобное расположение наблюдается в мозжечке. Связки миелинизированных аксонов составляют нервные пути в ЦНС. Там, где эти тракты пересекают среднюю линию мозга и соединяют противоположные области, они называются комиссурами. Самым крупным из них является мозолистое тело, которое соединяет два полушария головного мозга, и у него около 20 миллионов аксонов.
Структура нейрона, как видно, состоит из двух отдельных функциональных областей или компартментов — тела клетки вместе с дендритами в качестве одной области и аксональной области в качестве другой.
Аксональная область
Аксональная область или компартмент включает бугорок аксона, начальный сегмент, остальную часть аксона и телодендрии аксона и терминалы аксона. Он также включает миелиновую оболочку. тельца Ниссля, которые продуцируют нейрональные белки, отсутствуют в аксональной области. Белки, необходимые для роста аксона и удаления отходов жизнедеятельности, нуждаются в транспортном каркасе. Этот аксональный транспорт обеспечивается в аксоплазме за счет расположения микротрубочек и промежуточных филаментов, известных как нейрофиламентов.
Аксональный холмик
 Деталь, показывающая микротрубочки на бугорке аксона и начальном сегменте.
Деталь, показывающая микротрубочки на бугорке аксона и начальном сегменте.
бугорок аксона — это область, образованная из тела клетки нейрона, которая расширяется, чтобы стать аксоном. Он предшествует начальному сегменту. Полученные потенциалы действия, которые суммируются в нейроне, передаются на бугорок аксона для генерации потенциала действия из начального сегмента.
Начальный сегмент
начальный сегмент аксона (AIS) представляет собой структурно и функционально отдельный микродомен аксона. Одна из функций начального сегмента — отделить основную часть аксона от остальной части нейрона; другая функция — помочь инициировать. Обе эти функции поддерживают нейрон клеточную полярность, в которой дендриты (и в некоторых случаях сома ) нейрона получают входные сигналы в базальной области, а в апикальной области — аксон нейрона обеспечивает выходные сигналы.
Начальный сегмент аксона немиелинизирован и содержит специализированный комплекс белков. Его длина составляет примерно от 20 до 60 мкм, и он функционирует как место инициации потенциала действия. Как положение на аксоне, так и длина AIS могут меняться, показывая степень пластичности, которая может точно настраивать нейрональный выход. Более длинный AIS связан с большей возбудимостью. Пластичность также проявляется в способности AIS изменять свое распределение и поддерживать активность нейронных схем на постоянном уровне.
AIS очень специализирован для быстрого проведения нервных импульсов. Это достигается за счет высокой концентрации потенциал-управляемых натриевых каналов в начальном сегменте, где возникает потенциал действия. Ионные каналы сопровождаются большим количеством молекул клеточной адгезии и каркасных белков, которые прикрепляют их к цитоскелету. Взаимодействие с анкирином G важно, поскольку он является основным организатором в AIS.
Аксональный транспорт
аксоплазма эквивалентна цитоплазма в ячейке. Микротрубочки образуются в аксоплазме на бугорке аксона. Они расположены по длине аксона в перекрывающихся участках и все направлены в одном направлении — к окончанию аксона. Об этом говорят положительные окончания микротрубочек. Такое перекрывающееся расположение обеспечивает маршруты транспортировки различных материалов из тела клетки. Исследования аксоплазмы показали движение многочисленных пузырьков всех размеров, которые можно увидеть вдоль цитоскелетных филаментов — микротрубочек и нейрофиламентов, в обоих направлениях между аксоном и его окончаниями и телом клетки.
Исходящий антероградный транспорт из тела клетки по аксону переносит митохондрии и мембранные белки, необходимые для роста, к концу аксона. Входящий ретроградный транспорт переносит отходы клетки от терминала аксона к телу клетки. Исходящие и входящие треки используют разные наборы моторных белков. Исходящий транспорт обеспечивается kinesin, а входящий обратный трафик обеспечивается dynein. Динеин направлен на минус-конец. Существует множество форм моторных белков кинезина и динеина, и считается, что каждая из них несет свой груз. Исследования транспорта в аксоне привели к названию кинезина.
Миелинизация
 TEM миелинизированного аксона в поперечном сечении.
TEM миелинизированного аксона в поперечном сечении.  Поперечное сечение аксона: (1) Аксон (2) Ядро (3) Шванновская клетка (4) Миелиновая оболочка (5) Неврилемма
Поперечное сечение аксона: (1) Аксон (2) Ядро (3) Шванновская клетка (4) Миелиновая оболочка (5) Неврилемма
В нервной системе аксоны могут быть миелинизированы, или немиелинизированные. Это обеспечение изолирующего слоя, называемого миелиновой оболочкой. Миелиновая мембрана уникальна своим относительно высоким отношением липидов к белку.
В аксоны периферической нервной системы миелинизируются глиальные клетки, известные как клетки Шванна. В центральной нервной системе миелиновая оболочка представлена другим типом глиальных клеток, олигодендроцитом. Клетки Шванна миелинизируют единственный аксон. Олигодендроцит может миелинизировать до 50 аксонов.
Состав миелина этих двух типов различается. В ЦНС основным белком миелина является протеолипидный белок, а в ПНС — основной белок миелина.
Узлы Ранвье
Узлы Ранвье (также известные как миелиновая оболочка промежутки) представляют собой короткие немиелинизированные сегменты миелинизированного аксона, которые периодически встречаются между сегментами миелиновой оболочки. Следовательно, в точке узла Ранвье аксон уменьшается в диаметре. Эти узлы являются областями, где могут быть созданы потенциалы действия. В скачкообразной проводимости электрические токи, возникающие в каждом узле Ранвье, передаются с небольшим затуханием к следующему узлу в линии, где они остаются достаточно сильными, чтобы генерировать другой потенциал действия. Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Терминалы аксона
Аксон может делиться на множество ветвей, называемых телодендриями (греч. Конец дерева). В конце каждого телодендрона находится окончание аксона (также называемое синаптическим бутоном или терминальным бутоном). Терминалы аксонов содержат синаптические пузырьки, в которых хранится нейромедиатор для высвобождения в синапсе. Это делает возможными множественные синаптические связи с другими нейронами. Иногда аксон нейрона может синапсировать с дендритами того же нейрона, когда это известно как аутапс.
Потенциалы действия
| Структура типичного химического синапса |
|---|
 Постсинаптическая. плотность Напряжение -. закрытый Ca. канал Синаптический. везикула Нейротрансмиттер. транспортер Рецептор Нейротрансмиттер Аксонный терминал Синаптическая щель Дендрит Постсинаптическая. плотность Напряжение -. закрытый Ca. канал Синаптический. везикула Нейротрансмиттер. транспортер Рецептор Нейротрансмиттер Аксонный терминал Синаптическая щель Дендрит |
Большинство аксонов несут сигналы в виде потенциалов действия, которые представляют собой дискретные электрохимические импульсы, которые быстро проходят по аксону, начиная с тела клетки и заканчиваясь в точках, где аксон производит синаптический контакт с клетками-мишенями. Определяющей характеристикой потенциала действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Когда потенциал действия достигает пресинаптического терминала, он активирует процесс синаптической передачи. Первый шаг — это быстрое открытие каналов для ионов кальция в мембране аксона, позволяя ионам кальция проходить внутрь через мембрану. Результирующее увеличение внутриклеточной концентрации кальция приводит к тому, что синаптические везикулы (крошечные контейнеры, окруженные липидной мембраной), заполненные химическим веществом нейромедиатор, сливаются с мембраной аксона и выводят их содержимое во внеклеточное пространство.. Нейромедиатор высвобождается из пресинаптического нерва посредством экзоцитоза. Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или изменении ее метаболизма каким-либо образом. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор везикул перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона.
 (A) пирамидная клетка, интернейрон и форма волны короткой продолжительности (Axon), наложение трех средних форм волны;. ( B) Средняя и стандартная ошибка времени прохождения пика для интернейронов пирамидных клеток и предполагаемых аксонов;. (C) График разброса отношения сигнал / шум для отдельных единиц в зависимости от времени прохождения пика для аксонов, пирамидных клеток (PYR) и интернейронов (INT).
(A) пирамидная клетка, интернейрон и форма волны короткой продолжительности (Axon), наложение трех средних форм волны;. ( B) Средняя и стандартная ошибка времени прохождения пика для интернейронов пирамидных клеток и предполагаемых аксонов;. (C) График разброса отношения сигнал / шум для отдельных единиц в зависимости от времени прохождения пика для аксонов, пирамидных клеток (PYR) и интернейронов (INT).
Внеклеточные записи распространения потенциала действия в аксонах были продемонстрированы у свободно движущихся животных. В то время как внеклеточные соматические потенциалы действия использовались для изучения клеточной активности у свободно движущихся животных, таких как клетки места, аксональная активность как в белом, так и в сером веществе также может быть записано. Внеклеточные записи распространения потенциала действия аксонов отличаются от соматических потенциалов действия по трем причинам: 1. Сигнал имеет более короткую длительность спада пика (~ 150 мкс), чем у пирамидных клеток (~ 500 мкс) или интернейронов (~ 250 мкс). 2. Изменение напряжения трехфазное. 3. Активность, записанная на тетроде, видна только на одном из четырех проводов записи. В записях от свободно движущихся крыс аксональные сигналы были изолированы в трактах белого вещества, включая альвеус и мозолистое тело, а также серое вещество гиппокампа.
Фактически, генерация потенциалов действия в vivo является последовательным по своей природе, и эти последовательные всплески составляют цифровые коды в нейронах. Хотя предыдущие исследования указывают на аксональное происхождение одного спайка, вызванного кратковременными импульсами, физиологические сигналы in vivo запускают инициирование последовательных спайков в телах клеток нейронов.
Помимо распространения потенциалов действия на аксоны. терминалов, аксон способен усиливать потенциалы действия, что обеспечивает безопасное распространение последовательных потенциалов действия к окончанию аксона. Что касается молекулярных механизмов, потенциал-управляемые натриевые каналы в аксонах обладают более низким порогом и более коротким рефрактерным периодом в ответ на кратковременные импульсы.
Развитие и рост
Развитие
Развитие аксона до его мишени — одна из шести основных стадий в общем развитии нервной системы. Исследования, проведенные на культивируемых нейронах гиппокампа, предполагают, что нейроны изначально продуцируют несколько нейритов, которые эквивалентны, но только одному из этих нейритов суждено стать аксоном. Неясно, предшествует ли спецификация аксона удлинению аксона или наоборот, хотя недавние данные указывают на последнее. Если разрезать не полностью развитый аксон, полярность может измениться, и другие нейриты потенциально могут стать аксоном. Это изменение полярности происходит только в том случае, если аксон сокращен как минимум на 10 мкм короче, чем другие нейриты. Послетого, как разрез будет сделан, самый длинный нейрит станет будущим аксоном, все остальные нейриты, включая исходный аксон, превратятся в дендриты. Наложение внешней силы на нейрит, заставляющее его удлиняться, превращает его в аксон. Тем не менее, развитие аксонов достигается за счет сложного взаимодействия между внеклеточной передачей сигналов, внутриклеточной передачей сигналов и цитоскелетной динамикой.
Внеклеточная передача сигналов
Внеклеточные сигналы, которые распространяются через внеклеточный матрикс, окружающие нейроны, играют важную роль в развитии аксонов. Эти сигнальные молекулы включают белки, нейротрофические факторы, внеклеточный матрикс и молекулы адгезии. Нетрин (также известный как UNC-6), секретируемый белок, участвует в образовании аксонов. Когда рецептор нетрина UNC-5 мутируется, несколько нейритов нерегулярно проецируются из нейронов, и, наконец, один аксон вытягивается вперед. Нейротрофические факторы — фактор роста нервов (NGF), нейротрофический фактор мозга (BDNF) и нейротрофин-3 (NTF3) также участвуют в развитии аксонов. и связываются с рецепторами Трк.
. ганглиозид -превращающий фермент ганглиозид плазматической мембраны алидаза (PMGS), который участвует в активации TrkA на кончике нейтритов, требуется для удлинения аксонов. PMGS асимметрично распределяется по кончику нейрита, которому суждено стать будущим аксоном.
Внутриклеточная передача сигналов
Во время развития аксонов активности PI3K увеличивается в кончик предназначенного аксона. Нарушение активности PI3K тормозит развитие аксонов. Активация PI3K приводит к производству фосфатидилинозитол (3,4,5) -трисфосфата (PtdIns), который может вызвать значительное удлинение нейрита, превращая его в аксон. Таким образом, сверхэкспрессия фосфатаз, вызывающих нарушение поляризации, приводит к нарушению поляризации.
Цитоскелетная динамика
Нейрит с самым низким актиновым филаментом контент станет аксоном. Концентрация PGMS и содержание f-актина обратно коррелированы; когда PGMS становится обогащенным на кончике нейрита, содержание в нем f-актина снижается. Кроме того, воздействие лекарств, деполимеризующих актин, и токсина B (который инактивирует сигналы передачи Rho ) вызывает образование множественных аксонов. Следовательно, разрыв актиновой сети в конусе роста будет производить ее нейрита в аксон.
Рост
 Аксонятидневной мыши с видимым конусом роста
Аксонятидневной мыши с видимым конусом роста
Растущие аксоны проходят через их окружение через конус роста , который находится на кончике аксона. Конус роста имеет широкое пластинчатое расширение, называемое ламеллиподиумом, содержит выступы, называемые филоподиями. Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует нашу среду. Актин играет важную роль в подвижности этой системы. Среда с высоким уровнем молекулы клеточной адгезии (CAM) идеальная среда для роста аксонов. Похоже, это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нейронных систем, включают N-CAM, TAG-1 — аксональный гликопротеин — и MAG, все из которых являются частью суперсемейства иммуноглобулинов. Другой набор молекул, называемый внеклеточный матрикс — молекулы адгезии, также обеспечивает липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин, фибронектин, тенасцин и перлекан. Некоторые из них поверхностно связаны с клетками и, таким образом, как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь длительного длительного действия.
Клетки, называемые направляющими клетками, дают в первую ростом аксонов нейронов. Эти клетки, которые обеспечивают наведению аксонов, обычно представляют собой другие нейроны, иногда являющиеся незрелыми. Когда аксон завершит свой рост в месте его соединения с мишенью, диаметр аксона может увеличиться до раз, в зависимости от требуемой скорости проводимости.
Он также имеет исследованиям было обнаружено, что если аксоны нейрона были повреждены, пока сома (тело клетки нейрона ) не повреждена, аксоны будут регенерировать и воссоздавать синаптические связи с нейронами с помощью ячеек указателя. Это также называется нейрорегенерацией.
Nogo-A — это тип компонента, ингибирующего отрастание нейритов, который присутствует в миелиновых мембранах центральной нервной системы (обнаружен в аксоне). Он играет решающую роль в ограничении регенерации аксонов центральной нервной системы взрослых млекопитающих. В недавних исследованиях, если Nogo-A заблокирован и нейтрализован, можно вызвать регенерацию аксонов на большом расстоянии, что приводит к усилению функционального восстановления у крыс и спинного мозга мыши. Этого еще предстоит сделать на людях. Недавнее исследование также показало, что макрофаги, активируемые специфическим воспалительным процессом, активируемым рецептором дектина-1, способным восстановлением аксонов, однако также вызывают нейротоксичность в нейроне.
Регулировка длины
Аксоны в степени различаются по длине от нескольких микрометров до метров у некоторых животных. Это подчеркивает, что аксонует механизм регулирования длины клетки, позволяющий нейронам ощущать длину своихонов и соответственно должен контролировать их рост. Было обнаружено, что играет важную роль в регулировании длины аксонов. Основываясь на этом наблюдении, исследователи разработали модель роста аксонов, описывающую, как моторные белки, увеличивающие длину аксона на молекулярном уровне. Эти исследования предполагают, что моторные белки переносят сигнальные молекулы от сомы к конусу роста и наоборот, колеблется во времени с параметрами, зависящей от длины.
Классификация
Аксоны нейронов задней части периферической системы можно классифицировать на основе их физических характеристик и свойств проводимости сигнала. Известно, что эти аксоны имеют разную толщину (от 0,1 до 20 мкм), считалось, что эти аксоны связаны со скоростью, определяющей потенциал действия может перемещаться по аксону — скорость его проводимости. Эрлангер и Гассер подтвердили эти гипотезу и идентифицировали несколько типов волокон, установив связь между аксона и его скоростью проводимости нерва. Они опубликовали свои открытия в 1941 году, дав первую класси установкуонов.
