 Функции клеток нервной системы весьма разнообразны. Один из видов − это моторный нейрон (мотонейрон). Его название в переводе с латыни означает «приводящий в движение». Именно при его посредстве и происходит сокращение мышц.
Функции клеток нервной системы весьма разнообразны. Один из видов − это моторный нейрон (мотонейрон). Его название в переводе с латыни означает «приводящий в движение». Именно при его посредстве и происходит сокращение мышц.
Особенность двигательных нервных клеток в том, что их цитоплазма не окружает ядро равномерно, а образует два отростка. Один из них более короткий (дендрит) принимает нервный импульс, второй (аксон) передает его дальше.
Таким образом, двигательный периферический нейрон проводит нервный импульс от центральной нервной системы к мышце. В мышечной ткани его длинный отросток разветвляется и соединяется с десятками мышечных волокон.
Виды двигательных нейронов
По локализации моторные нейроны подразделяются на центральные и периферические. Центральные расположены в ткани головного мозга. Они отвечают за сознательные контролируемые мышечные сокращения.
Двигательные нейроны, идущие непосредственно к мышечным волокнам, называют соматическими.

Тела двигательных нейронов соматической нервной системы расположены в области передних рогов спинного мозга и располагаются группами, каждая из которых отвечает за сокращение строго определенной мускулатуры. К примеру, мотонейроны шейного отдела управляют мускулатурой рук, поясничного отдела отвечают за иннервацию ног.
Периферические нервные клетки, отвечающие за движения, классифицируются следующим образом:
- большие альфа-мотонейроны;
- малые альфа-мотонейроны;
- гамма-мотонейроны;
- клетки Реншо.
Большие альфа-клетки формируют крупные проводящие стволы. Малые альфа и гамма-нейроны имеют более тонкие аксоны. Клетки Реншо входят в состав крупных стволов и служат для коммутации сигналов.

Гамма-мотонейронная петля
Функции мотонейрона
Центральные и периферические двигательные нервные клетки работают согласовано. Совместно они обеспечивают сокращение определенных групп мышц и позволяют человеку выполнять какие-либо действия.
Для координированных движений конечностей необходимо одновременное сокращение сгибателей и разгибателей. При работе сгибателей первоначальный сигнал возбуждения возникает в области прецентральной извилины соответствующего полушария.
За это действие отвечают клетки, называемые пирамидными. Собранные вместе их отростки образуют так называемый пирамидный двигательный путь. Далее сигнал идет к передним рогам спинного мозга, откуда передается уже непосредственно в миофибриллы.
Активирующее влияние на мотонейроны мышц разгибателей оказывают специальные центры задних отделов больших полушарий. Они формируют дорсальный и вентральный пути. Таким образом, в формировании координированного движения участвуют две области головного мозга.

По характеру функции нервные клетки, задействованные в процессе мышечного сокращения, подразделяются на двигательные и вставочные нейроны. Первые ответственны за исполнительную функцию, в то время как для координации нервных импульсов служат вставочные. Эта особая разновидность имеет меньшие размеры и более многочисленна.
Для сравнения — в области передних рогов их в 30 раз больше чем двигательных. Когда возбуждение проводится по аксону двигательного нерва, оно переходит первоначально на вставочный нейрон. В зависимости от характера сигнала он может быть усилен либо ослаблен, после чего передается дальше.
Клетки вставочного типа имеют больше отростков и более чувствительны. Они обладают большим числом отростков и их еще называют мультиполярными.

Для оптимизации сигналов исходящих по аксонам и идущим к мышечным волокнам, служат специальные клетки Реншоу, которые передают возбуждение с одного отростка на другой. Такой механизм служит выравниванию интенсивности нервного сигнала.
По отростку мотонейрона импульс достигает мышечного волокна, которое сокращается. Каждая группа мотонейронов и иннервируемые ими мышечные волокна отвечают за определенные движения.
Нервные клетки, обеспечивающие двигательную функцию:
| Виды нейронов | Локализация | Функция |
|---|---|---|
| центральные иннервирующие сгибатели |
область прецентральной извилины | сокращение скелетных мышц сгибателей путем передачи импульса в область передних рогов |
| центральные иннервирующие разгибатели |
область заднего мозга | сокращение скелетных мышц разгибателей путем передачи импульса в область передних рогов |
| периферические альфа | передние рога спинного мозга | непосредственное сокращение скелетных мышц |
| периферические гамма | передние рога спинного мозга | регуляция тонуса |
| вставочные | все отделы ЦНС | коммуникация сигналов внутри ЦНС |
Большие альфа-нейроны, проводящие сильный импульс, вызывают сокращение миофибрилл. Малые проводят слабые сигналы и служат для поддержания тонуса мускулатуры.
Помимо волокон, отвечающих за сокращение, в мышечной ткани есть и специальные спиральные фибриллы, регулирующие силу напряжения мускулатуры.
Эти экстрафузальные мышечные волокна иннервируются гамма-нейронами.
Возбуждение гамма-мотонейрона приводит к увеличению растяжения миофибрилл и облегчает прохождение импульса сухожильных рефлексов. Примером может быть прохождение нервного сигнала по дуге коленного рефлекса.
Слаженной работой периферических мотонейронов достигается тонкая настройка мышечного тонуса, что позволяет выполнять точные координированные движения. При поражении периферических двигательных нейронов мышечный тонус исчезает и движения невозможны.

Как работает двигательный нейрон?
Для того чтобы возник биоэлектрический импульс, необходима разница потенциалов на оболочке нервной клетки. Это происходит в результате изменения концентрации ионов калия и натрия с наружной и внутренней поверхности мембраны.
В дальнейшем импульс проходит до конца длинного отростка ‒ аксона и достигает места соединения с другой клеткой. Место такого контакта называют синапс.

С другой стороны синапса к месту контакта прилежит короткий ветвящийся отросток ‒ дендрит. Передача сигнала через синапс обусловлена активными химическими веществами, так называемыми медиаторами.
Возникнув на дендрите, сигнал распространяется по его оболочке и переходит далее на аксон. Для сокращения скелетной мышцы сигнал зарождается в мотонейроне коры, проходит по пирамидному пути, переходит на вставочный нейрон и далее в область передних рогов спинного мозга. Заканчивается эта цепь в мышечной ткани.
Результатом возбуждения двигательного центра коры будет сокращение группы мышечных волокон.