Аксоны подразделяются на две системы. Первый, введенный Эрлангером и Гассером, сгруппировал волокна три основных, используя буквы A, B и C. Эти группы: группа A, группа B и группа C включает как сенсорные волокна (афференты ), так и моторные волокна (эфференты ). Первая группа A была разделена на альфа, бета, гамма и дельта волокна — Aα, Aβ, Aγ и Aδ. Моторными нейронами различных моторных волокон были нижние мотонейроны — альфа-мотонейрон, бета-мотонейрон и гамма-мотонейрон <55.>Имеющий нервные волокна Aα, Aβ и Aγ соответственно.
Позже другие исследователи обнаружили две группы волокон Aa, которые были сенсорными волокнами. Затем они были введены в систему, включающую только сенсорные волокна (хотя некоторые из них были смешанными нервами, а также двигательными волокнами). Эта система называет сенсорные группы Типами и использует римские цифры: Тип Ia, Тип Ib, Тип II, Тип III и Тип IV.
Мотор
Нижние мотонейроны имеют два типа волокон:
| Тип | Эрлангер-Гассер. Классификация | Диаметр. (мкм) | Миелин | Проводимость. скорость (м / с) | Связанные мышечные волокна |
|---|---|---|---|---|---|
| α | Aα | 13-20 | Да | 80–120 | Экстрафузальные мышечные волокна |
| β | Aβ | ||||
| γ | Aγ | 5-8 | Да | 4–24 | Внутрифузальные мышечные волокна |
Сенсорные
Различные сенсорные рецепторы иннервируют разные типы нервных волокон. Проприоцепторы иннервируются сенсорными волокнами типа Ia, Ib и II, механорецепторами сенсорными волокнами типа II и III и ноцицепторами и терморецепторами сенсорными волокнами III и IV типа.
| Тип | Эрлангер-Газсер. Классификация | Диаметр. (мкм) | Миелин | Проводимость. скорость (м / с) | Ассоциированные сенсорные рецепторы | Проприорецепторы | Механоцепторы | Ноцицепторы и. терморецепторы |
|---|---|---|---|---|---|---|---|---|
| Ia | Aα | 13-20 | Да | 80–120 | Первичные рецепторы мышечного веретена (кольцевидное окончание) | ✔ | ||
| Иб | Aα | 13-20 | Да | 80–120 | Сухожильный орган Гольджи | |||
| II | Aβ | 6-12 | Да | 33–75 | Вторичные рецепторы мышечное веретено (окончание цветочно-спрей).. Все кожные механорецепторы | ✔ | ||
| III | Aδ | 1-5 | Тонкий | 3–30 | Свободные нервные окончания прикосновения и давление. Ноцицепторы бокового спиноталамического тракта. Холодные терморецепторы | ✔ | ||
| IV | C | 0,2- 1,5 | No | 0,5-2,0 | Ноцицепторы переднего спинного мозга. Тепловые рецепторы |
вегетативные
вегетативная нервная система имеет два вида периферического волокна s:
| Тип | Erlanger-Gasser. Классификация | Диаметр. (мкм) | Myelin | Проводимость. скорость (м / с) |
|---|---|---|---|---|
| преганглионарные волокна | B | 1–5 | Да | 3–15 |
| постганглионарные волокна | C | 0,2–1,5 | No | 0,5–2, 0 |
Клиническая значимость
По степени тяжести повреждение нерва может быть описана как нейропраксия, аксонотмезис или невротмезис. Сотрясение мозга считается легкой формой диффузного повреждения аксонов. Поражение аксонов также может вызывать центральный хроматолиз. Дисфункция аксонов в нервной системе является одной из причин многих наследственных неврологических расстройств, которые поражают периферические, так и центральные нейроны.
Когда аксон разрушается, активный процесс дегенерация аксона происходит в части аксона, наиболее удаленной от тела клетки. Эта дегенерация происходит быстро после травмы, когда часть аксона блокируется мембранами и разрушается макрофагами. Это известно как валлеровское вырождение. Отмирание аксона также может иметь место при многих нейродегенеративных заболеваниях, особенно при нарушении транспорта аксонов, известно как дегенерация, подобная валлеровской. Исследования показывают, что аксональный белок NMNAT2 не может достичь всего аксона.
Демиелинизация аксонов вызывает множество неврологических симптомов, обнаруженных при заболевании рассеянный склероз.
Дисмиелинизация — это аномальное образование миелиновой оболочки. Это связано с использованием лейкодистрофиями, а также с шизофренией.
Тяжелая черепно-мозговая травма может привести к обширным пораженным нервным путям, повреждая аксоны в состоянии, известном как диффузное повреждение аксонов. Это может привести к устойчивому вегетативному состоянию. В исследованиих на крысе было показано, что повреждение аксонов в результате повреждения однократной черепно-мозговой травмы может привести к дальнейшему повреждению после повторных легких черепно-мозговых травм.
A канал для направления нервов является искусственным средством управления ростом аксонов для обеспечения нейрорегенерации и одним из многих методов лечения, используется для травм нерва.
История
Немецкий анатом Отто Фридриху Карлу Дейтерсу обычно приписывают открытие аксона, отличив его от дендритов. Швейцарский Рюдольф Альберт фон Келликер и немец Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Кёлликер назвал аксон в 1896 году. Луи-Антуан Ранвье был первым, кто описал промежутки или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлами Ранвье. Сантьяго Рамон-и-Кахал, испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции. Джозеф Эрлангер и Герберт Гассер ранее разработали классификацию система для периферических нервных волокон, основанная на скорости аксональной проводимости, миелинизации, размере волокон и т. д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), а к 1952 году они получили полное количественное описание ионной основы потенциала действия, что привело к формулировке модели Ходжкина-Хаксли. Ходжкин и Хаксли были совместно удостоены Нобелевской премии за эту работу в 1963 году. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхейзера-Хаксли. Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает множество деталей об отдельных ионных каналах.
других животных
Аксоны у беспозвоночных были тщательно изучены. прибрежный кальмар, часто используемый в качестве модельного организма, имеет самый длинный из известных аксонов. гигантский кальмар имеет самый большой известный аксон. Его размер составляет от половины (обычно) до одного миллиметра в диаметре, и он используется для управления его системой реактивного движения. Самая быстрая зарегистрированная скорость проводимости 210 м / с обнаружена в заключенных в оболочку аксонах некоторых пелагических креветок Penaeid, а обычный диапазон составляет от 90 до 200 м / с (cf 100– 120 м / с для самого быстрого аксона миелинизированных позвоночных.)
В других случаях, как видно из исследований на крысах, аксон происходит из дендрита; такие аксоны, как говорят, имеют «дендритное происхождение». Некоторые аксоны с дендритным происхождением аналогичным образом имеют «проксимальный» начальный сегмент, который начинается непосредственно в источнике аксона, в то время как другие имеют «дистальный» начальный сегмент, заметно отделенный от источника аксона. У многих видов некоторые из нейронов имеют аксоны, которые исходят из дендрита, а не из тела клетки, и они известны как дендриты, несущие аксоны. Во многих случаях аксон берет свое начало от бугорка аксона на соме; такие аксоны, как говорят, имеют «соматическое происхождение». Некоторые аксоны соматического происхождения имеют «проксимальный» начальный сегмент, прилегающий к бугорку аксона, в то время как другие имеют «дистальный» начальный сегмент, отделенный от сомы расширенным бугорком аксона.
См. Также
- Электрофизиология
- Ганглиозное возвышение
- Гигантская аксональная нейропатия
- Нейронное отслеживание
- Пионерный аксон
Ссылки
Внешние ссылки
- Изображение гистологии: 3_09 в Центре медицинских наук Университета Оклахомы — «Слайд 3 Спинной мозг »
Аксон – это волокнистая ось, отходящая от тела нейрона, покрытая миелиновым слоем, обеспечивающая связь с другими нейронами и клетками рабочих органов. Представляет собой удлиненный осевой отросток, по которому передаются потенциалы действия (возбуждения), что делает его важнейшим структурным элементом ЦНС.
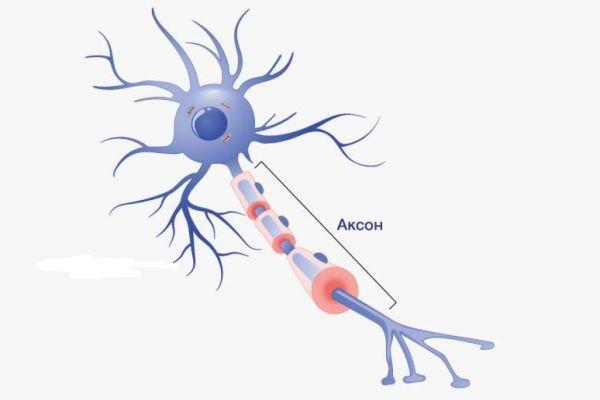
Определение
Мозговое вещество – высокоорганизованная структура, образованная нервными клетками, от которых отходят аксоны. Из нервных клеток состоит мозговая ткань. Аксон в переводе с греческого означает «ось» – это такой отросток, элемент мозгового вещества, который обеспечивает взаимодействие между клетками разного типа (нейроны, клетки иннервируемых органов), что ассоциируется с тонким, четким управлением работой органов и систем. Функции ткани ЦНС:
- Воспринимает раздражения, преобразуя их в импульсы.
- Поддерживает передачу импульсов от управляющих отделов мозга к исполнительным органам.
- Формирует ответную реакцию на раздражающее воздействие.
- Обеспечивает взаимодействие в работе систем и органов, поддерживает интеграцию структурных единиц организма.
- Обеспечивает взаимосвязь организма с внешней средой.
Согласно определению в биологии, аксон (англ. axon) – удлиненный отросток, по которому идут импульсы от тела нейрона к другим нервным клеткам и структурным элементам всех тканей организма. Мозговая ткань в период внутриутробного развития образуется из нервной пластины. Края пластинки прогибаются, что приводит к формированию валиков и желобка. В результате смыкания краев валиков возникает нервная трубка – основа ЦНС.
Дифференциация клеток, образующих трубку, приводит к появлению нейробластов и спонгиобластов. Первые служат основой для формирования нейронов, вторые – для образования нейроглии. Нейроны (анат.) – основные структурные элементы мозгового вещества. Они характеризуются отсутствием функции деления, что приводит к постепенному уменьшению их численности. Тело нейрона состоит из ядра и цитоплазмы. В зависимости от типа нейронов меняется геометрическая форма тела, которая бывает круглая, овальная, пирамидальная и другая.
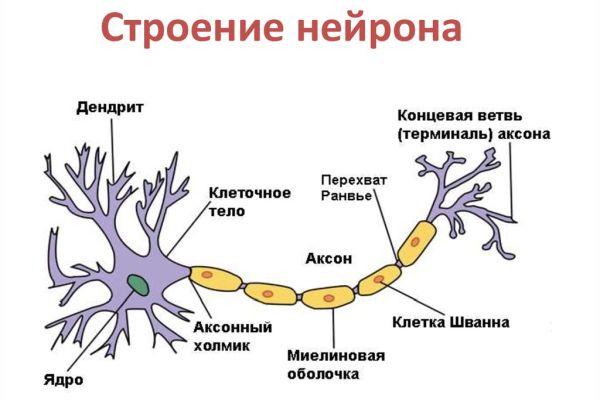
Цитоскелет, состоящий из микротрубочек и нейрофибриллов, обеспечивает опорную и трофическую функцию. Цитоскелет поддерживает форму нейрона, обеспечивает транспорт веществ и органелл. От тела ответвляются отростки – единичный аксон и множественные дендриты. Аксон нейрона почти не ветвится, иногда образует коллатеральные (обходные) сегменты. Концевые сегменты (окончания) разветвляются, называются терминали.
Терминали взаимосвязаны с окончаниями других нейронов и с клетками, образующими паренхиму (ткань) рабочих органов – мышц, желез. Количество дендритов варьируется от 1 до нескольких. Тонкие ответвления дендритов оканчиваются небольшими шипами, где сосредоточены терминали аксональных отростков многих тысяч других клеток. Дендриты воспринимают раздражения или потенциалы действия от других клеток и передают их по волокнам к телу своего нейрона.
Рост аксона зависит от особенностей строения и жизнедеятельности нейрона, который поддерживает функцию питания отростка. К примеру, если перерезать аксональный ствол, сегмент, связанный с телом, остается жизнеспособным и продолжает деятельность, участок, утративший связь с телом, отмирает. Аксоны образуют нервы, что предполагает сложную структурно-морфологическую организацию ЦНС.
Строение
Аксон – это длинный отросток нейрона, который обеспечивает взаимодействие между нервными клетками. Согласно анатомии, аксон ответвляется от холмика, находящегося на теле. Холмик аксона представляет собой структуру, где постсинаптический потенциал преобразуется в биоэлектрический сигнал. Чтобы в холмике происходила генерация биоэлектрических сигналов, необходима согласованная деятельность каналов –натриевых, кальциевых, нескольких типов калиевых.
Длина аксона у человека существенно варьируется в зависимости от вида нейрона, от которого отходит аксональная ось. Минимальная длина – около 1 миллиметра, максимальная – около 1,5 метров. Длина более 1 метра наблюдается в случаях, когда отросток отходит от спинного мозга в область конечностей. Диаметр аксональной оси также неодинаковый у разных типов клеток, равен около 1-20 микрон. Импульсы проходят быстрее по аксональным осям большего диаметра.
Размеры аксонального отростка нередко достигают 99% от общего объема нервной клетки, в структуру которой он входит. Аксон состоит из протоплазмы (аксоплазмы), где находятся тончайшие волокна, белковые нити – нейрофибриллы, из чего образован ствол аксонального ответвления. Согласно одной из теорий, нейрофибриллы – проводники питательных веществ. Аксональная протоплазма также содержит митохондрии и микротрубочки, которые представляют собой самые крупные элементы цитоскелета.
Диаметр микротрубочек составляет около 24 нанометров. Они обеспечивают внутриклеточный транспорт веществ, в том числе поддерживают трофику аксональных отростков. Тело (перикарион) – источник протеинов и нейромедиаторов, распространяющихся по аксональной оси посредством микротрубочек, которые у аксона имеют направленную полярную ориентацию (в отличие от микротрубочек дендритов).
Положительно заряженные концы микротрубочек направлены к сегменту терминали, отрицательно заряженные концы – к телу. Строение аксона предполагает наличие оболочки. Аксон покрыт глиальным (миелиновым) слоем по всей длине, чем защищен от разрушающих внешних воздействий. Миелиновый слой в аксональных отростках периферического отдела сформирован клетками Шванна.
Миелиновая оболочка, покрывающая нервную ось, обеспечивает ее механическую прочность, электрохимическую изоляцию, трофику (питание). Миелиновый слой ускоряет проведение биоэлектрических сигналов. Нервы – пучки объединенных аксональных отростков, которые покрыты оболочкой из соединительной ткани и снабжены кровеносными сосудами.
Функции
Основная задача нейронов – переработка данных. С их помощью осуществляется получение, обработка, передача информации отделам нервной и других систем организма.
Если дендриты проводят сигналы по направлению к телу нервной клетки (перикариону), то аксональный отросток передает импульсы от перикариона к другим клеткам.
Основная функция аксонов – проведение импульсов в пределах нейрональной сети и к исполнительным органам. Аксональные ответвления относятся к первичным проводниковым путям в нервной системе. Вспомогательная функция – транспорт веществ. При помощи аксонального транспорта осуществляется движение белков, синтезированных в теле, нейромедиаторов, органелл. Многие вещества способны двигаться в обоих направлениях.

В периферических сегментах аксона в него могут проникать вирусы и токсичные вещества, которые перемещаясь к телу нервной клетки, повреждают ее. Аксональный транспорт зависит от количества энергии АТФ. Если энергетический уровень АТФ понижается больше, чем в 2 раза, происходит блокировка аксонального транспорта.
Функции аксона заключаются в передаче импульсов. При взаимодействии аксона с телом другого нейрона образуется аксосоматический контакт. Если аксон взаимодействует с дендритами других клеток возникает аксодендритический контакт. Взаимодействие с аксоном другой клетки приводит к образованию аксо-аксонального контакта, который редко происходит в нервной системе, поддерживает тормозные рефлекторные реакции.