Симптомы поражения центрального двигательного нейрона
Поражения центральных моторных нервных клеток возникают чаще всего при инсульте. При ишемии или кровоизлиянии в вещество больших полушарий участок ткани отмирает. Такие поражения почти всегда односторонние.
Вследствие этого при поражении центральных двигательных нейронов наблюдаются нарушения функции мускулатуры с одной стороны. Самый заметный признак ‒ это односторонний паралич, приводящий к невозможности активных движений в руке и ноге.
С этой же стороны снижается мышечный тонус в туловище и мимической мускулатуре лица. Поражение центральных моторных областей сопровождается рядом изменений рефлекторной деятельности.
Клинически это выражается в появлении разнообразных патологических рефлексов. Их совокупность, снижение тонуса мышц и нарушения чувствительности позволяют врачу установить диагноз.
From Wikipedia, the free encyclopedia
| Motor neurons | |
|---|---|

Micrograph of the hypoglossal nucleus showing motor neurons with their characteristic coarse Nissl substance («tigroid» cytoplasm). H&E-LFB stain. |
|
| Details | |
| Location | Ventral horn of the spinal cord, some cranial nerve nuclei |
| Shape | Projection neuron |
| Function | Excitatory projection (to NMJ) |
| Neurotransmitter | UMN to LMN: glutamate; LMN to NMJ: ACh |
| Presynaptic connections | Primary motor cortex via the Corticospinal tract |
| Postsynaptic connections | Muscle fibers and other neurons |
| Identifiers | |
| MeSH | D009046 |
| NeuroLex ID | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A motor neuron (or motoneuron or efferent neuron[1]) is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands.[2] There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons.[3] The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors.[4] Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. Innervation takes place at a neuromuscular junction and twitches can become superimposed as a result of summation or a tetanic contraction. Individual twitches can become indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
Development[edit]
Motor neurons begin to develop early in embryonic development, and motor function continues to develop well into childhood.[6] In the neural tube cells are specified to either the rostral-caudal axis or ventral-dorsal axis. The axons of motor neurons begin to appear in the fourth week of development from the ventral region of the ventral-dorsal axis (the basal plate).[7] This homeodomain is known as the motor neural progenitor domain (pMN). Transcription factors here include Pax6, OLIG2, Nkx-6.1, and Nkx-6.2, which are regulated by sonic hedgehog (Shh). The OLIG2 gene being the most important due to its role in promoting Ngn2 expression, a gene that causes cell cycle exiting as well as promoting further transcription factors associated with motor neuron development.[8]
Further specification of motor neurons occurs when retinoic acid, fibroblast growth factor, Wnts, and TGFb, are integrated into the various Hox transcription factors. There are 13 Hox transcription factors and along with the signals, determine whether a motor neuron will be more rostral or caudal in character. In the spinal column, Hox 4-11 sort motor neurons to one of the five motor columns.[8]
| Motor column | Location in spinal cord | Target |
| Median motor column | Present entire length | Axial muscles |
| Hypaxial motor column | Thoracic region | Body wall muscles |
| Preganglionic motor column | Thoracic region | Sympathetic ganglion |
| Lateral motor column | Brachial and lumbar region (both regions are further divided into medial and lateral domains) | Muscles of the limbs |
| Phrenic motor column | Cervical region | Diaphragm[10] |
Anatomy and physiology[edit]


Location of lower motor neurons in spinal cord
Upper motor neurons[edit]
Upper motor neurons originate in the motor cortex located in the precentral gyrus. The cells that make up the primary motor cortex are Betz cells, which are giant pyramidal cells. The axons of these cells descend from the cortex to form the corticospinal tract.[11] Corticomotorneurons project from the primary cortex directly onto motor neurons in the ventral horn of the spinal cord.[12][13] Their axons synapse on the spinal motor neurons of multiple muscles as well as on spinal interneurons.[12][13] They are unique to primates and it has been suggested that their function is the adaptive control of the hands including the relatively independent control of individual fingers.[13][14] Corticomotorneurons have so far only been found in the primary motor cortex and not in secondary motor areas.[13]
Nerve tracts[edit]
Nerve tracts are bundles of axons as white matter, that carry action potentials to their effectors. In the spinal cord these descending tracts carry impulses from different regions. These tracts also serve as the place of origin for lower motor neurons. There are seven major descending motor tracts to be found in the spinal cord:[15]
- Lateral corticospinal tract
- Rubrospinal tract
- Lateral reticulospinal tract
- Vestibulospinal tract
- Medial reticulospinal tract
- Tectospinal tract
- Anterior corticospinal tract
Lower motor neurons[edit]
Lower motor neurons are those that originate in the spinal cord and directly or indirectly innervate effector targets. The target of these neurons varies, but in the somatic nervous system the target will be some sort of muscle fiber. There are three primary categories of lower motor neurons, which can be further divided in sub-categories.[16]
According to their targets, motor neurons are classified into three broad categories:[17]
- Somatic motor neurons
- Special visceral motor neurons
- General visceral motor neurons
Somatic motor neurons[edit]
Somatic motor neurons originate in the central nervous system, project their axons to skeletal muscles[18] (such as the muscles of the limbs, abdominal, and intercostal muscles), which are involved in locomotion. The three types of these neurons are the alpha efferent neurons, beta efferent neurons, and gamma efferent neurons. They are called efferent to indicate the flow of information from the central nervous system (CNS) to the periphery.
- Alpha motor neurons innervate extrafusal muscle fibers, which are the main force-generating component of a muscle. Their cell bodies are in the ventral horn of the spinal cord and they are sometimes called ventral horn cells. A single motor neuron may synapse with 150 muscle fibers on average.[19] The motor neuron and all of the muscle fibers to which it connects is a motor unit. Motor units are split up into 3 categories:[20]
- Slow (S) motor units stimulate small muscle fibers, which contract very slowly and provide small amounts of energy but are very resistant to fatigue, so they are used to sustain muscular contraction, such as keeping the body upright. They gain their energy via oxidative means and hence require oxygen. They are also called red fibers.[20]
- Fast fatiguing (FF) motor units stimulate larger muscle groups, which apply large amounts of force but fatigue very quickly. They are used for tasks that require large brief bursts of energy, such as jumping or running. They gain their energy via glycolytic means and hence don’t require oxygen. They are called white fibers.[20]
- Fast fatigue-resistant motor units stimulate moderate-sized muscles groups that don’t react as fast as the FF motor units, but can be sustained much longer (as implied by the name) and provide more force than S motor units. These use both oxidative and glycolytic means to gain energy.[20]
In addition to voluntary skeletal muscle contraction, alpha motor neurons also contribute to muscle tone, the continuous force generated by noncontracting muscle to oppose stretching. When a muscle is stretched, sensory neurons within the muscle spindle detect the degree of stretch and send a signal to the CNS. The CNS activates alpha motor neurons in the spinal cord, which cause extrafusal muscle fibers to contract and thereby resist further stretching. This process is also called the stretch reflex.