Особенности регенерации нервной ткани
Нервные клетки почти полностью лишены способности к регенерации. Однако нервные клетки способны восстанавливать поврежденные или утраченные ответвления. Процесс регенерации аксона возможен, если тело сохраняет жизнеспособность, и на пути роста аксонального отростка отсутствуют препятствия. В ходе процесса регенерации отросток вновь прорастает к органу-мишени.
Восстановление нервной проводимости в мышцах с нарушенной иннервацией – один из критериев успешного лечения невропатий разного генеза. При невропатиях травматического генеза восстановление функций мышц происходит за счет регенерации ствола прерванного аксона и ремиелинизации отростка (восстановление миелиновой оболочки). Периферический отдел нервной системы обладает более высоким потенциалом регенерационных возможностей в сравнении с центральным отделом.
Восстановление иннервации в мышечной или кожной ткани происходит благодаря сохранившимся аксонам, которые начинают ускоренно разрастаться и ветвиться. Процесс ветвления аксонов в зоне перехватов Ранвье (периодические разрывы миелинового слоя) получил название «спрутинг». В результате происходит частичное или полное возобновление первичной иннервации.
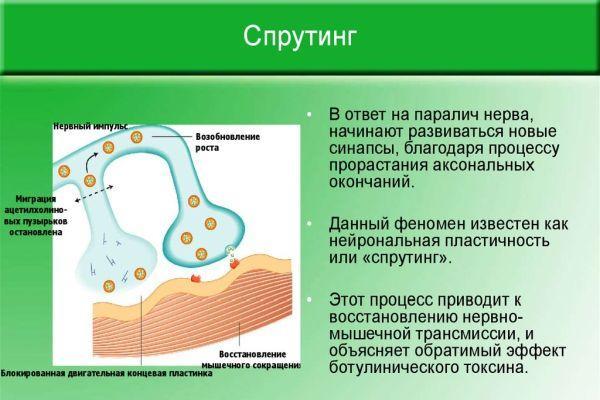
В ходе экспериментов установлено, что близлежащие интактные (не вовлеченные в патологический процесс) аксоны выпускают нервные волокна, которые иннервируют участок мускулатуры или кожных покровов с нарушенной проводимостью нервных импульсов. Различают виды спрутинга – коллатеральный (обходной) и регенераторный (терминальный).
Регенераторный спрутинг начинается после устранения в нейронах ретроградных изменений, обусловленных аксонотомией (повреждением, рассечением нервной оси). Это связано с потребностью в продукции аппарата ядра нервной клетки, производящего протеины. Материал, необходимый для регенераторного спрутинга, продуцируется в теле и транспортируется по микротрубочкам по всей длине оси. Параллельно происходит процесс ремиелинизации осевого ствола.
Аксон – удлиненный отросток нервной клетки, обеспечивающий взаимодействие между структурными элементами мозговой ткани и связь ЦНС с исполнительными органами.
Просмотров: 2 336
Установите соответствие между строением и функциями нейрона и его отростками.
СТРОЕНИЕ И ФУНКЦИИ
А) проводит сигнал к телу нейрона
Б) снаружи покрыт миелиновой оболочкой
В) короткий и сильно ветвится
Г) участвует в образовании нервных волокон
Д) проводит сигнал от тела нейрона
Запишите в ответ цифры, расположив их в порядке, соответствующем буквам:
| A | Б | В | Г | Д |
Спрятать пояснение
Пояснение.
Дендрит — это короткий (до 700 мкм; 700 мкм = 0,7 мм) ветвящийся цитоплазматический отросток нейрона, проводящий нервные импульсы к телу нейрона. Остальные признаки принадлежат аксону.
Ответ: 21211.
Примечание.
Дендрит — дихотомически ветвящийся отросток нервной клетки (нейрона), воспринимающий сигналы от других нейронов, рецепторных клеток или непосредственно от внешних раздражителей. Проводит нервные импульсы к телу нейрона. В поверхностной структуре нейроцитов могут насчитываться от нуля до множества дендритов. Аксон чаще всего единственный. Поверхность дендритов не имеет миелиновой оболочки в отличие от аксонных отростков.
Дендритные окончания имеют разные размеры, но всегда характеризуются постепенным уменьшением диаметра претерминальных веточек. Длина обычно от нескольких мкм до 1 мм. Но, например, у некоторых чувствительных нейронов спинномозговых ганглиев дендриты очень длинные – до метра и более.
| Аксон | |
|---|---|
|
Аксон многополярного нейрона |
|
| Идентификаторы | |
| MeSH | D001369 |
| Анатомическая терминология
[редактировать в Викиданных ] |
An аксон (от греч. ἄξων áxōn, ось), или нервное волокно (или же нерв волокно: видеть орфографические различия ), представляет собой длинный тонкий выступ нервной клетки, или нейрон, у позвоночных, который обычно проводит электрические импульсы, известные как потенциалы действия подальше от тело нервной клетки. Функция аксона — передавать информацию различным нейронам, мышцам и железам. В определенных сенсорные нейроны (псевдоуниполярные нейроны ), например аксоны прикосновения и тепла, аксоны называются афферентные нервные волокна и электрический импульс проходит по ним от периферия к телу клетки и от тела клетки к спинному мозгу по другой ветви того же аксона. Дисфункция аксонов является причиной многих наследственных и приобретенных неврологических расстройств, которые могут поражать как периферические, так и центральные нейроны. Нервные волокна классифицированный на три типа — нервные волокна группы А, нервные волокна группы B, и нервные волокна группы C. Группы A и B являются миелинизированный, и группа C немиелинизированы. Эти группы включают как сенсорные волокна, так и двигательные волокна. Другая классификация группирует только сенсорные волокна как Тип I, Тип II, Тип III и Тип IV.
Аксон — это один из двух типов цитоплазматический выступы из тела клетки нейрона; другой тип — это дендрит. Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно имеют постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты получают сигналы, тогда как аксоны передают их). Некоторые типы нейронов не имеют аксона и передают сигналы от своих дендритов. У некоторых видов аксоны могут исходить из дендритов, известных как дендриты, несущие аксоны.[1] Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга.[2]
Аксоны покрыты мембраной, известной как аксолемма; цитоплазма аксона называется аксоплазма. Большинство аксонов разветвляются, в некоторых случаях очень обильно. Концевые ветви аксона называются телодендрия. Распухший конец телодендрона известен как терминал аксона который присоединяется к дендрону или телу клетки другого нейрона, образуя синаптическое соединение. Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсы. В некоторых случаях аксон одного нейрона может образовывать синапс с дендритами того же нейрона, что приводит к autapse. В синапсе мембрана аксона близко примыкает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Некоторые синаптические соединения появляются вдоль аксона по мере его расширения — они называются мимоходом («попутно») синапсов и может быть сотнями или даже тысячами вдоль одного аксона.[3] Другие синапсы выглядят как терминалы на концах аксональных ветвей.
Отдельный аксон со всеми его ветвями, взятыми вместе, может иннервировать несколько частей мозга и генерируют тысячи синаптических окончаний. Связка аксонов делает нервный тракт в Центральная нервная система,[4] и пучок в периферическая нервная система. В плацентарные млекопитающие самый большой белое вещество тракт в головном мозге мозолистое тело, образованный примерно из 200 миллионов аксонов в человеческий мозг.[4]
Анатомия
Типичный миелинизированный аксон
Аксоны — это основные линии передачи нервная система, и как пучки они образуют нервы. Некоторые аксоны могут достигать одного метра и более, а другие — всего лишь один миллиметр. Самые длинные аксоны в человеческом теле — аксоны седалищный нерв, которые идут от основания спинной мозг к большому пальцу каждой стопы. Диаметр аксонов также варьируется. Большинство отдельных аксонов микроскопические в диаметре (обычно около одного микрометр (мкм) в поперечнике). Самые большие аксоны млекопитающих могут достигать в диаметре до 20 мкм. В гигантский аксон кальмара, который специализируется на очень быстром проведении сигналов, близок к 1 миллиметр в диаметре, размером с грифель карандаша. Количество аксональных телодендрий (разветвленных структур на конце аксона) также может отличаться от одного нервного волокна к другому. Аксоны в Центральная нервная система (ЦНС) обычно показывают несколько телодендрий с множеством синаптических конечных точек. Для сравнения: гранулярная клетка мозжечка аксон характеризуется одним Т-образным узлом ветви, от которого два параллельные волокна продлевать. Продуманное ветвление позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
Есть два типа аксонов в нервная система: миелинизированный и немиелинизированный аксоны.[5] Миелин представляет собой слой изолирующего жирового вещества, который образован двумя типами глиальные клетки Клетки Шванна и олигодендроциты. в периферическая нервная система Шванновские клетки образуют миелиновую оболочку миелинизированного аксона. в Центральная нервная система олигодендроциты образуют изолирующий миелин. Вдоль миелинизированных нервных волокон пробелы в миелиновой оболочке, известные как узлы Ранвье происходят через равные промежутки времени. Миелинизация обеспечивает особенно быстрый режим распространения электрических импульсов, называемый скачкообразное проведение.
Миелинизированные аксоны из корковые нейроны образуют основную часть нервной ткани, называемой белое вещество в мозгу. Миелин придает белому цвету ткань в отличие от серое вещество коры головного мозга, содержащей тела нейрональных клеток. Аналогичное расположение можно увидеть на мозжечок. Связки миелинизированных аксонов составляют нервные пути в ЦНС. Там, где эти тракты пересекают среднюю линию мозга и соединяют противоположные области, они называются комиссур. Самый крупный из них — мозолистое тело что соединяет два полушария головного мозга, а это около 20 миллионов аксонов.[4]
Видно, что структура нейрона состоит из двух отдельных функциональных областей или компартментов — тела клетки вместе с дендритами в качестве одной области и аксональной области в качестве другой.
Аксональная область
Аксональная область или компартмент включает бугорок аксона, начальный сегмент, остальную часть аксона и телодендрии аксона, а также терминалы аксона. Он также включает миелиновую оболочку. В Тела Nissl которые продуцируют нейрональные белки, отсутствуют в аксональной области.[3] Белки, необходимые для роста аксона и удаления отходов жизнедеятельности, нуждаются в транспортном каркасе. Этот аксональный транспорт обеспечивается в аксоплазме за счет расположения микротрубочки и промежуточные нити известный как нейрофиламенты.
Аксонный бугорок
Деталь, показывающая микротрубочки на бугорке аксона и начальном сегменте.
В аксональный бугорок это область, образованная клеточным телом нейрона по мере того, как она расширяется, чтобы стать аксоном. Он предшествует начальному сегменту. Полученные потенциалы действия, которые подведены в нейроне передаются на бугорок аксона для генерации потенциала действия из начального сегмента.
Начальный сегмент
В аксональный начальный сегмент (AIS) представляет собой структурно и функционально отдельный микродомен аксона.[6][7] Одна из функций начального сегмента — отделить основную часть аксона от остальной части нейрона; другая функция — помочь инициировать потенциалы действия.[8] Обе эти функции поддерживают нейрон полярность ячейки, в котором дендриты (а в некоторых случаях сома ) нейрона получают входные сигналы в базальной области, а в апикальной области аксон нейрона обеспечивает выходные сигналы.[9]
Начальный сегмент аксона немиелинизирован и содержит специализированный комплекс белков. Его длина составляет примерно от 20 до 60 мкм, и он функционирует как место инициации потенциала действия.[10][11] Как положение на аксоне, так и длина AIS могут изменяться, показывая степень пластичности, которая может точно настроить вывод нейронов.[10][12] Более длинный AIS связан с большей возбудимостью.[12] Пластичность также проявляется в способности AIS изменять свое распределение и поддерживать активность нейронных схем на постоянном уровне.[13]
AIS узкоспециализирована для быстрого проведения нервные импульсы. Это достигается за счет высокой концентрации напряжение-управляемые натриевые каналы в начальном сегменте, где возникает потенциал действия.[13] Ионные каналы сопровождаются большим количеством молекулы клеточной адгезии и каркасные белки, которые прикрепляют их к цитоскелету.[10] Взаимодействие с Анкирин Г важны, так как это главный организатор в САИ.[10]
Аксональный транспорт
В аксоплазма эквивалентен цитоплазма в клетка. Микротрубочки образуются в аксоплазме на бугорке аксона. Они расположены по длине аксона в перекрывающихся участках и все направлены в одном направлении — к окончанию аксона.[14] Об этом говорят положительные окончания микротрубочек. Такое перекрывающееся расположение обеспечивает маршруты транспортировки различных материалов из тела клетки.[14] Исследования аксоплазмы показали движение многочисленных пузырьков всех размеров, которые можно увидеть вдоль цитоскелетных филаментов — микротрубочек и нейрофиламенты, в обоих направлениях между аксоном и его окончаниями и телом клетки.
Исходящий антероградный транспорт из тела клетки по аксону переносит митохондрии и мембранные белки, необходимые для роста, к концу аксона. Входящий ретроградный транспорт переносит отходы клетки от терминала аксона к телу клетки.[15] Исходящие и входящие треки используют разные наборы моторные белки.[14] Исходящий транспорт обеспечивает кинезин, а входящий обратный трафик обеспечивается динеин. Динеин направлен на минус-конец.[15] Существует множество форм моторных белков кинезина и динеина, и считается, что каждая из них несет свой груз.[14] Исследования транспорта в аксоне привели к названию кинезина.[14]
Миелинизация
ТЕМ миелинизированного аксона в поперечном сечении.
В нервной системе аксоны могут быть миелинизированный, или немиелинизированные. Это обеспечение изолирующего слоя, называемого миелиновой оболочкой. Миелиновая мембрана уникальна своим относительно высоким соотношением липидов к белку.[16]
В периферической нервной системе аксоны миелинизируются глиальные клетки известный как Клетки Шванна. В центральной нервной системе миелиновую оболочку представляют глиальные клетки другого типа, олигодендроцит. Клетки Шванна миелинизируют единственный аксон. Олигодендроцит может миелинизировать до 50 аксонов.[17]
Состав миелина у этих двух типов разный. В ЦНС основным белком миелина является протеолипидный белок, а в ПНС это основной белок миелина.
Узлы Ранвье
Узлы Ранвье (также известен как щели миелиновой оболочки) — короткие немиелинизированные сегменты миелинизированный аксон, которые находятся периодически вкраплениями между сегментами миелиновой оболочки. Следовательно, в точке узла Ранвье аксон уменьшается в диаметре.[18] Эти узлы являются областями, где могут быть созданы потенциалы действия. В скачкообразное проведение электрические токи, возникающие в каждом узле Ранвье, передаются с небольшим затуханием к следующему узлу в линии, где они остаются достаточно сильными, чтобы генерировать другой потенциал действия. Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Терминалы Axon
Аксон может делиться на множество ветвей, называемых телодендриями (греч. Конец дерева). В конце каждого телодендрон является аксонный терминал (также называемый синаптическим бутоном или терминальным бутоном). Терминалы Axon содержат синаптические везикулы которые хранят нейротрансмиттер для выпуска в синапс. Это делает возможными множественные синаптические связи с другими нейронами. Иногда аксон нейрона может синапсировать с дендритами того же нейрона, когда он известен как autapse.
Потенциалы действия
| Структура типового химический синапс |
|---|
|
Постсинаптический Напряжение- Синаптический Нейротрансмиттер Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит |
Большинство аксонов несут сигналы в виде потенциалы действия, которые представляют собой дискретные электрохимические импульсы, которые быстро проходят по аксону, начиная с тела клетки и заканчивая в точках, где аксон производит синаптический контакт с клетками-мишенями. Определяющей характеристикой потенциала действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Когда потенциал действия достигает пресинаптического терминала, он активирует процесс синаптической передачи. Первый шаг — это быстрое открытие каналов для ионов кальция в мембране аксона, позволяя ионам кальция проходить внутрь через мембрану. Возникающее в результате повышение концентрации внутриклеточного кальция вызывает синаптические везикулы (крошечные контейнеры, окруженные липидной мембраной), заполненные нейротрансмиттер химикат, чтобы сливаться с мембраной аксона и выводить их содержимое во внеклеточное пространство. Нейромедиатор высвобождается из пресинаптического нерва через экзоцитоз. Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или каким-либо образом изменяющем ее метаболизм. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор везикул перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона.[5]
(A) пирамидная ячейка, интернейрон и форма волны короткой длительности (Axon), наложение трех средних форм волны;
(B) Средняя и стандартная ошибка времени прохождения пика для интернейронов пирамидных клеток и предполагаемых аксонов;
(C) График разброса отношения сигнал / шум для отдельных единиц от времени пика для аксонов, пирамидных клеток (PYR) и интернейронов (INT).
Внеклеточные записи потенциал действия распространение в аксонах было продемонстрировано у свободно перемещающихся животных. В то время как внеклеточные соматические потенциалы действия использовались для изучения клеточной активности у свободно перемещающихся животных, таких как разместить клетки, аксональная активность в обоих белый и серое вещество также могут быть записаны. Внеклеточные записи распространения потенциала действия аксона отличаются от соматических потенциалов действия по трем причинам: 1. Сигнал имеет более короткую продолжительность пика и спада (~ 150 мкс), чем у пирамидных клеток (~ 500 мкс) или интернейронов (~ 250 мкс). 2. Изменение напряжения трехфазное. 3. Активность, записанная на тетроде, видна только на одном из четырех проводов записи. В записях от свободно движущихся крыс аксональные сигналы были изолированы в трактах белого вещества, включая альвеус и мозолистое тело, а также серое вещество гиппокампа.[19]
Фактически, поколение потенциалы действия in vivo носит последовательный характер, и эти последовательные всплески составляют цифровые коды в нейроны. Хотя предыдущие исследования указывают на аксональное происхождение одиночного спайка, вызванного кратковременными импульсами, физиологические сигналы in vivo запускают инициирование последовательных спайков в телах клеток нейронов.[20][21]
В дополнение к распространению потенциалов действия к окончанию аксона, аксон способен усиливать потенциалы действия, что гарантирует безопасное распространение последовательных потенциалов действия к окончанию аксона. Что касается молекулярных механизмов, напряжение-управляемые натриевые каналы в аксонах обладают нижними порог и короче период отражения в ответ на кратковременные импульсы.[22]
Развитие и рост
Разработка
Развитие аксона к его цели — один из шести основных этапов в общей развитие нервной системы.[23] Исследования проведены на культивированных гиппокамп нейроны предполагают, что нейроны изначально производят несколько невриты которые эквивалентны, но только одному из этих нейритов суждено стать аксоном.[24] Неясно, предшествует ли спецификация аксона удлинению аксона или наоборот,[25] хотя недавние свидетельства указывают на последнее. Если разрезать не полностью развитый аксон, полярность может измениться, и другие нейриты потенциально могут стать аксоном. Это изменение полярности происходит только в том случае, если аксон сокращен как минимум на 10 мкм короче, чем другие нейриты. После того, как разрез будет сделан, самый длинный нейрит станет будущим аксоном, а все остальные нейриты, включая исходный аксон, превратятся в дендриты.[26] Воздействие внешней силы на нейрит, заставляющее его удлиниться, превращает его в аксон.[27] Тем не менее, развитие аксонов достигается за счет сложного взаимодействия между внеклеточной передачей сигналов, внутриклеточной передачей сигналов и цитоскелет динамика.
Внеклеточные сигналы, распространяющиеся через внеклеточный матрикс окружающие нейроны играют важную роль в развитии аксонов.[28] Эти сигнальные молекулы включают белки, нейротрофические факторы, внеклеточный матрикс и молекулы адгезии. Нетрин (также известный как UNC-6) секретируемый белок, участвующий в образовании аксонов. Когда UNC-5 рецептор нетрина мутирован, несколько нейритов нерегулярно проецируются из нейронов, и, наконец, один аксон выходит вперед.[29][30][31][32] Нейротрофические факторы — фактор роста нервов (NGF), нейротрофический фактор головного мозга (BDNF) и нейротрофин-3 (NTF3) также участвуют в развитии аксонов и связываются с Рецепторы Trk.[33]
В ганглиозид -превращающий фермент ганглиозид плазматической мембраны сиалидаза (PMGS), который участвует в активации TrkA на кончике нейтритов, требуется для удлинения аксонов. PMGS асимметрично распределяется по кончику нейрита, которому суждено стать будущим аксоном.[34]
Внутриклеточная передача сигналов
Во время развития аксонов активность PI3K увеличивается на конце предназначенного аксона. Нарушение активности PI3K тормозит развитие аксонов. Активация PI3K приводит к образованию фосфатидилинозит (3,4,5) -трисфосфат (PtdIns), которые могут вызывать значительное удлинение нейрита, превращая его в аксон. Таким образом, чрезмерное проявление фосфатазы что дефосфорилирование PtdIns приводит к нарушению поляризации.[28]
Цитоскелетная динамика
Неврит с самым низким актин содержимое филамента станет аксоном. Концентрация PGMS и f-актин содержание обратно коррелировано; когда PGMS становится обогащенным на кончике нейрита, содержание в нем f-актина существенно снижается.[34] Кроме того, воздействие препаратов, деполимеризующих актин, и токсина B (который инактивирует Rho-сигнализация ) вызывает образование множественных аксонов. Следовательно, разрыв актиновой сети в конусе роста будет способствовать превращению ее нейрита в аксон.[35]
Рост
Аксон девятидневной мыши с видимым конусом роста
Растущие аксоны перемещаются в окружающей среде через конус роста, который находится на кончике аксона. Конус роста имеет широкое пластинчатое расширение, называемое ламеллиподиум которые содержат выступы, называемые филоподия. Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует окружающую среду. Актин играет важную роль в мобильности этой системы. Среды с высоким уровнем молекулы клеточной адгезии (CAM) создают идеальную среду для роста аксонов. Похоже, что это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нейронных систем, включают: N-CAM, ТЕГ-1 — аксональный гликопротеин —[36]-и МАГ, все из которых являются частью иммуноглобулин надсемейство. Другой набор молекул называется внеклеточный матрикс -молекулы адгезии также обеспечивают липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин, фибронектин, тенасцин, и перлекан. Некоторые из них поверхностно связаны с клетками и, таким образом, действуют как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь эффекты длительного действия.
Ячейки называются клетки-указатели помочь в руководство роста аксонов нейронов. Эти клетки, которые помогают управление аксоном, как правило, другие нейроны, которые иногда незрелые. Когда аксон завершит свой рост в месте его соединения с мишенью, диаметр аксона может увеличиться до пяти раз, в зависимости от скорость проведения требуется.[37]
В ходе исследований также было обнаружено, что если аксоны нейрона были повреждены, то до тех пор, пока сома (тело клетки нейрон ) не повреждается, аксоны регенерируют и восстанавливают синаптические связи с нейронами с помощью клетки-указатели. Это также называется нейрорегенерация.[38]
Ного-А это тип компонента, ингибирующего рост нейритов, который присутствует в миелиновых мембранах центральной нервной системы (обнаружен в аксоне). Он играет решающую роль в ограничении регенерации аксонов в центральной нервной системе взрослых млекопитающих. В недавних исследованиях, если Nogo-A заблокирован и нейтрализован, можно вызвать регенерацию аксонов на большом расстоянии, что приводит к усилению функционального восстановления у крыс и спинного мозга мыши. Этого еще предстоит сделать на людях.[39] Недавнее исследование также показало, что макрофаги активируется специфическим воспалительным путем, активируемым Дектин-1 рецепторы способны способствовать восстановлению аксонов, однако также вызывают нейротоксичность в нейроне.[40]
Регулировка длины
Аксоны в значительной степени различаются по длине от нескольких микрометров до метров у некоторых животных. Это подчеркивает, что должен существовать механизм регулирования длины клетки, позволяющий нейронам ощущать длину своих аксонов и соответственно контролировать их рост. Было обнаружено, что моторные белки играют важную роль в регулировании длины аксонов.[41] Основываясь на этом наблюдении, исследователи разработали четкую модель роста аксонов, описывающую, как моторные белки могут влиять на длину аксона на молекулярном уровне.[42][43][44][45] Эти исследования предполагают, что моторные белки переносят сигнальные молекулы от сомы к конусу роста и наоборот, концентрация которых колеблется во времени с частотой, зависящей от длины.
Классификация
Аксоны нейронов человека периферическая нервная система могут быть классифицированы по их физическим характеристикам и свойствам проводимости сигнала. Известно, что аксоны имеют разную толщину (от 0,1 до 20 мкм).[3] и считалось, что эти различия связаны со скоростью, с которой потенциал действия может перемещаться по аксону — его скорость проводимости. Эрлангер и Гассер доказали эту гипотезу и идентифицировали несколько типов нервных волокон, установив связь между диаметром аксона и его скорость нервной проводимости. Они опубликовали свои открытия в 1941 году, дав первую классификацию аксонов.
Аксоны подразделяются на две системы. Первый, введенный Эрлангером и Гассером, сгруппировал волокна в три основные группы, используя буквы A, B и C. группа А, группа B, и группа C включают оба сенсорных волокна (афференты ) и двигательные волокна (эфференты ). Первая группа A была разделена на альфа, бета, гамма и дельта волокна — Aα, Aβ, Aγ и Aδ. Моторные нейроны различных моторных волокон были нижние двигательные нейроны – альфа двигательный нейрон, бета мотонейрон, и гамма мотонейрон с нервными волокнами Aα, Aβ и Aγ соответственно.
Позже другие исследователи обнаружили две группы волокон Aa, которые были сенсорными волокнами. Затем они были введены в систему, которая включала только сенсорные волокна (хотя некоторые из них были смешанными нервами, а также двигательными волокнами). Эта система называет сенсорные группы Типами и использует римские цифры: Тип Ia, Тип Ib, Тип II, Тип III и Тип IV.
Мотор
Нижние двигательные нейроны имеют два вида волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин | Проведение скорость (м / с) |
Связанный мышечные волокна |
|---|---|---|---|---|---|
| α | Aα | 13-20 | да | 80–120 | Экстрафузионные мышечные волокна |
| β | Aβ | ||||
| γ | Aγ | 5-8 | да | 4–24[46][47] | Внутрифузионные мышечные волокна |
Сенсорный
Разные Рецепторы чувств иннервируют разные типы нервных волокон. Проприоцепторы иннервируются сенсорными волокнами типа Ia, Ib и II, механорецепторы сенсорными волокнами II и III типов и ноцицепторы и терморецепторы сенсорными волокнами III и IV типа.
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин | Проведение скорость (м / с) |
Связанный Рецепторы чувств | Проприоцепторы | Механоцепторы | Ноцицепторы и терморецепторы |
|---|---|---|---|---|---|---|---|---|
| Я | Aα | 13-20 | да | 80–120 | Первичные рецепторы мышечное веретено (аннулоспиральное окончание) | ✔ | ||
| Ib | Aα | 13-20 | да | 80–120 | Орган сухожилия Гольджи | |||
| II | Aβ | 6-12 | да | 33–75 | Вторичные рецепторы мышечное веретено (окончание цветочно-спрей). Все кожные механорецепторы |
✔ | ||
| III | Aδ | 1-5 | Тонкий | 3–30 | Свободные нервные окончания прикосновения и давления Ноцицепторы из боковой спиноталамический тракт Холодный терморецепторы |
✔ | ||
| IV | C | 0.2-1.5 | Нет | 0.5-2.0 | Ноцицепторы из передний спиноталамический тракт Рецепторы тепла |
Автономный
В автономная нервная система имеет два вида периферических волокон:
| Тип | Эрлангер-Гассер Классификация |
Диаметр (мкм) |
Миелин[48] | Проведение скорость (м / с) |
|---|---|---|---|---|
| преганглионарные волокна | B | 1–5 | да | 3–15 |
| постганглионарные волокна | C | 0.2–1.5 | Нет | 0.5–2.0 |
Клиническое значение
В порядке степени тяжести повреждение нерва можно описать как неврапраксия, аксонотмезис, или же невротмезис.Сотрясение считается легкой формой диффузное повреждение аксонов.[49] Аксональное повреждение также может вызвать центральный хроматолиз. Нарушение функции аксонов нервной системы является одной из основных причин многих наследственных заболеваний. неврологические расстройства которые влияют как на периферические, так и на центральные нейроны.[5]
Когда аксон раздавлен, активный процесс дегенерация аксонов происходит в части аксона, наиболее удаленной от тела клетки. Эта дегенерация происходит быстро после травмы, когда часть аксона блокируется мембранами и разрушается макрофагами. Это известно как Валлеровское вырождение.[50] Отмирание аксона также может иметь место при многих нейродегенеративных заболеваниях, особенно при нарушении транспорта аксонов, это известно как дегенерация, подобная валлеровской.[51] Исследования показывают, что дегенерация происходит в результате действия аксонального белка. NMNAT2, будучи не в состоянии достичь всего аксона.[52]
Демиелинизация аксонов вызывает множество неврологических симптомов, обнаруженных при заболевании рассеянный склероз.
Дисмиелинизация это аномальное образование миелиновой оболочки. Это связано с несколькими лейкодистрофии, а также в шизофрения.[53][54][55]
Суровый травматическое повреждение мозга может привести к обширным поражениям нервных путей, повреждая аксоны в состоянии, известном как диффузное повреждение аксонов. Это может привести к стойкое вегетативное состояние.[56] Это было показано в исследованиях на крыса такое повреждение аксонов в результате единичного легкого черепно-мозгового повреждения может оставить восприимчивость к дальнейшему повреждению после повторных легких черепно-мозговых травм.[57]
А нервный проводник является искусственным средством направления роста аксонов, чтобы нейрорегенерация, и является одним из многих методов лечения, используемых для различных видов повреждение нерва.
История
Немецкий анатом Отто Фридрих Карл Дайтерс обычно приписывают открытие аксона, отличив его от дендритов.[5] Швейцарский Рюдольф Альберт фон Кёлликер и немецкий Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Келликер назвал аксон в 1896 году.[58] Луи-Антуан Ранвье был первым, кто описал бреши или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлы Ранвье. Сантьяго Рамон-и-Кахаль, испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции.[5] Джозеф Эрлангер и Герберт Гассер ранее разработали систему классификации периферических нервных волокон,[59] на основе скорости аксональной проводимости, миелинизация, размер волокна и т. д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), а к 1952 году они получили полное количественное описание ионной основы потенциал действия, что привело к формулировке Модель Ходжкина – Хаксли. Ходжкин и Хаксли были награждены совместно Нобелевская премия для этой работы в 1963 году. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхойзера-Хаксли. Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает много деталей об отдельных ионные каналы.
Другие животные
Аксоны в беспозвоночные были широко изучены. В Прибрежный кальмар, часто используется как модельный организм имеет самый длинный из известных аксонов.[60] В Гигантский кальмар имеет самый большой аксон известен. Его размер варьируется от половины (обычно) до одного миллиметра в диаметре и используется для управления его реактивный двигатель система. Самая быстрая зарегистрированная скорость проводимости 210 м / с обнаружена в заключенных в оболочку аксонах некоторых пелагических Креветки Penaeid[61] и обычный диапазон составляет от 90 до 200 м / с.[62] (ср 100–120 м / с для аксона самого быстрого миелинизированного позвоночного.)
В других случаях, как показали исследования на крысах, аксон происходит от дендрита; такие аксоны, как говорят, имеют «дендритное происхождение». Некоторые аксоны с дендритным происхождением аналогичным образом имеют «проксимальный» начальный сегмент, который начинается непосредственно в источнике аксона, в то время как другие имеют «дистальный» начальный сегмент, заметно отделенный от источника аксона.[63] У многих видов некоторые из нейронов имеют аксоны, которые исходят из дендрита, а не из тела клетки, и они известны как дендриты, несущие аксоны.[1] Во многих случаях аксон берет свое начало от бугорка аксона на соме; такие аксоны, как говорят, имеют «соматическое происхождение». Некоторые аксоны соматического происхождения имеют «проксимальный» начальный сегмент, прилегающий к бугорку аксона, в то время как другие имеют «дистальный» начальный сегмент, отделенный от сомы протяженным бугорком аксона.[63]
Смотрите также
- Электрофизиология
- Ганглионарное возвышение
- Гигантская аксональная нейропатия
- Нейрональная трассировка
- Пионерский аксон
Рекомендации
- ^ а б Triarhou LC (2014). «Аксоны, исходящие из дендритов: филогенетические последствия с оттенками Кахаля». Границы нейроанатомии. 8: 133. Дои:10.3389 / fnana.2014.00133. ЧВК 4235383. PMID 25477788.