- Beta motor neurons innervate intrafusal muscle fibers of muscle spindles, with collaterals to extrafusal fibres. There are two types of beta motor neurons: Slow Contracting- These innervate extrafusal fibers. Fast Contracting- These innervate intrafusal fibers.[21]
- Gamma motor neurons innervate intrafusal muscle fibers found within the muscle spindle. They regulate the sensitivity of the spindle to muscle stretching. With activation of gamma neurons, intrafusal muscle fibers contract so that only a small stretch is required to activate spindle sensory neurons and the stretch reflex. There are two types of gamma motor neurons: Dynamic- These focus on Bag1 fibers and enhance dynamic sensitivity. Static- These focus on Bag2 fibers and enhance stretch sensitivity.[21]
- Regulatory factors of lower motor neurons
- Size Principle – this relates to the soma of the motor neuron. This restricts larger neurons to receive a larger excitatory signal in order to stimulate the muscle fibers it innervates. By reducing unnecessary muscle fiber recruitment, the body is able to optimize energy consumption.[21]
- Persistent Inward Current (PIC) – recent animal study research has shown that constant flow of ions such as calcium and sodium through channels in the soma and dendrites influence the synaptic input. An alternate way to think of this is that the post-synaptic neuron is being primed before receiving an impulse.[21]
- After Hyper-polarization (AHP) – A trend has been identified that shows slow motor neurons to have more intense AHPs for a longer duration. One way to remember this is that slow muscle fibers can contract for longer, so it makes sense that their corresponding motor neurons fire at a slower rate.[21]
Special visceral motor neurons[edit]
These are also known as branchial motor neurons, which are involved in facial expression, mastication, phonation, and swallowing. Associated cranial nerves are the oculomotor, abducens, trochlear, and hypoglossal nerves.[17]
| Branch of NS | Position | Neurotransmitter |
|---|---|---|
| Somatic | n/a | Acetylcholine |
| Parasympathetic | Preganglionic | Acetylcholine |
| Parasympathetic | Ganglionic | Acetylcholine |
| Sympathetic | Preganglionic | Acetylcholine |
| Sympathetic | Ganglionic | Norepinephrine* |
| *Except fibers to sweat glands and certain blood vessels Motor neuron neurotransmitters |
General visceral motor neurons[edit]
These motor neurons indirectly innervate cardiac muscle and smooth muscles of the viscera ( the muscles of the arteries): they synapse onto neurons located in ganglia of the autonomic nervous system (sympathetic and parasympathetic), located in the peripheral nervous system (PNS), which themselves directly innervate visceral muscles (and also some gland cells).
In consequence, the motor command of skeletal and branchial muscles is monosynaptic involving only one motor neuron, either somatic or branchial, which synapses onto the muscle. Comparatively, the command of visceral muscles is disynaptic involving two neurons: the general visceral motor neuron, located in the CNS, synapses onto a ganglionic neuron, located in the PNS, which synapses onto the muscle.
All vertebrate motor neurons are cholinergic, that is, they release the neurotransmitter acetylcholine. Parasympathetic ganglionic neurons are also cholinergic, whereas most sympathetic ganglionic neurons are noradrenergic, that is, they release the neurotransmitter noradrenaline. (see Table)
Neuromuscular junctions[edit]
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. As a result, if an action potential arrives before a twitch has completed, the twitches can superimpose on one another, either through summation or a tetanic contraction. In summation, the muscle is stimulated repetitively such that additional action potentials coming from the somatic nervous system arrive before the end of the twitch. The twitches thus superimpose on one another, leading to a force greater than that of a single twitch. A tetanic contraction is caused by constant, very high frequency stimulation — the action potentials come at such a rapid rate that individual twitches are indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
The interface between a motor neuron and muscle fiber is a specialized synapse called the neuromuscular junction. Upon adequate stimulation, the motor neuron releases a flood of acetylcholine (Ach) neurotransmitters from the axon terminals from synaptic vesicles bind with the plasma membrane. The acetylcholine molecules bind to postsynaptic receptors found within the motor end plate. Once two acetylcholine receptors have been bound, an ion channel is opened and sodium ions are allowed to flow into the cell. The influx of sodium into the cell causes depolarization and triggers a muscle action potential. T tubules of the sarcolemma are then stimulated to elicit calcium ion release from the sarcoplasmic reticulum. It is this chemical release that causes the target muscle fiber to contract.[19]
In invertebrates, depending on the neurotransmitter released and the type of receptor it binds, the response in the muscle fiber could be either excitatory or inhibitory. For vertebrates, however, the response of a muscle fiber to a neurotransmitter can only be excitatory, in other words, contractile. Muscle relaxation and inhibition of muscle contraction in vertebrates is obtained only by inhibition of the motor neuron itself. This is how muscle relaxants work by acting on the motor neurons that innervate muscles (by decreasing their electrophysiological activity) or on cholinergic neuromuscular junctions, rather than on the muscles themselves.
See also[edit]
- Betz cell
- Central chromatolysis
- Motor dysfunction
- Motor neuron disease
- Nerve
- Efferent nerve fiber
- Motor nerve
References[edit]
- ^ «Afferent vs. Efferent: AP® Psych Crash Course Review | Albert.io». Albert Resources. 2019-12-02. Retrieved 2021-04-25.
- ^ Tortora, Gerard; Derrickson, Bryan (2014). Principles of Anatomy & Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 406, 502, 541. ISBN 978-1-118-34500-9.
- ^ Pocock, Gillian; Richards, Christopher D. (2006). Human physiology : the basis of medicine (3rd ed.). Oxford: Oxford University Press. pp. 151–153. ISBN 978-0-19-856878-0.