- ^ Яу К.В. (декабрь 1976 г.). «Рецептивные поля, геометрия и проводящий блок сенсорных нейронов центральной нервной системы пиявки». Журнал физиологии. 263 (3): 513–38. Дои:10.1113 / jphysiol.1976.sp011643. ЧВК 1307715. PMID 1018277.
- ^ а б c Сквайр, Ларри (2013). Фундаментальная нейробиология (4-е изд.). Амстердам: Elsevier / Academic Press. С. 61–65. ISBN 978-0-12-385-870-2.
- ^ а б c Людерс Э., Томпсон П.М., Тога А.В. (август 2010 г.). «Развитие мозолистого тела в мозгу здорового человека». Журнал неврологии. 30 (33): 10985–90. Дои:10.1523 / JNEUROSCI.5122-09.2010. ЧВК 3197828. PMID 20720105.
- ^ а б c d е Дебанн Д., Кампанак Е., Бяловас А., Карлье Е., Алькарас Г. (апрель 2011 г.). «Аксонная физиология» (PDF). Физиологические обзоры. 91 (2): 555–602. Дои:10.1152 / Physrev.00048.2009. PMID 21527732. S2CID 13916255.
- ^ Нельсон А.Д., Дженкинс П.М. (2017). «Аксональные мембраны и их домены: сборка и функция начального сегмента аксона и узла Ранвье». Границы клеточной неврологии. 11: 136. Дои:10.3389 / fncel.2017.00136. ЧВК 5422562. PMID 28536506.
- ^ Летерье С., Клерк Н., Руэда-Борони Ф, Монтерсино А., Дарджент Б., Кастетс Ф (2017). «Мембранные партнеры Ankyrin G стимулируют создание и поддержание начального сегмента Axon». Границы клеточной неврологии. 11: 6. Дои:10.3389 / fncel.2017.00006. ЧВК 5266712. PMID 28184187.
- ^ Летерье С (февраль 2018 г.). «Начальный сегмент аксона: обновленная точка зрения». Журнал неврологии. 38 (9): 2135–2145. Дои:10.1523 / jneurosci.1922-17.2018. ЧВК 6596274. PMID 29378864.
- ^ Расбанд М.Н. (август 2010 г.). «Начальный сегмент аксона и поддержание полярности нейронов». Обзоры природы. Неврология. 11 (8): 552–62. Дои:10.1038 / nrn2852. PMID 20631711. S2CID 23996233.
- ^ а б c d Джонс С.Л., Свиткина Т.М. (2016). «Цитоскелет начального сегмента аксона: архитектура, развитие и роль в полярности нейронов». Нейронная пластичность. 2016: 6808293. Дои:10.1155/2016/6808293. ЧВК 4967436. PMID 27493806.
- ^ Кларк Б.Д., Голдберг Е.М., Руди Б. (декабрь 2009 г.). «Электрогенная настройка начального сегмента аксона». Нейробиолог. 15 (6): 651–68. Дои:10.1177/1073858409341973. ЧВК 2951114. PMID 20007821.
- ^ а б Ямада Р., Куба Х (2016). «Структурная и функциональная пластичность в начальном сегменте аксона». Границы клеточной неврологии. 10: 250. Дои:10.3389 / fncel.2016.00250. ЧВК 5078684. PMID 27826229.
- ^ а б Сусуки К., Куба Х (март 2016 г.). «Активно-зависимая регуляция возбудимых аксональных доменов». Журнал физиологических наук. 66 (2): 99–104. Дои:10.1007 / s12576-015-0413-4. PMID 26464228. S2CID 18862030.
- ^ а б c d е Альбертс Б. (2004). Essential Cell Biology: введение в молекулярную биологию клетки (2-е изд.). Нью-Йорк: Гарленд. стр.584–587. ISBN 978-0-8153-3481-1.
- ^ а б Альбертс Б. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Гарленд. С. 979–981. ISBN 978-0-8153-4072-0.
- ^ Озген, Н; Барон, W; Hoekstra, D; Кахья, Н. (сентябрь 2016 г.). «Динамика олигодендроглиальной мембраны в связи с биогенезом миелина». Клеточные и молекулярные науки о жизни. 73 (17): 3291–310. Дои:10.1007 / s00018-016-2228-8. ЧВК 4967101. PMID 27141942.
- ^ Сэдлер, Т. (2010). Медицинская эмбриология Лангмана (11-е изд.). Филадельфия: Липпинкотт Уильям и Уилкинс. п.300. ISBN 978-0-7817-9069-7.
- ^ Гесс А., Янг Дж. З. (ноябрь 1952 г.). «Узлы Ранвье». Труды Лондонского королевского общества. Серия B, Биологические науки. Серия Б. 140 (900): 301–20. Bibcode:1952RSPSB.140..301H. Дои:10.1098 / rspb.1952.0063. JSTOR 82721. PMID 13003931. S2CID 11963512.
- ^ Роббинс А.А., Фокс С.Е., Холмс Г.Л., Скотт Р.К., Барри Дж. М. (ноябрь 2013 г.). «Кратковременные сигналы, записанные внеклеточно у свободно движущихся крыс, представляют активность аксонов». Границы в нейронных цепях. 7 (181): 181. Дои:10.3389 / fncir.2013.00181. ЧВК 3831546. PMID 24348338.
- ^ Rongjing Ge, Hao Qian и Jin-Hui Wang * (2011) Molecular Brain 4 (19), 1 ~ 11
- ^ Rongjing Ge, Hao Qian, Na Chen и Jin-Hui Wang * (2014) Molecular Brain 7 (26): 1-16
- ^ Чен Н, Ю Дж, Цянь Х, Ге Р, Ван Дж Х (июль 2010 г.). «Аксоны усиливают соматические неполные спайки до однородных амплитуд в пирамидных нейронах коры головного мозга мышей». PLOS ONE. 5 (7): e11868. Bibcode:2010PLoSO … 511868C. Дои:10.1371 / journal.pone.0011868. ЧВК 2912328. PMID 20686619.
- ^ Вольперт, Льюис (2015). Принципы развития (5-е изд.). С. 520–524. ISBN 978-0-19-967814-3.
- ^ Флетчер Т.Л., Банкир Г.А. (декабрь 1989 г.). «Установление полярности нейронами гиппокампа: взаимосвязь между стадией развития клетки in situ и ее последующим развитием в культуре». Биология развития. 136 (2): 446–54. Дои:10.1016/0012-1606(89)90269-8. PMID 2583372.
- ^ Цзян Х., Рао Й. (май 2005 г.). «Формирование аксона: судьба против роста». Природа Неврология. 8 (5): 544–6. Дои:10.1038 / nn0505-544. PMID 15856056. S2CID 27728967.
- ^ Гослин К., Банкир Г. (апрель 1989 г.). «Экспериментальные наблюдения за развитием полярности нейронов гиппокампа в культуре». Журнал клеточной биологии. 108 (4): 1507–16. Дои:10.1083 / jcb.108.4.1507. ЧВК 2115496. PMID 2925793.
- ^ Lamoureux P, Ruthel G, Buxbaum RE, Heidemann SR (ноябрь 2002 г.). «Механическое напряжение может определять судьбу аксонов в нейронах гиппокампа». Журнал клеточной биологии. 159 (3): 499–508. Дои:10.1083 / jcb.200207174. ЧВК 2173080. PMID 12417580.
- ^ а б Аримура Н., Кайбути К. (март 2007 г.). «Полярность нейронов: от внеклеточных сигналов к внутриклеточным механизмам». Обзоры природы. Неврология. 8 (3): 194–205. Дои:10.1038 / номер 2056. PMID 17311006. S2CID 15556921.
- ^ Нейроглия и пионерные нейроны express UNC-6 для предоставления глобальных и локальных сетевых сигналов для управления миграциями в C. elegans
- ^ Серафини Т., Кеннеди Т.Э., Галко М.Дж., Мирзаян С., Джессел Т.М., Тесье-Лавин М. (август 1994 г.). «Нетрины определяют семейство белков, способствующих отрастанию аксонов, гомологичных C. elegans UNC-6». Клетка. 78 (3): 409–24. Дои:10.1016/0092-8674(94)90420-0. PMID 8062384. S2CID 22666205.
- ^ Хонг К., Хинк Л., Нишияма М., Пу М.М., Тесье-Лавин М., Штейн Э. (июнь 1999 г.). «Управляемая лигандом ассоциация между цитоплазматическими доменами рецепторов семейства UNC5 и DCC превращает индуцированное нетрином притяжение конуса роста в отталкивание». Клетка. 97 (7): 927–41. Дои:10.1016 / S0092-8674 (00) 80804-1. PMID 10399920. S2CID 18043414.
- ^ Hedgecock EM, Culotti JG, Hall DH (январь 1990 г.). «Гены unc-5, unc-6 и unc-40 управляют периферической миграцией первичных аксонов и мезодермальных клеток по эпидермису C. elegans». Нейрон. 4 (1): 61–85. Дои:10.1016 / 0896-6273 (90) 90444-К. PMID 2310575. S2CID 23974242.
- ^ Хуанг Э.Дж., Райхардт Л.Ф. (2003). «Рецепторы Trk: роли в передаче сигнала нейронов». Ежегодный обзор биохимии. 72: 609–42. Дои:10.1146 / annurev.biochem.72.121801.161629. PMID 12676795. S2CID 10217268.
- ^ а б Да Силва Дж. С., Хасегава Т., Мияги Т., Дотти К. Г., Абад-Родригес Дж. (Май 2005 г.). «Активность асимметричной мембранной ганглиозидной сиалидазы определяет судьбу аксонов». Природа Неврология. 8 (5): 606–15. Дои:10.1038 / nn1442. PMID 15834419. S2CID 25227765.
- ^ Брадке Ф., Дотти К.Г. (март 1999 г.). «Роль локальной нестабильности актина в формировании аксонов». Наука. 283 (5409): 1931–4. Bibcode:1999Научный … 283.1931B. Дои:10.1126 / science.283.5409.1931. PMID 10082468.
- ^ Ферли А.Дж., Мортон С.Б., Манало Д., Карагогеос Д., Додд Дж., Джессел Т.М. (апрель 1990 г.). «Аксональный гликопротеин TAG-1 является членом суперсемейства иммуноглобулинов с активностью, способствующей росту нейритов». Клетка. 61 (1): 157–70. Дои:10.1016/0092-8674(90)90223-2. PMID 2317872. S2CID 28813676.
- ^ Альбертс, Брюс (2015). Молекулярная биология клетки (Шестое изд.). п. 947. ISBN 9780815344643.
- ^ Куник Д., Дион С., Одзаки Т., Левин Л.А., Константино С. (2011). «Лазерная пересечение одного аксона для исследований повреждения и регенерации аксонов с высоким содержанием аксонов». PLOS ONE. 6 (11): e26832. Bibcode:2011PLoSO … 626832K. Дои:10.1371 / journal.pone.0026832. ЧВК 3206876. PMID 22073205.
- ^ Schwab ME (февраль 2004 г.). «Регенерация ног и аксонов». Текущее мнение в нейробиологии. 14 (1): 118–24. Дои:10.1016 / j.conb.2004.01.004. PMID 15018947. S2CID 9672315.
- ^ Гензель Дж. К., Накамура С., Гуан З., ван Ройен Н., Анкени Д. П., Попович П. Г. (март 2009 г.). «Макрофаги способствуют регенерации аксонов с одновременной нейротоксичностью». Журнал неврологии. 29 (12): 3956–68. Дои:10.1523 / JNEUROSCI.3992-08.2009. ЧВК 2693768. PMID 19321792.
- ^ Майерс К.А., Баас П.В. (сентябрь 2007 г.). «Кинезин-5 регулирует рост аксона, действуя как тормоз на его массив микротрубочек». Журнал клеточной биологии. 178 (6): 1081–91. Дои:10.1083 / jcb.200702074. ЧВК 2064629. PMID 17846176.
- ^ Ришал И., Кам Н., Перри Р. Б., Шиндер В., Фишер Е. М., Скьяво Г., Файнзильбер М. (июнь 2012 г.). «Механизм с приводом от двигателя для измерения длины клетки». Отчеты по ячейкам. 1 (6): 608–16. Дои:10.1016 / j.celrep.2012.05.013. ЧВК 3389498. PMID 22773964.
- ^ Карамчед Б.Р., Бресслофф ПК (май 2015 г.). «Модель отсроченной обратной связи для измерения длины аксонов». Биофизический журнал. 108 (9): 2408–19. Bibcode:2015BpJ … 108.2408K. Дои:10.1016 / j.bpj.2015.03.055. ЧВК 4423051. PMID 25954897.
- ^ Бресслов П.С., Карамчед Б.Р. (2015). «Механизм частотно-зависимого декодирования для определения длины аксона». Границы клеточной неврологии. 9: 281. Дои:10.3389 / fncel.2015.00281. ЧВК 4508512. PMID 26257607.
- ^ Фольц Ф., Веттманн Л., Мориджи Г., Круз К. (май 2019 г.). «Звук роста аксона». Физический обзор E. 99 (5–1): 050401. arXiv:1807.04799. Bibcode:2019PhRvE..99e0401F. Дои:10.1103 / PhysRevE.99.050401. PMID 31212501.
- ^ Эндрю Б.Л., часть Нью-Джерси (апрель 1972 г.). «Свойства быстрых и медленных двигательных единиц в мышцах задних конечностей и хвоста крысы». Ежеквартальный журнал экспериментальной физиологии и когнитивных медицинских наук. 57 (2): 213–25. Дои:10.1113 / expphysiol.1972.sp002151. PMID 4482075.
- ^ Рассел Нью-Джерси (январь 1980 г.). «Скорость аксональной проводимости изменяется после тенотомии мышц или деафферентации во время развития у крысы». Журнал физиологии. 298: 347–60. Дои:10.1113 / jphysiol.1980.sp013085. ЧВК 1279120. PMID 7359413.
- ^ Покок Г., Ричардс С.Д. и др. (2004). Физиология человека (2-е изд.). Нью-Йорк: Издательство Оксфордского университета. С. 187–189. ISBN 978-0-19-858527-5.
- ^ Dawodu ST (16 августа 2017 г.). «Травматическая травма головного мозга (ЧМТ) — определение, эпидемиология, патофизиология». Medscape. В архиве с оригинала 12 июня 2018 г.. Получено 14 июля 2018.
- ^ Травма и валлеровская дегенерация В архиве 2 мая 2006 г. Wayback Machine, Калифорнийский университет в Сан-Франциско
- ^ Coleman MP, Freeman MR (1 июня 2010 г.). «Валлеровское вырождение, мир (а) и сущность». Ежегодный обзор нейробиологии. 33 (1): 245–67. Дои:10.1146 / annurev-neuro-060909-153248. ЧВК 5223592. PMID 20345246.
- ^ Джилли Дж., Член парламента Коулмана (январь 2010 г.). «Эндогенный Nmnat2 является важным фактором выживания для поддержания здоровья аксонов». PLOS Биология. 8 (1): e1000300. Дои:10.1371 / journal.pbio.1000300. ЧВК 2811159. PMID 20126265.
- ^ Кремер-Альберс Е.М., Гериг-Бургер К., Тиле С., Троттер Дж., Наве К.А. (ноябрь 2006 г.). «Нарушенные взаимодействия мутантного протеолипидного белка / DM20 с холестерином и липидными рафтами в олигодендроглии: последствия для дисмиелинизации при спастической параплегии». Журнал неврологии. 26 (45): 11743–52. Дои:10.1523 / JNEUROSCI.3581-06.2006. ЧВК 6674790. PMID 17093095.
- ^ Маталон Р., Михалс-Маталон К., Сурендран С., Тайринг С.К. (2006). «Болезнь Канавана: исследования на нокаутной мыши». N-ацетиласпартат. Adv. Exp. Med. Биол. Успехи экспериментальной медицины и биологии. 576. С. 77–93, обсуждение 361–3. Дои:10.1007/0-387-30172-0_6. ISBN 978-0-387-30171-6. PMID 16802706. S2CID 44405442.
- ^ Ткачев Д., Миммак М.Л., Хаффакер С.Дж., Райан М., Bahn S (август 2007 г.). «Дальнейшие доказательства изменения биосинтеза миелина и глутаматергической дисфункции при шизофрении». Международный журнал нейропсихофармакологии. 10 (4): 557–63. Дои:10.1017 / S1461145706007334. PMID 17291371.
- ^ «Травма головного мозга, травматическая». Медциклопедия. GE. Архивировано из оригинал 26 мая 2011 г.
- ^ Райт Д.К., Брэди Р.Д., Камнакш А., Трезизе Дж., Сан М., Макдональд С.Дж. и др. (Октябрь 2019 г.). «Повторные легкие черепно-мозговые травмы вызывают стойкие изменения белков плазмы и биомаркеров магнитно-резонансной томографии у крыс». Научные отчеты. 9 (1): 14626. Дои:10.1038 / с41598-019-51267-ш. ЧВК 6787341. PMID 31602002.
- ^ Палец S (1994). Истоки нейробиологии: история исследований функций мозга. Издательство Оксфордского университета. п. 47. ISBN 9780195146943. OCLC 27151391.
Келликер дал название аксону в 1896 году.
- ^ Грант G (декабрь 2006 г.). «Нобелевские премии 1932 и 1944 годов по физиологии и медицине: награды за новаторские исследования в области нейрофизиологии». Журнал истории неврологии. 15 (4): 341–57. Дои:10.1080/09647040600638981. PMID 16997762. S2CID 37676544.
- ^ Хелье, Дженнифер Л. (16 декабря 2014 г.). Мозг, нервная система и их болезни [3 тома]. ABC-CLIO. ISBN 9781610693387. В архиве из оригинала 14 марта 2018 г.
- ^ Хсу К., Теракава С. (июль 1996 г.). «Фенестрация в миелиновой оболочке нервных волокон креветки: новый узел возбуждения для скачкообразной проводимости». Журнал нейробиологии. 30 (3): 397–409. Дои:10.1002 / (SICI) 1097-4695 (199607) 30: 3 <397 :: AID-NEU8> 3.0.CO; 2- #. PMID 8807532.
- ^ Зальцер Дж. Л., Залц Б. (октябрь 2016 г.). «Миелинизация». Текущая биология. 26 (20): R971 – R975. Дои:10.1016 / j.cub.2016.07.074. PMID 27780071.
- ^ а б Хёффлин Ф., Джек А., Ридель С., Мак-Бухер Дж., Роос Дж., Корчелли С. и др. (2017). «Неоднородность начального сегмента аксона в интернейронах и пирамидных клетках зрительной коры грызунов». Границы клеточной неврологии. 11: 332. Дои:10.3389 / fncel.2017.00332. ЧВК 5684645. PMID 29170630.
внешняя ссылка
- Гистологическое изображение: 3_09 в Центре медицинских наук Университета Оклахомы — «Слайд 3 Спинной мозг «

![]()
Аксон 9-дневной мыши
Аксон (греч. ἀξον — ось) — нейрит, осевой цилиндр, отросток нервной клетки, по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.
Нейрон состоит из одного аксона, тела и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные, мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону, а затем сгенерированный потенциал действия от начального сегмента аксона передается назад к дендритам [1]. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС).
В месте соединения аксона с телом нейрона у наиболее крупных пирамидных клеток 5-ого слоя коры находится аксонный холмик. Ранее предполагалось, что здесь происходит преобразование постсинаптического потенциала нейрона в нервные импульсы, но экспериментальные данные это не подтвердили. Регистрация электрических потенциалов выявила, что нервный импульс генерируется в самом аксоне, а именно в начальном сегменте на расстоянии ~50 мкм от тела нейрона [2]. Для генерации потенциала действия в начальном сегменте аксона требуется повышенная концентрация натриевых каналов (до ста раз по сравнению с телом нейрона[3]).
Питание и рост аксона зависят от тела нейрона: при перерезке аксона его периферическая часть отмирает, а центральная сохраняет жизнеспособность. При диаметре в несколько микронов длина аксона может достигать у крупных животных 1 метра и более (например, аксоны, идущие от нейронов спинного мозга в конечности). У многих животных (кальмаров, рыб, кольчатых червей, форонид, ракообразных) встречаются гигантские аксоны толщиной в сотни мкм (у кальмаров — до 2—3 мм). Обычно такие аксоны отвечают за проведение сигналов к мышцам, обеспечивающим «реакцию бегства» (втягивание в норку, быстрое плавание и др.). При прочих равных условиях с увеличением диаметра аксона увеличивается скорость проведения по нему нервных импульсов.
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная (гладкая) эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна.
Миелиновая оболочка аксонов имеется только у позвоночных. Её образуют «накручивающиеся» на аксон специальные шванновские клетки (в центральной нервной системе — олигодендроциты), между которыми остаются свободные от миелиновой оболочки участки — перехваты Ранвье. Только на перехватах присутствуют потенциал-зависимые натриевые каналы и заново возникает потенциал действия. При этом нервный импульс распространяется по миелинизированным волокнам ступенчато, что в несколько раз повышает скорость его распространения. Скорость передачи сигнала по покрытым миелиновой оболочкой аксонам достигает 100 метров в секунду.[4]
Безмякотные аксоны меньше размерами чем аксоны покрытые миелиновой оболочкой, что компенсирует потери в скорости распространения сигнала по сравнению с мякотными аксонами.
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
Примечания
- ↑ Dendritic backpropagation and the state of the awa… [J Neurosci. 2007] — PubMed result
- ↑ Action potentials initiate in the axon initial seg… [J Neurosci. 2010] — PubMed result
- ↑ Action potential generation requires a high sodium… [Nat Neurosci. 2008] — PubMed result
- ↑ Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М., 1988
Ссылки
- Савельев А. В. Моделирование логики самоорганизации активности нервного пучка эфаптическими взаимодействиями аксонного уровня // сб.: Моделирование неравновесных систем. — Институт вычислительного моделирования СО РАН, Красноярск, 2004. — С. 142-143.
См. также
- Аксональный поиск пути
- Аксональный транспорт
- Аксон-рефлекс
- Конус роста
- Дендрит
- Валлерова дегенерация — при разрыве аксона
| |
|
|---|---|
| Нейроны (Серое вещество) |
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты) Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит, Базальный дендрит) типы: Биполярные нейроны · Псевдополярные нейроны · Мультиполярные нейроны · Пирамидальный нейрон · Клетка Пуркинье · Гранулярная клетка |
| Афферентный нерв/ Сенсорный нерв/ Сенсорный нейрон |
GSA · GVA · SSA · SVA · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено, II or Aβ, Aδ-волокна, C-волокна) |
| Эфферентный нерв/ Моторный нерв/ Моторный нейрон |
GSE · GVE · SVE · Верхний моторный нейрон · Нижний моторный нейрон (α мотонейроны, γ мотонейроны) |
| Синапс | Нейропиль · Синаптический пузырек · Нервно-мышечный синапс · Электрический синапс · Химический синапс · Интернейрон (Клетки Реншоу) |
| Сенсорный рецептор | Чувствительное тельце Мейснера · Нервное окончание Меркеля · Тельца Пачини · Окончание Руффини · Нервномышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица |
| Нейроглия | Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия |
| Миелин (Белое вещество) |
CNS: Олигодендроцит PNS: Клетки Шванна · Невролемма · Перехват Ранвье/Межузловой сегмент · Насечка миелина |
| Соединительная ткань | Эпиневрий · Периневрий · Эндоневрий · Нервные пучки · Мозговые оболочки: твёрдая, паутинная, мягкая |
-
Особенности строения и функции аксонов, аксонный транспорт.
Аксон
(греч.
ἀξον —
ось) — нейрит, осевой цилиндр, отросток
нервной клетки, по которому нервные
импульсы идут от тела клетки
(сомы) к иннервируемым органам и другим
нервным клеткам.
Нейрон состоит из
одного аксона, тела и нескольких
дендритов,
в зависимости от числа которых нервные
клетки делятся на униполярные, биполярные,
мультиполярные. Передача нервного
импульса происходит от дендритов (или
от тела клетки) к аксону, а затем
сгенерированный потенциал действия от
начального сегмента аксона передается
назад к дендритам . Если аксон в нервной
ткани соединяется с телом следующей
нервной клетки, такой контакт называется
аксо-соматическим, с дендритами —
аксо-дендритический, с другим аксоном —
аксо-аксональный (редкий тип соединения,
встречается в ЦНС).
В месте соединения
аксона с телом нейрона у наиболее крупных
пирамидных клеток 5-ого слоя коры
находится аксонный
холмик.
Ранее предполагалось, что здесь происходит
преобразование постсинаптического
потенциала нейрона в нервные импульсы,
но экспериментальные данные это не
подтвердили. Регистрация электрических
потенциалов выявила, что нервный импульс
генерируется в самом аксоне, а именно
в начальном сегменте на расстоянии ~50
мкм от тела нейрона. Для генерации
потенциала действия в начальном сегменте
аксона требуется повышенная концентрация
натриевых каналов (до ста раз по сравнению
с телом нейрона[3]).
Питание и рост
аксона зависят от тела нейрона: при
перерезке аксона его периферическая
часть отмирает, а центральная сохраняет
жизнеспособность. При диаметре в
несколько микронов
длина аксона может достигать у крупных
животных 1 метра и более (например,
аксоны, идущие от нейронов спинного
мозга
в конечности).
У многих животных (кальмаров,
рыб,
кольчатых
червей,
форонид,
ракообразных)
встречаются гигантские аксоны толщиной
в сотни мкм (у кальмаров — до 2—3 мм).
Обычно такие аксоны отвечают за проведение
сигналов к мышцам. обеспечивающим
«реакцию бегства» (втягивание в норку,
быстрое плавание и др.). При прочих равных
условиях с увеличением диаметра аксона
увеличивается скорость проведения по
нему нервных импульсов.
В протоплазме
аксона — аксоплазме — имеются
тончайшие волоконца — нейрофибриллы,
а также микротрубочки,
митохондрии
и агранулярная (гладкая) эндоплазматическая
сеть.
В зависимости от того, покрыты ли аксоны
миелиновой
(мякотной) оболочкой или лишены её, они
образуют мякотные или безмякотные
нервные
волокна.
Миелиновая оболочка
аксонов имеется только у позвоночных.
Её образуют «накручивающиеся» на аксон
специальные шванновские
клетки,
между которыми остаются свободные от
миелиновой оболочки участки —
перехваты
Ранвье.
Только на перехватах присутствуют
потенциал-зависимые натриевые каналы
и заново возникает потенциал
действия.
При этом нервный импульс распространяется
по миелинизированным волокнам ступенчато,
что в несколько раз повышает скорость
его распространения.
Концевые участки
аксона — терминали — ветвятся и
контактируют с другими нервными,
мышечными или железистыми клетками. На
конце аксона находится синаптическое
окончание —
концевой участок терминали, контактирующий
с клеткой-мишенью. Вместе с постсинаптической
мембраной клетки-мишени синаптическое
окончание образует синапс.
Через синапсы
передаётся возбуждение.
Специфическая
функция аксона — проведение потенциала
действия
от тела клетки к другим клеткам или
периферическим органам. Другая его
функция — аксонный
транспорт
веществ.
Помимо своей
специфической функции в качестве
проводника потенциалов
действия
аксон
является каналом для транспорта веществ.
Аксонный транспорт
— это перемещение веществ по аксону
. Белки, синтезированные в теле клетки,
синаптические
медиаторные вещества
и низкомолекулярные соединения
перемещаются по аксону вместе с клеточными
органеллами, в частности, митохондриями
. Для большинства веществ и органелл
обнаружен также транспорт в обратном
направлении. Вирусы
и токсины
могут проникать в аксон на его периферии
и перемещаться по нему. Аксонный транспорт
— активный процесс.
Аксонный транспорт
зависит от достаточного снабжения
энергией, при снижении уровня АТФ вдвое
аксонный транспорт блокируется, при
возобновлении доступа энергии —
возобновляется.
Белки цитоскелета
доставляются из тела клетки, двигаясь
по аксону со скоростью от 1 до 5 мм в
сутки. Это медленный
аксонный транспорт
(похожий на него транспорт имеется и в
дендритах). Многие ферменты и другие
белки цитозоля также переносятся при
помощи этого типа транспорта.
Нецитозольные
материалы, которые необходимы в синапсе,
такие как секретируемые белки и
мембраносвязанные молекулы, двигаются
по аксону с гораздо большей скоростью.
Эти вещества переносятся из места их
синтеза, эндоплазматического ретикулума,
к аппарату Гольджи, который часто
располагается у основания аксона. Затем
эти молекулы, упакованные в мембранные
пузырьки, переносятся вдоль
рельсов-микротрубочек путем быстрого
аксонного транспорта
со скоростью до 400 мм в сутки. Таким
образом по аксону транспортируются
митохондрии, различные белки, включая
нейропептиды (нейромедиаторы пептидной
природы), непептидные нейромедиаторы.
Транспорт материалов
от тела нейрона к синапсу называется
антероградным,
а в обратном направлении — ретроградным.
Транспорт по аксону
на большие расстояния происходит с
участием микротрубочек. Микротрубочки
в аксоне обладают присущей им полярностью
и ориентированны быстрорастущим
(плюс-)концом к синапсу, а медленнорастущим
(минус-) — к телу нейрона. Белки-моторы
аксонного транспорта принадлежат к
кинезиновому и динеиновому суперсемействам.
Кинезины являются,
в основном, плюс-концевыми моторными
белками, транспортирующими такие грузы,
как предшественники синаптических
везикул и мембранные органеллы. Этот
транспорт идет в направлению к синапсу
(антероградно). Цитоплазматические
динеины — это минус-концевые моторные
белки, транспортирующие нейротрофные
сигналы, эндосомы и другие грузы
ретроградно к телу нейрона. Ретроградный
транспорт осуществляется динеинами не
эксклюзивно: обнаружены несколько
кинезинов, перемещающихся в ретроградном
направлении.
-
Миенилизированные
и немиенилизированные волокна. Процесс
миелинизации.
Миелин
(в некоторых изданиях употребляется
некорректная теперь форма миэлин) —
вещество, образующее миелиновую
оболочку
нервных
волокон.
Миелиновая
оболочка —
электроизолирующая оболочка, покрывающая
аксоны
многих нейронов. Миелиновую оболочку
образуют глиальные клетки: в периферической
нервной системе — Шванновские
клетки,
в центральной нервной системе —
олигодендроциты.
Миелиновая оболочка формируется из
плоского выроста тела глиальной клетки,
многократно оборачивающего аксон
подобно изоляционной ленте. Цитоплазма
в выросте практически отсутствует, в
результате чего миелиновая оболочка
представляет собой, по сути, множество
слоёв клеточной мембраны.
Миелин прерывается
только в области перехватов Ранвье,
которые встречаются через правильные
промежутки длиной примерно 1 мм. В
связи с тем, что ионные токи не могут
проходить сквозь миелин, вход и выход
ионов осуществляется лишь в области
перехватов. Это ведёт к увеличению
скорости проведения нервного импульса.
Таким образом, по миелинизированным
волокнам импульс проводится приблизительно
в 5—10 раз быстрее, чем по немиелинизированным.
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Безмиелиновые
нервные волокна находятся преимущественно
в составе вегетативной нервной системы.
Клетки олигодендроглии оболочек
безмиелиновых нервных волокон,
располагаясь плотно, образуют тяжи, в
которых на определенном расстоянии
друг от друга видны овальные ядра. В
нервных волокнах внутренних органов,
как правило, в таком тяже располагается
не один, а несколько (10-20) осевых цилиндров,
принадлежащих различным нейронам. Они
могут, покидая одно волокно, переходить
в смежное, такие волокна, содержащие
несколько осевых цилиндров, называются
волокнами кабельного типа. При электронной
микроскопии безмиелиновых нервных
волокон видно, что по мере погружения
осевых цилиндров в тяж леммоцитов
последние одевают их как муфтой.