- ^ Schacter D.L., Gilbert D.T., and Wegner D.M. (2011) Psychology second edition. New York, NY: Worth
- ^ a b Russell, Peter (2013). Biology — Exploring the Diversity of Life. Toronto: Nelson Education. p. 946. ISBN 978-0-17-665133-6.
- ^ Tortora, Gerard; Derrickson, Bryan (2011). Principles of Anatomy Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 1090–1099. ISBN 978-1-118-34500-9.
- ^ Sadler, T. (2010). Langman’s medical embryology (11th ed.). Philadelphia: Lippincott William & Wilkins. pp. 299–301. ISBN 978-0-7817-9069-7.
- ^ a b Davis-Dusenbery, BN; Williams, LA; Klim, JR; Eggan, K (February 2014). «How to make spinal motor neurons». Development. 141 (3): 491–501. doi:10.1242/dev.097410. PMID 24449832.
- ^ Edgar R, Mazor Y, Rinon A, Blumenthal J, Golan Y, Buzhor E, Livnat I, Ben-Ari S, Lieder I, Shitrit A, Gilboa Y, Ben-Yehudah A, Edri O, Shraga N, Bogoch Y, Leshansky L, Aharoni S, West MD, Warshawsky D, Shtrichman R (2013). «LifeMap Discovery™: The Embryonic Development, Stem Cells, and Regenerative Medicine Research Portal». PLOS ONE. 8 (7): e66629. Bibcode:2013PLoSO…866629E. doi:10.1371/journal.pone.0066629. ISSN 1932-6203. PMC 3714290. PMID 23874394.
- ^ Philippidou, Polyxeni; Walsh, Carolyn; Aubin, Josée; Jeannotte, Lucie; Dasen, Jeremy S. (2012). «Sustained Hox5 Gene Activity is Required for Respiratory Motor Neuron Development». Nature Neuroscience. 15 (12): 1636–1644. doi:10.1038/nn.3242. ISSN 1097-6256. PMC 3676175. PMID 23103965.
- ^ Fitzpatrick, D. (2001) The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b Mack, Sarah; Kandel, Eric R.; Jessell, Thomas M.; Schwartz, James H.; Siegelbaum, Steven A.; Hudspeth, A. J. (2013). Principles of neural science. Kandel, Eric R. (5th ed.). New York. ISBN 9780071390118. OCLC 795553723.
- ^ a b c d Lemon, Roger N. (April 4, 2008). «Descending Pathways in Motor Control». Annual Review of Neuroscience. 31 (1): 195–218. doi:10.1146/annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853. S2CID 16139768.
- ^ Isa, T (April 2007). «Direct and indirect cortico-motoneuronal pathways and control of hand/arm movements». Physiology. 22 (2): 145–152. doi:10.1152/physiol.00045.2006. PMID 17420305.
- ^ Tortora, G. J., Derrickson, B. (2011). The Spinal Cord and Spinal Nerves. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 443-468). New Jersey: John Wiley & Sons, Inc.
- ^ Fitzpatrick, D. (2001) Lower Motor Neuron Circuits and Motor Control: Overview. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «Lower Motor Neuron Circuits and Motor Control — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b «CHAPTER NINE». www.unc.edu. Archived from the original on 2017-11-05. Retrieved 2017-12-08.
- ^ Silverthorn, Dee Unglaub (2010). Human Physiology: An Integrated Approach. Pearson. p. 398. ISBN 978-0-321-55980-7.
- ^ a b Tortora, G. J., Derrickson, B. (2011). Muscular Tissue. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 305-307, 311). New Jersey: John Wiley & Sons, Inc.
- ^ a b c d Purves D, Augustine GJ, Fitzpatrick D, et al., editors: Neuroscience. 2nd edition, 2001 «The Motor Unit — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-09-05.
- ^ a b c d e Manuel, Marin; Zytnicki, Daniel (2011). «Alpha, Beta, and Gamma Motoneurons: Functional Diversity in the Motor System’s Final Pathway». Journal of Integrative Neuroscience. 10 (3): 243–276. doi:10.1142/S0219635211002786. ISSN 0219-6352. PMID 21960303. S2CID 21582283.
Sources[edit]
- Sherwood, L. (2001). Human Physiology: From Cells to Systems (4th ed.). Pacific Grove, CA: Brooks-Cole. ISBN 0-534-37254-6.
- Marieb, E. N.; Mallatt, J. (1997). Human Anatomy (2nd ed.). Menlo Park, CA: Benjamin/Cummings. ISBN 0-8053-4068-8.
From Wikipedia, the free encyclopedia
| Motor neurons | |
|---|---|
|
Micrograph of the hypoglossal nucleus showing motor neurons with their characteristic coarse Nissl substance («tigroid» cytoplasm). H&E-LFB stain. |
|
| Details | |
| Location | Ventral horn of the spinal cord, some cranial nerve nuclei |
| Shape | Projection neuron |
| Function | Excitatory projection (to NMJ) |
| Neurotransmitter | UMN to LMN: glutamate; LMN to NMJ: ACh |
| Presynaptic connections | Primary motor cortex via the Corticospinal tract |
| Postsynaptic connections | Muscle fibers and other neurons |
| Identifiers | |
| MeSH | D009046 |
| NeuroLex ID | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A motor neuron (or motoneuron or efferent neuron[1]) is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands.[2] There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons.[3] The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors.[4] Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. Innervation takes place at a neuromuscular junction and twitches can become superimposed as a result of summation or a tetanic contraction. Individual twitches can become indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
Development[edit]
Motor neurons begin to develop early in embryonic development, and motor function continues to develop well into childhood.[6] In the neural tube cells are specified to either the rostral-caudal axis or ventral-dorsal axis. The axons of motor neurons begin to appear in the fourth week of development from the ventral region of the ventral-dorsal axis (the basal plate).[7] This homeodomain is known as the motor neural progenitor domain (pMN). Transcription factors here include Pax6, OLIG2, Nkx-6.1, and Nkx-6.2, which are regulated by sonic hedgehog (Shh). The OLIG2 gene being the most important due to its role in promoting Ngn2 expression, a gene that causes cell cycle exiting as well as promoting further transcription factors associated with motor neuron development.[8]
Further specification of motor neurons occurs when retinoic acid, fibroblast growth factor, Wnts, and TGFb, are integrated into the various Hox transcription factors. There are 13 Hox transcription factors and along with the signals, determine whether a motor neuron will be more rostral or caudal in character. In the spinal column, Hox 4-11 sort motor neurons to one of the five motor columns.[8]
| Motor column | Location in spinal cord | Target |
| Median motor column | Present entire length | Axial muscles |
| Hypaxial motor column | Thoracic region | Body wall muscles |
| Preganglionic motor column | Thoracic region | Sympathetic ganglion |
| Lateral motor column | Brachial and lumbar region (both regions are further divided into medial and lateral domains) | Muscles of the limbs |
| Phrenic motor column | Cervical region | Diaphragm[10] |
Anatomy and physiology[edit]
Location of lower motor neurons in spinal cord
Upper motor neurons[edit]