Оболочки леммоцитов
при этом прогибаются, плотно охватывают
осевые цилиндры и, смыкаясь над ними,
образуют глубокие складки, на дне которых
и располагаются отдельные осевые
цилиндры. Сближенные в области складки
участки оболочки нейролеммоцита образуют
сдвоенную мембрану — мезаксон, на которой
как бы подвешен осевой цилиндр. Оболочки
нейролеммоцитов очень тонкие, поэтому
ни мезаксона, ни границ этих клеток под
световым микроскопом нельзя рассмотреть,
и оболочка безмиелиновых нервных волокон
в этих условиях выявляется как однородный
тяж цитоплазмы, «одевающий» осевые
цилиндры. С поверхности каждое нервное
волокно покрыто базальной мембраной.
-
Миелиновые
– более толстые, т.к. содержат миелин-
находятся внутри под оболочкой; входят
в состав соматической нервной системы.
Миелин через несколько участков резко
истончается, образуя таким образом
перехват. Он называется перехват Ранвье.
Отросток между перехватами называется
межузловой сигмент. Миелин на подобии
жира. -
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Аксон
-
Аксон (др.-греч. ἄξων «ось») — это нейрит (длинный цилиндрический отросток нервной клетки), по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.
Каждый нейрон состоит из одного аксона, тела (перикариона) и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные или мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону, а затем сгенерированный потенциал действия от начального сегмента аксона передаётся назад к дендритам. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС).
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
Источник: Википедия
Связанные понятия
Си́напс (греч. σύναψις, от συνάπτειν — соединение, связь) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём, посредством прохождения ионов из одной клетки в другую.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться один с другим, формируя биологические…
Дендрит (от греч. δένδρον (dendron) — дерево) — разветвлённый отросток нейрона, который получает информацию через химические (или электрические) синапсы от аксонов (или дендритов и сомы) других нейронов и передаёт её через электрический сигнал телу нейрона (перикариону), из которого вырастает. Термин «дендрит» ввёл в научный оборот швейцарский ученый В. Гис в 1889 году.
Миелин (в некоторых изданиях употребляется некорректная теперь форма миэлин) — вещество, образующее миелиновую оболочку нервных волокон.
Вставочный нейрон (синонимы: интернейрон, промежуточный нейрон; англ. interneuron, relay neuron, association neuron, bipolar neuron) — нейрон, связанный только с другими нейронами, в отличие от двигательных нейронов, иннервирующих мышечные волокна, и сенсорных нейронов, преобразующих стимулы из внешней среды в электрические сигналы.
Упоминания в литературе
Аксон — длинное нервное волокно, отходящее от тела некоторых нейронов и заканчивающееся терминалями (до нескольких сотен) на клетках-мишенях: нейронах, миоцитах, клетках железы и др. В зависимости от наличия или отсутствия аксона выделяют нейроны I и II типа. Область тела нейрона, которой начинается аксон, называют начальным сегментом аксон, или аксонным холмиком. Мембрана аксонного холмика генерирует потенциалы действия (нервные импульсы), распространяющиеся по аксону. Некоторые аксоны покрыты прерывистыми цепочками миелиновых оболочек (муфт) и называются миелинизированными (волокна спинномозговых нервов). Аксоны, свободные от миелина, называются немиелинизированными (волокна вегетативных нервов). Между муфтами имеются промежутки – перехваты Ранвье. В участках, лишенных миелиновой оболочки, мембрана аксона контактирует непосредственно с внеклеточной средой. В ЦНС миелиновые муфты образованы глиальными клетками, олигодендритами. Миелиновые муфты аксонов, выходящих за пределы ЦНС (периферических волокон), образованы шванновскими клетками. Одна шванновская клетка может образовывать муфты, покрывая до девяти нервных волокон. Главная функция аксонов – передача нервных импульсов от тела нейрона к терминалям, которые заканчиваются на клетках-мишенях утолщениями, синаптическими бляшками. Скорость движения информации по аксонам составляет от нескольких миллиметров в секунду в мелких немиелинизированных волокнах до 90 м/с в крупных миелинизированных. Миелиновая оболочка увеличивает скорость передачи потенциалов действия по аксону.
НЕЙРО́Н, нервная клетка; основная структурная и функциональная единица нервной системы. Нейроны обладают особыми свойствами – возбудимостью и проводимостью. Из нейронов построена нервная ткань. Как и другие клетки, нейрон имеет ядро, цитоплазму и клеточную мембрану. Ядро содержится в теле нейрона (его расширенная часть), от которого отходят отростки – короткие (дендриты) и один длинный (аксон). Длина аксона может достигать 1 м и более. Аксоны образуют тяжки, которые называются нервами (наиболее крупные – нервными стволами). Дендриты принимают сигналы из внешней среды или от другой нервной клетки и проводят возбуждение к аксону, который передаёт его в удалённые от тела нейрона области. Тела нейронов образуют серое вещество головного и спинного мозга, а дендриты и аксоны – белое мозговое вещество.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка – интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
Нейрон. Основной функциональной и структурной единицей нервной системы является нейрон, строение которого показано на рис. 1.1. Нейрон состоит из клеточного тела, или сомы, коротких нервных отростков, называемых дендритами, и длинного нервного волокна – аксона. Область выхода аксона из клеточного тела нейрона называется аксонным холмиком. Хотя формально и дендриты и аксоны являются нервными волокнами, термин «нервное волокно» в основном употребляется при упоминании аксона. Дендриты передают нервный импульс к телу нейрона, тогда как аксон направляет импульс из тела нейрона на другие нервные клетки или на мышечные волокна.
Связанные понятия (продолжение)
Клетки Пуркинье (англ. Purkinje cells) — крупные нервные клетки коры мозжечка. Своё название клетки получили в честь их первооткрывателя, чешского врача и физиолога Яна Эвангелисты Пуркинье.
Пирамидальные нейроны, или пирамидные нейроны, — основные возбудительные нейроны мозга млекопитающих. Также обнаруживаются у рыб, птиц, рептилий. Напоминают по форме пирамиду, из которой вверх ведёт большой апикальный дендрит; имеют один аксон, идущий вниз, и множество базальных дендритов. Впервые были исследованы Рамон-и-Кахалем. Отмечены в таких структурах, как кора мозга, гиппокамп, миндалевидное тело (амигдала), но отсутствуют в обонятельной луковице, стриатуме, среднем мозге, ромбовидном мозге…
Подробнее: Пирамидальный нейрон
Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов. Термин ввёл в 1846 году Рудольф Вирхов.
Перикарион (др.-греч. περι- — приставка со значением «около, вокруг, кругом» + κάρυον — «орех») — сома (тело) нейрона, может иметь различную величину и форму. На цитолемме перикариона образуются многочисленные синаптические контакты с отростками других нейронов.
Нервная ткань — ткань эктодермального происхождения, представляет собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию.
Мотонейро́н (от лат. motor — приводящий в движение и нейрон; двигательный нейро́н) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса.
Шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Астроцит (лат. astrocytus; от греч. astron — звезда; и kýtos, здесь — клетка) — тип нейроглиальной клетки звездчатой формы с многочисленными отростками. Совокупность астроцитов называется астроглией.
Гранулярные клетки — несколько разновидностей мелких нейронов мозга. Название «гранулярная клетка» («зернистая клетка», «клетка-зерно») используется анатомами для нескольких разных типов нейронов, единственной общей особенностью которых является крайне малый размер тел этих клеток.
Подробнее: Гранулярная клетка
Гипотала́мус (лат. hypothalamus, от греч. ὑπό — «под» и θάλαμος — «комната, камера, отсек, таламус») — небольшая область в промежуточном мозге, включающая в себя большое число групп клеток (свыше 30 ядер), которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма. Гипоталамус связан нервными путями практически со всеми отделами центральной нервной системы, включая кору, гиппокамп, миндалину, мозжечок, ствол мозга и спинной мозг. Вместе с гипофизом гипоталамус образует гипоталамо-гипофизарную…
Не́рвные воло́кна — длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие.
Корзинчатые нейроны — тормозные ГАМК-эргические вставочные нейроны молекулярного слоя мозжечка. Длинные аксоны корзинчатых нейронов образуют корзиноподобные синапсы с телами клеток Пуркинье. Корзинчатые нейроны многополярны, их дендриты свободно ветвятся.
Подробнее: Корзинчатый нейрон
Тала́мус, иногда — зри́тельные бугры (лат. Thalamus; от др.-греч. θάλαμος — «камера, комната, отсек») — отдел головного мозга, представляющий собой большую массу серого вещества, расположенную в верхней части таламической области промежуточного мозга хордовых животных, в том числе и человека. Впервые описан древнеримским врачом и анатомом Галеном. Таламус — это парная структура, состоящая из двух половинок, симметричных относительно межполушарной плоскости. Таламус находится глубже структур большого…
Полоса́тое те́ло (лат. corpus striatum) — анатомическая структура конечного мозга, относящаяся к базальным ядрам полушарий головного мозга.
Гиппока́мп (от др.-греч. ἱππόκαμπος — морской конёк) — часть лимбической системы головного мозга (обонятельного мозга). Участвует в механизмах формирования эмоций, консолидации памяти (то есть перехода кратковременной памяти в долговременную). Генерирует тета-ритм при удержании внимания.
Латеральное коленчатое тело (наружное коленчатое тело, ЛКТ) — легко распознаваемая структура мозга, которая помещается на нижней латеральной стороне подушки таламуса в виде достаточно большого плоского бугорка. В ЛКТ приматов и человека морфологически определено шесть слоев: 1 и 2 — слои крупных клеток, 3-6 — слои мелких клеток. Слои 1, 4 и 6 получают афференты от контрлатерального (расположенного в противоположном по отношению к ЛКТ полушарии) глаза, а слои 2, 3 и 5 — от ипсилатерального (расположенного…
Зубчатая извилина (лат. gyrus dentatus) или зубчатая фасция гиппокампа (лат. fascia dentata hippocampi) — зазубренная извилина, расположенная в глубине борозды гиппокампа и переходящая в ленточную извилину. В некоторых классификациях она вместе с аммоновым рогом считается частью самого гиппокампа, однако большинство авторов относят её к гиппокамповой формации. В её структуре выделяют три слоя: полиморфный хилус, гранулярный слой и молекулярный слой, который непрерывно переходит в молекулярный слой…
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в виде кратковременного изменения мембранного потенциала на небольшом участке возбудимой клетки (нейрона или кардиомиоцита), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к внутренней поверхности мембраны, в то время, как в покое она заряжена положительно. Потенциал действия является физиологической основой нервного импульса.
Обонятельная луковица — часть обонятельного мозга, парное образование, состоящее из тел вторых нейронов биполярного типа обонятельного анализатора. Располагается во внутричерепной полости между лобной долей сверху и решётчатой пластинкой решётчатой кости снизу, через отверстие которой в неё поступают нервные волокна обонятельной области носа, а сзади продолжается в обонятельный тракт.
Реце́птор — объединение из терминалей (нервных окончаний) дендритов чувствительных нейронов, глии, специализированных образований межклеточного вещества и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражитель) в нервный импульс. В некоторых рецепторах (например, вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками эпителиального происхождения или…
Ганглий (др.-греч. γάγγλιον — узел), или нервный узел — скопление нервных клеток, состоящее из тел, дендритов и аксонов нервных клеток и глиальных клеток. Обычно ганглий имеет также оболочку из соединительной ткани. Имеются у многих беспозвоночных и всех позвоночных животных. Часто соединяются между собой, образуя различные структуры (нервные сплетения, нервные цепочки и т. п.).
Голубое пятно (голубоватое место, голубоватое пятно, синее пятно/место, лат. locus coeruleus) — ядро, расположенное в стволе мозга на уровне моста (участок голубоватого цвета в верхнелатеральной части ромбовидной ямки ствола головного мозга кнаружи от верхней ямки), часть ретикулярной формации. Система его проекций очень широка — аксоны восходят к верхним слоям коры больших полушарий, гиппокампу, миндалине, перегородке, полосатому телу, коре мозжечка. Нисходящие проекции идут в спинной мозг к симпатическим…
Афферентация (от лат. afferens — «приносящий») — постоянный поток нервных импульсов, поступающих в центральную нервную систему от органов чувств, воспринимающих информацию как от внешних раздражителей (экстерорецепция), так и от внутренних органов (интерорецепция). Находится в прямой зависимости от количества и силы воздействующих раздражителей, а также от состояния — активности или пассивности — индивида.
Белое вещество (лат. substantia alba) — компонент центральной нервной системы позвоночных животных и человека, состоящий главным образом из пучков аксонов, покрытых миелином. Противопоставляется серому веществу мозга, состоящему из клеточных тел нейронов. Цветовая дифференциация белого и серого вещества нервной ткани обусловлена белым цветом миелина.
Нервно-мышечный синапс (также нейромышечный, либо мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в этом синапсе является ацетилхолин.
Ретикулярная формация (лат. reticulum — сеточка, formatio — образование) — это образование, тянущееся вдоль всей оси ствола головного мозга. Своим названием оно обязано сетчатой структуре, образуемой его нервными клетками с очень сложными связями. Формация состоит из ретикулярных ядер и большой сети нейронов с разветвлёнными аксонами и дендритами, представляющих единый комплекс, который осуществляет активацию коры головного мозга и контролирует рефлекторную деятельность спинного мозга. Эта сеть нейронов…
Средний мозг, или мезэнцефалон (англ. Midbrain, лат. Mesencephalon; термин «мезэнцефалон» происходит от др.-греч. μέσος — «месос» — средний, и ἐγκέφᾰλος — «энкефалос» — буквально «находящийся внутри головы», то есть головной мозг) — это отдел головного мозга хордовых животных, развивающийся из среднего из трёх первичных мозговых пузырей эмбриона. Этот отдел мозга ответствен за осуществление многих важных физиологических функций, таких, как зрение, слух, контроль движений, регуляция циклов сна и бодрствования…
Мозжечо́к (лат. cerebellum — дословно «малый мозг») — отдел головного мозга позвоночных, отвечающий за координацию движений, регуляцию равновесия и мышечного тонуса. У человека располагается позади продолговатого мозга и варолиева моста, под затылочными долями полушарий головного мозга. Посредством трёх пар ножек мозжечок получает информацию из коры головного мозга, базальных ганглиев, экстрапирамидной системы, ствола головного мозга и спинного мозга. У различных таксонов позвоночных взаимоотношения…
Мозгово́й ствол, или ствол головного мозга, — традиционно выделяющийся отдел третьего мозга, представляющий собой протяжённое образование, продолжающее спинной мозг.
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг…
Конечный мозг (лат. telencephalon) — самый передний отдел головного мозга. Состоит из двух полушарий большого мозга (покрытых корой), мозолистого тела, полосатого тела и обонятельного мозга. Является наиболее крупным отделом головного мозга. Это также самая развитая структура, покрывающая собой все отделы головного мозга.
Дендритный шипик — мембранный вырост на поверхности дендрита, способный образовать синаптическое соединение. Шипики обычно имеют тонкую дендритную шейку, оканчивающуюся шарообразной дендритной головкой. Дендритные шипики обнаруживаются на дендритах большинства основных типов нейронов мозга. В создании шипиков участвует белок калирин.
Эпиталамус (буквально «надталамус») — это самая дорсальная (верхняя) задняя часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо эпиталамуса, входят также таламус, субталамус и метаталамус, но не входят гипоталамус и гипофиз, причисляемые к гипоталамической области. Эпиталамус возвышается над таламусом. В число его структур входят поводок эпиталамуса, также называемый поводком мозга, треугольник поводка, спайка поводков, подспаечный орган…
Передний мозг, или прозэнцефалон (лат. prosencephalon, англ. forebrain) — это отдел головного мозга хордовых животных, выделяемый на основании особенностей его эмбрионального развития у этих видов животных. Передний мозг (прозэнцефалон) является одним из трёх первичных мозговых пузырей, образующихся в первичной нервной трубке вскоре после нейруляции и образования нейромер, на так называемой трёхпузырьковой стадии развития ЦНС эмбриона. Двумя другими первичными мозговыми пузырями являются средний…
Миоци́ты, или мы́шечные клетки — особый тип клеток, составляющий основную часть мышечной ткани. Миоциты представляют собой длинные, вытянутые клетки, развивающиеся из клеток-предшественников — миобластов. Существует несколько типов миоцитов: миоциты сердечной мышцы (кардиомиоциты), скелетной и гладкой мускулатуры. Каждый из этих типов обладает особыми свойствами. Например, кардиомиоциты, помимо прочего, генерируют электрические импульсы, задающие сердечный ритм.