Upper motor neurons originate in the motor cortex located in the precentral gyrus. The cells that make up the primary motor cortex are Betz cells, which are giant pyramidal cells. The axons of these cells descend from the cortex to form the corticospinal tract.[11] Corticomotorneurons project from the primary cortex directly onto motor neurons in the ventral horn of the spinal cord.[12][13] Their axons synapse on the spinal motor neurons of multiple muscles as well as on spinal interneurons.[12][13] They are unique to primates and it has been suggested that their function is the adaptive control of the hands including the relatively independent control of individual fingers.[13][14] Corticomotorneurons have so far only been found in the primary motor cortex and not in secondary motor areas.[13]
Nerve tracts[edit]
Nerve tracts are bundles of axons as white matter, that carry action potentials to their effectors. In the spinal cord these descending tracts carry impulses from different regions. These tracts also serve as the place of origin for lower motor neurons. There are seven major descending motor tracts to be found in the spinal cord:[15]
- Lateral corticospinal tract
- Rubrospinal tract
- Lateral reticulospinal tract
- Vestibulospinal tract
- Medial reticulospinal tract
- Tectospinal tract
- Anterior corticospinal tract
Lower motor neurons[edit]
Lower motor neurons are those that originate in the spinal cord and directly or indirectly innervate effector targets. The target of these neurons varies, but in the somatic nervous system the target will be some sort of muscle fiber. There are three primary categories of lower motor neurons, which can be further divided in sub-categories.[16]
According to their targets, motor neurons are classified into three broad categories:[17]
- Somatic motor neurons
- Special visceral motor neurons
- General visceral motor neurons
Somatic motor neurons[edit]
Somatic motor neurons originate in the central nervous system, project their axons to skeletal muscles[18] (such as the muscles of the limbs, abdominal, and intercostal muscles), which are involved in locomotion. The three types of these neurons are the alpha efferent neurons, beta efferent neurons, and gamma efferent neurons. They are called efferent to indicate the flow of information from the central nervous system (CNS) to the periphery.
- Alpha motor neurons innervate extrafusal muscle fibers, which are the main force-generating component of a muscle. Their cell bodies are in the ventral horn of the spinal cord and they are sometimes called ventral horn cells. A single motor neuron may synapse with 150 muscle fibers on average.[19] The motor neuron and all of the muscle fibers to which it connects is a motor unit. Motor units are split up into 3 categories:[20]
- Slow (S) motor units stimulate small muscle fibers, which contract very slowly and provide small amounts of energy but are very resistant to fatigue, so they are used to sustain muscular contraction, such as keeping the body upright. They gain their energy via oxidative means and hence require oxygen. They are also called red fibers.[20]
- Fast fatiguing (FF) motor units stimulate larger muscle groups, which apply large amounts of force but fatigue very quickly. They are used for tasks that require large brief bursts of energy, such as jumping or running. They gain their energy via glycolytic means and hence don’t require oxygen. They are called white fibers.[20]
- Fast fatigue-resistant motor units stimulate moderate-sized muscles groups that don’t react as fast as the FF motor units, but can be sustained much longer (as implied by the name) and provide more force than S motor units. These use both oxidative and glycolytic means to gain energy.[20]
In addition to voluntary skeletal muscle contraction, alpha motor neurons also contribute to muscle tone, the continuous force generated by noncontracting muscle to oppose stretching. When a muscle is stretched, sensory neurons within the muscle spindle detect the degree of stretch and send a signal to the CNS. The CNS activates alpha motor neurons in the spinal cord, which cause extrafusal muscle fibers to contract and thereby resist further stretching. This process is also called the stretch reflex.
- Beta motor neurons innervate intrafusal muscle fibers of muscle spindles, with collaterals to extrafusal fibres. There are two types of beta motor neurons: Slow Contracting- These innervate extrafusal fibers. Fast Contracting- These innervate intrafusal fibers.[21]
- Gamma motor neurons innervate intrafusal muscle fibers found within the muscle spindle. They regulate the sensitivity of the spindle to muscle stretching. With activation of gamma neurons, intrafusal muscle fibers contract so that only a small stretch is required to activate spindle sensory neurons and the stretch reflex. There are two types of gamma motor neurons: Dynamic- These focus on Bag1 fibers and enhance dynamic sensitivity. Static- These focus on Bag2 fibers and enhance stretch sensitivity.[21]
- Regulatory factors of lower motor neurons
- Size Principle – this relates to the soma of the motor neuron. This restricts larger neurons to receive a larger excitatory signal in order to stimulate the muscle fibers it innervates. By reducing unnecessary muscle fiber recruitment, the body is able to optimize energy consumption.[21]
- Persistent Inward Current (PIC) – recent animal study research has shown that constant flow of ions such as calcium and sodium through channels in the soma and dendrites influence the synaptic input. An alternate way to think of this is that the post-synaptic neuron is being primed before receiving an impulse.[21]
- After Hyper-polarization (AHP) – A trend has been identified that shows slow motor neurons to have more intense AHPs for a longer duration. One way to remember this is that slow muscle fibers can contract for longer, so it makes sense that their corresponding motor neurons fire at a slower rate.[21]
Special visceral motor neurons[edit]
These are also known as branchial motor neurons, which are involved in facial expression, mastication, phonation, and swallowing. Associated cranial nerves are the oculomotor, abducens, trochlear, and hypoglossal nerves.[17]
| Branch of NS | Position | Neurotransmitter |
|---|---|---|
| Somatic | n/a | Acetylcholine |
| Parasympathetic | Preganglionic | Acetylcholine |
| Parasympathetic | Ganglionic | Acetylcholine |
| Sympathetic | Preganglionic | Acetylcholine |
| Sympathetic | Ganglionic | Norepinephrine* |
| *Except fibers to sweat glands and certain blood vessels Motor neuron neurotransmitters |
General visceral motor neurons[edit]
These motor neurons indirectly innervate cardiac muscle and smooth muscles of the viscera ( the muscles of the arteries): they synapse onto neurons located in ganglia of the autonomic nervous system (sympathetic and parasympathetic), located in the peripheral nervous system (PNS), which themselves directly innervate visceral muscles (and also some gland cells).