Головно́й мозг (лат. cerebrum, др.-греч. ἐγκέφαλος) — главный орган центральной нервной системы подавляющего большинства хордовых, её головной конец; у позвоночных находится внутри черепа. В анатомической номенклатуре позвоночных, в том числе человека, мозг в целом чаще всего обозначается как encephalon — латинизированная форма греческого слова; изначально латинское cerebrum стало синонимом большого мозга (telencephalon).
Спинно́й мозг (лат. medulla spinalis) — орган центральной нервной системы позвоночных, расположенный в позвоночном канале. Принято считать, что граница между спинным и головным мозгом проходит на уровне перекреста пирамидных волокон (хотя эта граница весьма условна). Внутри спинного мозга имеется полость, называемая центральным каналом (лат. canalis centralis). Спинной мозг защищён мягкой, паутинной и твёрдой мозговой оболочкой. Пространства между оболочками и спинномозговым каналом заполнены спинномозговой…
Бледный шар (лат. globus pallidus s. pallidum) — парная структура переднего мозга, относящаяся к базальным ядрам, часть чечевицеобразного ядра, вентромедиальная часть полосатого тела. Подразделяется на латеральную и медиальную части.
Промежуточный мозг, или диэнцефалон (лат. Diencephalon, англ. Diencephalon; термин «диэнцефалон» происходит от др.-греч. διά — «диа-», обозначающее «через», «между», и ἐγκέφαλος — «энкефалос», буквально «находящийся внутри головы», то есть головной мозг) — отдел головного мозга хордовых животных, который образуется в процессе эмбрионального развития из задней части зародышевого переднего мозга (прозэнцефалона). На пятипузырьковой стадии из задней части зародышевого переднего мозга (прозэнцефалона…
Олигодендроциты, или олигодендроглия — это вид нейроглии, открытый Пио дель Рио-Ортегой (1928 год). Олигодендроциты есть только в центральной нервной системе, которая у позвоночных включает в себя головной мозг и спинной мозг.
Синаптогенез — процесс формирования синапсов между нейронами в нервной системе. Синаптогенез происходит на протяжении всей жизни здорового человека, а взрыв формирования синапсов, т. н. избыточный синаптогенез (exuberant synaptogenesis), наблюдается на ранних стадиях развития головного мозга. Синаптогенез особенно важен в ходе критического периода развития особи (в биологии развития, такого периода, когда нервная система особенно чувствительна к экзогенным стимулам), когда имеет место быть интенсивное…
Хвостатое ядро (лат. nucleus caudatus) — парная структура головного мозга, относящаяся к стриатуму. Расположена спереди от таламуса, от которого (на горизонтальном срезе) его отделяет белая полоска вещества — внутренняя капсула. Передний отдел хвостатого ядра утолщён и образует головку, caput nuclei caudati, которая составляет латеральную стенку переднего рога бокового желудочка. Головка хвостатого ядра примыкает внизу к переднему продырявленному веществу, в этом месте головка соединяется с чечевицеобразным…
Продолговатый мозг (лат. myelencephalon, medulla oblongata), или луковица головного мозга (лат. bulbus cerebri), — задний отдел головного мозга, непосредственное продолжение спинного мозга. Происходит из ромбовидного мозга и входит в ствол головного мозга. Регулирует такие основные процессы жизнедеятельности, как дыхание и кровообращение, поэтому в случае повреждения продолговатого мозга мгновенно наступает смерть.
Химический синапс — особый тип межклеточного контакта между нейроном и клеткой-мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество.
Электри́ческий си́напс (англ. electrical synapse) — место высокоспециализированных контактов (щелевых контактов) между нейронами, где происходит прямое перетекание электрических токов от одного нейрона к другому. В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм, в то время как в химическом синапсе расстояние между двумя нейронами составляет от 20 до 40 нм. У многих животных в нервной системе имеются как химические, так и электрические синапсы. По сравнению с химическими…
Упоминания в литературе (продолжение)
Основная единица нервной системы — нервная клетка (нейрон). Нейроны покрыты глиальными клетками, доставляющими им питание и удерживающими их на месте. Каждая нервная клетка состоит из тела, дендрита и аксона. В периферической нервной системе аксоны собраны в пучки – нервы. Бо́льшая часть аксонов покрыта миелиновой оболочкой, которая увеличивает скорость передачи информации в нервной системе, а также препятствует передаче нервных импульсов близлежащим клеткам. Современные исследователи опровергли бытовавшее ранее мнение о том, что «нервные клетки не восстанавливаются». Доказано, что клетки в зонах мозга, связанных с памятью и процессом обучения, продолжают развиваться и делиться в течение жизни человека.
Нервные клетки имеют несколько отростков – чувствительных древовидно ветвящихся дендритов, которые проводят к телу нейронов возбуждение, возникающее на их чувствительных нервных окончаниях, расположенных в органах, и одних двигательный аксон, по которому нервный импульс передается от нейрона к рабочему органу или другому нейрону. Нейроны вступают друг с другом в контакт с помощью окончаний отростков, образуя рефлекторные цепи, по которым передаются (распространяются) нервные импульсы.
От нервных центров по основному отростку (аксону) идут сигналы к каждой клетке и каждому органу тела, заставляя их путем электрической стимуляции выполнять определенную функцию. Нервные центры состоят из сотен и даже тысяч нервных клеток. Соответственно, существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Основной ее элемент – нейрон: микроскопически малая нервная клетка с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Вспомогательные клетки нейроглии (астроциты) не только создают физическую опору для нейронов, но вместе с сосудами обеспечивают потребности нервной ткани в кислороде и необходимых для жизни веществах, включая аминокислоты, липиды, гликопротеиды. Тело нервной клетки имеет микроскопические размеры, но длина аксона может достигать одного метра! Отростки нейронов, как правило, укрыты миелиновой оболочкой, которая обеспечивает стабильность обмена веществ в длинных нервных проводниках и высокую скорость передачи возбуждения.
Нейрон представляет собой микроскопически малую нервную клетку с отходящими от нее отростками. Длинный отросток называется аксоном, короткий древовидной формы – дендритом. Посредством этих отростков все нервные клетки связаны между собой. Место соединения отростков называется синапсом. Пучки нервных волокон, идущие к поверхности тела, к внутренним органам и мышцам, образуют периферическую нервную систему. Разветвленные окончания нервных волокон образуют органы чувств.
Главная структурная и функциональная единица нервной системы – нейрон, представляющий собой клетку с отростками – длинным аксоном и короткими дендритами. Нейроны соединяются между собой синапсами, образуя нейронные цепи, приводимые в действие рефлекторно: в ответ на раздражение, поступаемое из внешней или внутренней среды, возбуждение из нервных окончаний передается по центростремительным волокнам в головной и спинной мозг, оттуда импульсы по центробежным волокнам поступают в различные органы, а по двигательным – к мышцам.
Аксон – вырост цитоплазмы, приспособленный для проведения информации, которая собирается ден-дритами и перерабатывается в нейроне. Аксон дендритной клетки имеет постоянный диаметр и покрыт миелиновой оболочкой, которая образована из глии, у аксона разветвленные окончания, в которых находятся митохондрии и секреторные образования.
Два слова о строении нейрона. У него есть центральная толстенькая часть – «тело», в котором находится клеточное ядро с генами. От тела отходят два вида отростков: «входные» (дендриты) и «выходные» (аксоны). Дендритов обычно много, а аксон, как правило, один, но на конце он может ветвиться.
При поражении центров коры головного мозга очень важна локализация, ведь повреждения происходят на микроскопическом уровне. Головной мозг состоит из нейронов и глиальных клеток, которые соединены между собой сложными проводимыми путями – аксонами и дендритами. По ним передаются нервные импульсы. Все мы еще со школьного курса биологии знаем, что в головном мозге есть серое и белое вещества. Серым веществом является скопление нейронов и коротких проводимых путей между ними. А вот белое вещество составляют длинные, сложные проводимые пути – нервные волокна, которые покрыты особой миелиновой оболочкой – она необходима для нормального прохождения импульсов и подобна изоляционной ленте на электрическом проводе.
В состоянии относительного физиологического покоя синапс находится в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается и ее содержимое в виде одного кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
Таким образом, нейрофизиологические и психофизиологические механизмы tDCS и ТКМП достаточно многообразны, даже при приложении стандартизированных параметров тока и площади электродов (при tDCS). Нам представляется маловероятным сведение механизмов поляризаций к делоляризации или гиперполяризации мембраны нейронов. Если для пирамидных нейронов с их пространственной ориентацией в коре можно себе представить, что, например, при анодной поляризации входящий (гиперполяризующий) ток протекает преимущественно через дендрита, а выходящий (деполяризующий) ток – через начальный сегмент аксона (где генерируются спайки), вызывая его деполяризацию и увеличение возбудимости моторной коры, то это допущение плохо приложимо к другим областям коры, где ориентация нейронов не столь очевидна. Кроме того, возбуждающие или тормозные потенциалы длятся не более 10 мсек. Пассивная деполяризация мембраны еще короче. Установленные эффекты tDCS длятся десятки минут, а то и дольше. Следовательно, де– или гиперполяризация, если и играют существенную роль в возникновении эффектов, то она сводится к пусковой роли, запускающей каскад синаптических и метаболических реакций которые и лежат в основе более длительных эффектов поляризаций.
Надо сказать, что нейрон устроен очень интересно: у него, как у всякого компьютера, есть корпус – тело. И все же славится нейрон не этим, главное в нем – это его отростки. Весь нейрон чем-то напоминает дерево, которое имеет развитую крону и ствол. Крону у дерева образуют ветки, а у нейрона это короткие отростки, они называются дендритами. У дерева ствол, а у нейрона – длинный отросток, аксон. А теперь о месте соединения нервных клеток – это синапс. Если два компьютера соединить шнуром, то контакт компьютера и шнура будет как раз тем самым синапсом в нервной системе.
Активность генов в нейронах основана на свойствах молекул ДНК, т. е. активность вызвана причинно-следственными связями, которые определяют особенности контактов молекул ДНК с окружающими атомами и молекулами. В свою очередь, активность генов является причиной всех событий клеточного метаболизма и, значит, обмена веществ между нейронами и с окружающей средой. Все процессы в сообществе нервных клеток (клеточном сообществе – КС) – рождение клеток, их возбуждение и торможение, функциональная специализация и формирование в них устойчивых молекулярных структур, синтез в клетках нейромедиаторов и образование у них аксонов, дендритов и шипиков, образование и разрыв межклеточных связей, распространение и прекращение возбуждений в сообществе, гибель клеток – это детерминистские процессы. Так у нейронов в течение суток изменяются параметры метаболизма: количество и соотношение синтезируемых и разлагающихся веществ, в т. ч. количество синтезируемых нейромедиаторов и гормонов. У нейронов, как и у всех организмов, имеются суточные ритмы. В соответствии с этими ритмами в продолжение суток изменяются состав и количество клеток, возбуждающихся от воздействий окружающей среды и следов таких воздействий, результаты конкуренции клеток и узлов за прекращение своего возбуждения, используемые возбуждениями маршруты связей между клетками, конфигурация формирующихся способов прекращения возбуждений, распространяющихся из одних и тех же узлов.
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь. Коллатерали, идущие от задних корешков шейных сегментов, входят в состав клиновидного пучка, идут вверх к его ядру и к дополнительному клиновидному ядру. Его аксоны соединяются с мозжечком. Третья группа коллатеральных афферентных волокон заканчивается в задних рогах спинного мозга. Там расположены вторые нейроны, чьи аксоны образуют передний спино-мозжечковый путь.
Соответственно существует такое же количество аксонов. Они собираются в пучки (так называемые тракты), которые, соединяясь вместе, образуют общий «кабель» – спинной мозг.
Двигательная единица – это отдельный мотонейрон и мышечные волокна, которые он иннервирует. Таким образом, нейрон определяет, являются ли волокна медленно или быстро сокращающимися. Мотонейрон в МС двигательной единице имеет небольшое клеточное тело и иннервирует группу из 10-180 мышечных волокон. У мотонейрона в БС двигательной единице большое клеточное тело и больше аксонов, и он иннервирует от 300 до 800 мышечных волокон. Отсюда следует, что каждый МС-мотонейрон в состоянии активировать значительно меньшее количество мышечных волокон в противоположность БС-мотонейрону. При этом необходимо отметить, что сила, производимая отдельными МС- и БС-волокнами по величине отличается незначительно. МС- и БС-волокна имеют разные функции во время физической активности. МС-волокнам присущ высокий уровень аэробной выносливости, они эффективны в производстве АТФ на основе окисления углеводов и жиров и более приспособлены к выполнению длительной работы невысокой интенсивности. Быстро сокращающиеся мышечные волокна приспособлены к анаэробной деятельности (без кислорода), и при их работе АТФ образуется благодаря анаэробным реакциям. Б С двигательные единицы производят большую силу, однако легко устают ввиду ограниченной выносливости и используются главным образом при выполнении кратковременной работы высокой интенсивности.
• длинный отросток, отходящий от тела клетки, который тянется на большое расстояние – до 1,5–1,7 м. Он составляет основной, или осевой, отросток нервной клетки. Его называют аксоном (в переводе с латыни axis — ось, основание, основной).
Другая часть коллатералей соединяется с клетками ядра Кларка, расположенного в заднем роге спинного мозга. Данное ядро расположено от VIII шейного до II поясничного сегментов по длиннику спинного мозга. Клетки грудного ядра являются вторыми нейронами, чьи аксоны образуют задний спиномозжечковый путь.
Формирование мышечной ткани начинается на 4-6-й неделе внутриутробного развития. В это время формируются первичные мышечные волокна. Несколько позже в мышцы прорастают аксоны мотонейронов спинного мозга. С этой стадии начинается синхронное формирование нервно-мышечного аппарата, причем определяющее значение имеет развитие нервных элементов, происходящее на 6-7-м месяцах внутриутробного развития. К моменту рождения примерно половина мышечных волокон уже прошла стадию первичной дифференцировки, и уже определены как «белые» или «красные». Дифференцировочные процессы усиливаются в возрасте от 1 до 2 лет, а затем на стадии полового созревания.
Нейросекреторные клетки, как и обычные нервные клетки, воспринимают сигналы, поступающие к ним от других отделов нервной системы, но далее передают полученную информацию уже гуморальным путем (не по аксонам, а по сосудам) – посредством нейрогормонов. Таким образом, совмещая свойства нервных и эндокринных клеток, нейросекреторные клетки объединяют нервные и эндокринные регуляторные механизмы в единую нейроэндокринную систему. Этим обеспечивается, в частности, способность организма адаптироваться к изменяющимся условиям внешней среды. Объединение нервных эндокринных механизмов регуляции осуществляется на уровне гипоталамуса и гипофиза.
Гидроцефалический (интерстициальный) отек. Еще одной формой отека головного мозга, сопровождающейся увеличением объема интерстициального пространства, является гидроцефалический отек, обусловленный блокадой путей, соединяющих интерстициальное пространство головного мозга с макроскопическими ликворосодержащими пространствами. Для клинициста эта форма отека имеет практическое значение. Так, у больных с острой гидроцефалией в начале происходит увеличение объема интерстициальной жидкости в перивентрикулярных отделах. Узкие в норме пространства между глиальными клетками и аксонами расширяются. Астроциты набухают, атрофируются и погибают. У больных с хронической гидроцефалией деструкция аксонов, разрушение миелина, фагоцитоз липидов микроглии являются характерными гистологическими признаками. Кроме стаза внеклеточной жидкости, причиной отека у этих больных может быть и обратный ток спинномозговой жидкости из желудочков мозга. Так же, как и при любой иной форме отека мозга, в зоне отек; снижается регионарный мозговой кровоток. По-видимому, часть функциональных расстройств в ЦНС, наблюдаемых у больных с гидроцефалией, обусловлена снижением регионарного кровотока в зоне отека.
Эти два гормона относятся к особой группе, поскольку, синтезируясь в гипоталамусе, транспортируются по аксонам (отросткам нейронов) в задний гипофиз и там выделяются в системный кровоток.
Между концами может остаться небольшой диастаз, но он не должен превышать 1 мм. Свободный промежуток между концами нерва заполнит гематома, а в дальнейшем образуется соединительно—тканная прослойка, через эту гематому и соединительную ткань будут прорастать тяжи швашювских клеток и вновь сформированные аксоны.