In consequence, the motor command of skeletal and branchial muscles is monosynaptic involving only one motor neuron, either somatic or branchial, which synapses onto the muscle. Comparatively, the command of visceral muscles is disynaptic involving two neurons: the general visceral motor neuron, located in the CNS, synapses onto a ganglionic neuron, located in the PNS, which synapses onto the muscle.
All vertebrate motor neurons are cholinergic, that is, they release the neurotransmitter acetylcholine. Parasympathetic ganglionic neurons are also cholinergic, whereas most sympathetic ganglionic neurons are noradrenergic, that is, they release the neurotransmitter noradrenaline. (see Table)
Neuromuscular junctions[edit]
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. As a result, if an action potential arrives before a twitch has completed, the twitches can superimpose on one another, either through summation or a tetanic contraction. In summation, the muscle is stimulated repetitively such that additional action potentials coming from the somatic nervous system arrive before the end of the twitch. The twitches thus superimpose on one another, leading to a force greater than that of a single twitch. A tetanic contraction is caused by constant, very high frequency stimulation — the action potentials come at such a rapid rate that individual twitches are indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
The interface between a motor neuron and muscle fiber is a specialized synapse called the neuromuscular junction. Upon adequate stimulation, the motor neuron releases a flood of acetylcholine (Ach) neurotransmitters from the axon terminals from synaptic vesicles bind with the plasma membrane. The acetylcholine molecules bind to postsynaptic receptors found within the motor end plate. Once two acetylcholine receptors have been bound, an ion channel is opened and sodium ions are allowed to flow into the cell. The influx of sodium into the cell causes depolarization and triggers a muscle action potential. T tubules of the sarcolemma are then stimulated to elicit calcium ion release from the sarcoplasmic reticulum. It is this chemical release that causes the target muscle fiber to contract.[19]
In invertebrates, depending on the neurotransmitter released and the type of receptor it binds, the response in the muscle fiber could be either excitatory or inhibitory. For vertebrates, however, the response of a muscle fiber to a neurotransmitter can only be excitatory, in other words, contractile. Muscle relaxation and inhibition of muscle contraction in vertebrates is obtained only by inhibition of the motor neuron itself. This is how muscle relaxants work by acting on the motor neurons that innervate muscles (by decreasing their electrophysiological activity) or on cholinergic neuromuscular junctions, rather than on the muscles themselves.
See also[edit]
- Betz cell
- Central chromatolysis
- Motor dysfunction
- Motor neuron disease
- Nerve
- Efferent nerve fiber
- Motor nerve
References[edit]
- ^ «Afferent vs. Efferent: AP® Psych Crash Course Review | Albert.io». Albert Resources. 2019-12-02. Retrieved 2021-04-25.
- ^ Tortora, Gerard; Derrickson, Bryan (2014). Principles of Anatomy & Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 406, 502, 541. ISBN 978-1-118-34500-9.
- ^ Pocock, Gillian; Richards, Christopher D. (2006). Human physiology : the basis of medicine (3rd ed.). Oxford: Oxford University Press. pp. 151–153. ISBN 978-0-19-856878-0.
- ^ Schacter D.L., Gilbert D.T., and Wegner D.M. (2011) Psychology second edition. New York, NY: Worth
- ^ a b Russell, Peter (2013). Biology — Exploring the Diversity of Life. Toronto: Nelson Education. p. 946. ISBN 978-0-17-665133-6.
- ^ Tortora, Gerard; Derrickson, Bryan (2011). Principles of Anatomy Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 1090–1099. ISBN 978-1-118-34500-9.
- ^ Sadler, T. (2010). Langman’s medical embryology (11th ed.). Philadelphia: Lippincott William & Wilkins. pp. 299–301. ISBN 978-0-7817-9069-7.
- ^ a b Davis-Dusenbery, BN; Williams, LA; Klim, JR; Eggan, K (February 2014). «How to make spinal motor neurons». Development. 141 (3): 491–501. doi:10.1242/dev.097410. PMID 24449832.
- ^ Edgar R, Mazor Y, Rinon A, Blumenthal J, Golan Y, Buzhor E, Livnat I, Ben-Ari S, Lieder I, Shitrit A, Gilboa Y, Ben-Yehudah A, Edri O, Shraga N, Bogoch Y, Leshansky L, Aharoni S, West MD, Warshawsky D, Shtrichman R (2013). «LifeMap Discovery™: The Embryonic Development, Stem Cells, and Regenerative Medicine Research Portal». PLOS ONE. 8 (7): e66629. Bibcode:2013PLoSO…866629E. doi:10.1371/journal.pone.0066629. ISSN 1932-6203. PMC 3714290. PMID 23874394.
- ^ Philippidou, Polyxeni; Walsh, Carolyn; Aubin, Josée; Jeannotte, Lucie; Dasen, Jeremy S. (2012). «Sustained Hox5 Gene Activity is Required for Respiratory Motor Neuron Development». Nature Neuroscience. 15 (12): 1636–1644. doi:10.1038/nn.3242. ISSN 1097-6256. PMC 3676175. PMID 23103965.
- ^ Fitzpatrick, D. (2001) The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b Mack, Sarah; Kandel, Eric R.; Jessell, Thomas M.; Schwartz, James H.; Siegelbaum, Steven A.; Hudspeth, A. J. (2013). Principles of neural science. Kandel, Eric R. (5th ed.). New York. ISBN 9780071390118. OCLC 795553723.
- ^ a b c d Lemon, Roger N. (April 4, 2008). «Descending Pathways in Motor Control». Annual Review of Neuroscience. 31 (1): 195–218. doi:10.1146/annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853. S2CID 16139768.
- ^ Isa, T (April 2007). «Direct and indirect cortico-motoneuronal pathways and control of hand/arm movements». Physiology. 22 (2): 145–152. doi:10.1152/physiol.00045.2006. PMID 17420305.
- ^ Tortora, G. J., Derrickson, B. (2011). The Spinal Cord and Spinal Nerves. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 443-468). New Jersey: John Wiley & Sons, Inc.
- ^ Fitzpatrick, D. (2001) Lower Motor Neuron Circuits and Motor Control: Overview. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «Lower Motor Neuron Circuits and Motor Control — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b «CHAPTER NINE». www.unc.edu. Archived from the original on 2017-11-05. Retrieved 2017-12-08.
- ^ Silverthorn, Dee Unglaub (2010). Human Physiology: An Integrated Approach. Pearson. p. 398. ISBN 978-0-321-55980-7.
- ^ a b Tortora, G. J., Derrickson, B. (2011). Muscular Tissue. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 305-307, 311). New Jersey: John Wiley & Sons, Inc.
- ^ a b c d Purves D, Augustine GJ, Fitzpatrick D, et al., editors: Neuroscience. 2nd edition, 2001 «The Motor Unit — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-09-05.
- ^ a b c d e Manuel, Marin; Zytnicki, Daniel (2011). «Alpha, Beta, and Gamma Motoneurons: Functional Diversity in the Motor System’s Final Pathway». Journal of Integrative Neuroscience. 10 (3): 243–276. doi:10.1142/S0219635211002786. ISSN 0219-6352. PMID 21960303. S2CID 21582283.
Sources[edit]
- Sherwood, L. (2001). Human Physiology: From Cells to Systems (4th ed.). Pacific Grove, CA: Brooks-Cole. ISBN 0-534-37254-6.
- Marieb, E. N.; Mallatt, J. (1997). Human Anatomy (2nd ed.). Menlo Park, CA: Benjamin/Cummings. ISBN 0-8053-4068-8.
Рассмотрена иннервация скелетной мышцы. Описан состав и строение нейрона, а также функции нейрона. Рассмотрены функции двигательных, чувствительных и вегетативных нейронов. Описан состав периферического нерва. Введено понятие двигательной единицы и концевой пластинки (синапса).
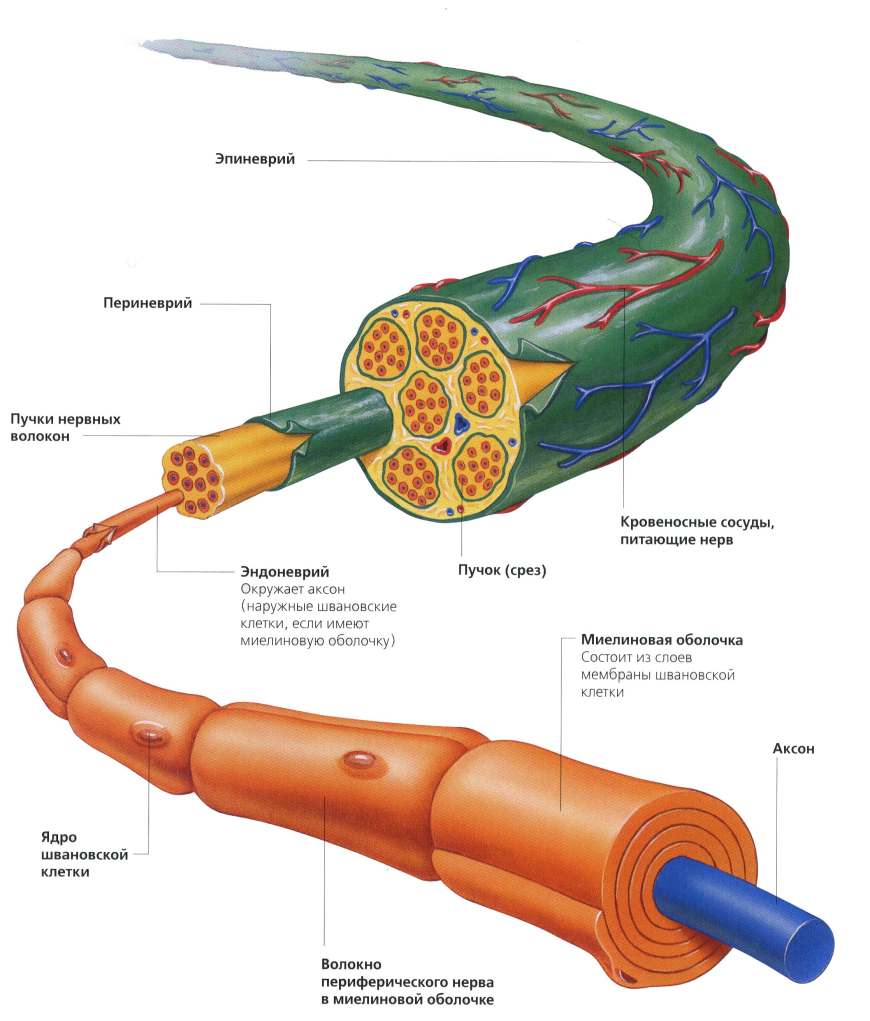
Иннервация скелетной мышцы
Определения
Иннервация (от in — в, внутри и nervus — нервы) — снабжение органов и тканей нервами, что обеспечивает их связь с центральной нервной системой (ЦНС).
Иннервация скелетной мышцы — это наличие нервных волокон, которые передают импульсы из ЦНС к мышце и от мышцы в ЦНС.
Денервация скелетной мышцы — нарушение передачи импульсов из ЦНС к мышце или от мышцы в ЦНС. Денервация мышечных волокон возможна из-за перерезки или повреждения нерва. Денервация мышечных волокон происходит также по мере старения из-за уменьшения количества нервных волокон, иннервирующих скелетную мышцу.
Теперь рассмотрим этот вопрос подробнее. Давайте сначала разберемся, что представляет собой нейрон.
Нейрон
Нейрон – это структурная единица нашей нервной системы, главная функция которого – передача информации от одного участка тела другому. Чтобы передать информацию нейрон возбуждается, затем вырабатывает нервный импульс. Нейроны также участвуют в обработке и хранении информации. Но этого вопроса мы касаться не будем.
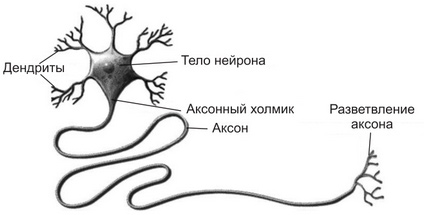
Нейрон – это высокоспециализированная клетка. Он состоит из тела и длинного отростка – аксона (рис. 1.).
Длина аксона может достигать одного метра и более. На теле есть много мелких отростков – дендритов. Через эти отростки нейрон получает информацию от других нейронов и передает ее через аксон или другому нейрону, или органу, к которому он подходит (в нашем случае – мышце).
Периферический нерв
Скелетные мышцы иннервируются одним или несколькими периферическими нервами. За несколькими исключениями (лицевой и подъязычный нерв) все периферические нервы являются смешанными. Смешанный периферический нерв в своем составе содержит:
- двигательные волокна (нейроны);
- чувствительные волокна (нейроны);
- вегетативные волокна (нейроны).
Когда мы рассматривали строение скелетной мышцы, то указывалось, что периферические нервы имеют каналы в перимизии. Через эти каналы периферические нервы подходят к мышечным волокнам.
Двигательные волокна (нейроны)
Двигательные нейроны (мотонейроны) несут информацию от ЦНС к мышце. Тела мотонейронов расположены в передних рогах спинного мозга. Аксоны мотонейронов идут к мышцам в составе периферического нерва.
Двигательные нейроны делятся на два типа: α-мотонейроны (альфа-мотонейроны) и γ-мотонейроны (гамма-мотонейроны).
- α-мотонейроны иннервируют мышечные волокна. Через α-мотонейроны к мышечному волокну поступают импульсы из центральной нервной системы (ЦНС) в результате которых мышечное волокно развивает возбуждение (сокращается).
- γ-мотонейроны иннервируют мышечные веретена (рецепторы мышц). Эти мотонейроны иннервируют особые мышечные волокна (интрафузальные), расположенные внутри мышечных веретен. Напряжение и расслабление интрафузальных волокон изменяет чувствительность рецептора — мышечного веретена. Благодаря этому происходит более «тонкое» управление движениями. За открытие этого механизма шведский нейрофизиолог Рагнар Гранит был удостоен Нобелевской премии.
Один α-мотонейрон иннервирует (то есть пускает веточки) достаточно много мышечных волокон. Иногда на один аксон приходится более 2000 мышечных волокон. Такая система, состоящая из одного нейрона и мышечных волокон, которые он иннервирует, называется двигательной единицей (ДЕ). Это понятие ввел в физиологию нобелевский лауреат Чарльз Скотт Шеррингтон в начале XX века. Особенности состава и функционирования ДЕ мы рассмотрим позднее.
Место, где аксон α-мотонейрона соединяется с мышечным волокном называется концевой пластинкой (синапсом). Через синапс к мышечному волокну из ЦНС (центральной нервной системы) поступают сигналы, которые вызывают его возбуждение.
Чувствительные волокна (нейроны)
Чувствительные волокна несут в ЦНС информацию о различных показателях активности мышцы (длине мышцы, скорости ее сокращения, степени напряжения). Если бы ЦНС не могла получать эту информацию, управление напряжением мышцы было бы невозможно. Точно так же было бы невозможно управление нашими движениями. Тела чувствительных нейронов расположены в задних рогах спинного мозга.
Чувствительные нейроны либо заканчиваются свободными нервными окончаниями, либо иннервируют рецепторы скелетных мышц (мышечные веретена и рецепторы сухожилий).
- Свободные нервные окончания называются ноцицепторами. Они расположены между мышечными волокнами и несут в ЦНС информацию о боли.
- От мышечных веретен отходят чувствительные нейроны (Ia-афференты), несущие в ЦНС информацию о длине и скорости сокращения скелетной мышцы.
- От сухожильных рецепторов отходят чувствительные нейроны (Ib-афференты), несущие в ЦНС информацию о напряжении скелетной мышцы.
Вегетативные волокна (нейроны)
Вегетативные нейроны иннервируют гладкие мышцы стенок кровеносных сосудов скелетных мышц.
Влияние старения на иннервацию скелетных мышц
Иннервация скелетных мышц пожилых людей ухудшается. Ученые находят, что с возрастом уменьшается количество α-мотонейронов, иннервирующих скелетную мышцу. Это является одним из факторов, обусловливающих возрастное уменьшение массы скелетных мышц и их силы — саркопению.
Литература
- Ванек, Ю. Спортивная анатомия.– М.: Академия, 2008.- 304 с.
- Мак-Комас, А.Дж. Скелетные мышцы.– Киев: Олимпийская литература, 2001.– 407 с.
- Мышцы в спорте. Анатомия. Физиология. Тренировка. Реабилитация. — М.: Практическая медицина, 2016.- 408 с. (отличная книга, рекомендую!).
- Самсонова, А.В. Гипертрофия скелетных мышц человека. – СПб: Кинетика, 2018. – 159 с.
С уважением, А.В. Самсонова
Похожие записи:
Типы конституции женщин по И.Б. Галанту
Описана биография известного советского психиатра И.Б.Галанта, предложившего естественную систему конституциональных типов женщин. Дана характеристика предложенных И.Б.Галантом конституциональных…
Сила тяжести
Дано определение силы тяжести. Показано, что сила тяжести является частным случаем силы гравитации. Описаны факторы, определяющие силу тяжести:…
Сила
Дано определение силы в механике. Описаны факторы, определяющие действие на тело силы: направление, точка приложения и численное значение.
Метаболический стресс. Накопление лактата в мышцах
Описан механизм влияния метаболического стресса (накопления лактата) на гипертрофию мышечных волокон. Показано, что накопление лактата приводит…
Механическое повреждение мышечных волокон
Описаны механизмы механического повреждения мышечных волокон при силовой тренировке, приводящие к гипертрофии скелетных мышц. Показано, что…
Механическое напряжение (механотрансдукция) в скелетных мышцах
Описаны процессы передачи механического напряжения в скелетных мышцах. Показано, что механическое напряжение, возникающее вследствие сокращения скелетных…
Влияние возраста и тренировки на результаты в спринтерском беге, структуру и сократительные свойства мышц спортсменов
Изучалось влияние возраста и тренировки на результаты в спринтерском беге, структуру и сократительные свойства мышц спортсменов. Результаты…
Снижение массы скелетных мышц при старении в основном объясняется уменьшением размера мышечных волокон II типа
Снижение массы скелетных мышц при старении в основном связано с меньшим размером мышечных волокон II типа.
ДГЭА (дегидроэпиандростерон, DHEA)
Дано определение, характеристика изменения концентрации ДГЭА с возрастом. Описаны функции ДГЭА и ДГЭА (С) у молодых и пожилых…