Биология,
вопрос задал pusakova548,
4 года назад
На терминалах аксона расположены специальные приспособления для передачи сигналов – химические …
Ответы на вопрос
Ответил 024857
0
Ответ:
скорее всего это синапсы
Объяснение:
Предыдущий вопрос
Следующий вопрос
Новые вопросы
Биология,
7 месяцев назад
ПОМОГИТЕ, ПОЖАЛУЙСТА, СРОЧНО НУЖНО!
Какие способы размножения характерны для растений?
Какое размножение называют половым?
Что такое зигота?
География,
7 месяцев назад
большой водораздельный хребет: 1. Строение земной коры( что-то связанное с кристалическим фундаментом или осадочным чехлом) 2. Климат( средняя температура в январе и летом) 3. Внутренние воды 4.
Химия,
4 года назад
Сколько нейтронов в элементе,если относительная масса атома-52,а количество протонов-24?
Биология,
4 года назад
пожалуйста помогите!!!!
фрагмент молекулы ДНК содержит 60000 азотистых оснований из них на основание А приходится 10%.Найдите число оснований Ц.Г.Т …
Математика,
6 лет назад
помогите пожалйуста 3класс математика стр.7 н3…
Биология,
6 лет назад
Сколько см3 CO2 выдыхает человек при одном спокойном выдохе?
A) 4 см3 Б)20 см3 С)16см3 Д)80см3 Е)105см
Пожалуйста напишите с решением:)
2)Выберите относящиеся к жизненому объему легких?
1)…
Предмет: Биология,
автор: pusakova548
На терминалах аксона расположены специальные приспособления для передачи сигналов – химические …
Ответы
Автор ответа: 024857
0
Ответ:
скорее всего это синапсы
Объяснение:
Предыдущий вопрос
Следующий вопрос
Интересные вопросы
Предмет: Химия,
автор: hristuksofia96
а)Силіцій і Кисень →Силіцій(ıv) оксидів
3 года назад
Предмет: Химия,
автор: mvvip
Углекислый газ какого объёма ( н. у.) образуется при сгорании угарного газа объёмом (н. у) 2.24 м³ в избытке кислорода
3 года назад
Предмет: Английский язык,
автор: sashaberkita
помогите пжжж даю сто балов
12 і 13 вправу
3 года назад
Предмет: Алгебра,
автор: Student1111111111
φ(х)=2/х^8-10
ребят, мне нужно срочно в течении 10-15 минут.
6 лет назад
Предмет: Физика,
автор: Dvoechnik019
Помогите пожалуйста. срочно надо. Постройте изображение предмета AB , если в данный предмет находится в 3 фокусе собирающей линзы. Сделайте вывод.
6 лет назад
![]()
From Wikipedia, the free encyclopedia
This article is about chemical synapses of the nervous system. For general information, see synapse. For other uses, see synapse (disambiguation).

Artistic interpretation of the major elements in chemical synaptic transmission. An electrochemical wave called an action potential travels along the axon of a neuron. When the action potential reaches the presynaptic terminal, it provokes the release of a synaptic vesicle, secreting its quanta of neurotransmitter molecules. The neurotransmitter binds to chemical receptor molecules located in the membrane of another neuron, the postsynaptic neuron, on the opposite side of the synaptic cleft.
Chemical synapses are biological junctions through which neurons’ signals can be sent to each other and to non-neuronal cells such as those in muscles or glands. Chemical synapses allow neurons to form circuits within the central nervous system. They are crucial to the biological computations that underlie perception and thought. They allow the nervous system to connect to and control other systems of the body.
At a chemical synapse, one neuron releases neurotransmitter molecules into a small space (the synaptic cleft) that is adjacent to another neuron. The neurotransmitters are contained within small sacs called synaptic vesicles, and are released into the synaptic cleft by exocytosis. These molecules then bind to neurotransmitter receptors on the postsynaptic cell. Finally, the neurotransmitters are cleared from the synapse through one of several potential mechanisms including enzymatic degradation or re-uptake by specific transporters either on the presynaptic cell or on some other neuroglia to terminate the action of the neurotransmitter.
The adult human brain is estimated to contain from 1014 to 5 × 1014 (100–500 trillion) synapses.[1] Every cubic millimeter of cerebral cortex contains roughly a billion (short scale, i.e. 109) of them.[2] The number of synapses in the human cerebral cortex has separately been estimated at 0.15 quadrillion (150 trillion)[3]
The word «synapse» was introduced by Sir Charles Scott Sherrington in 1897.[4] Chemical synapses are not the only type of biological synapse: electrical and immunological synapses also exist. Without a qualifier, however, «synapse» commonly refers to chemical synapse.
Structure[edit]
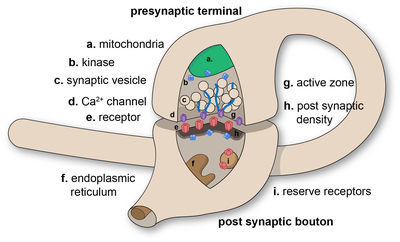
Diagram of a chemical synaptic connection
Further information on formation of synapses: Synaptogenesis
Synapses are functional connections between neurons, or between neurons and other types of cells.[5][6] A typical neuron gives rise to several thousand synapses, although there are some types that make far fewer.[7] Most synapses connect axons to dendrites,[8][9] but there are also other types of connections, including axon-to-cell-body,[10][11] axon-to-axon,[10][11] and dendrite-to-dendrite.[9] Synapses are generally too small to be recognizable using a light microscope except as points where the membranes of two cells appear to touch, but their cellular elements can be visualized clearly using an electron microscope.
Chemical synapses pass information directionally from a presynaptic cell to a postsynaptic cell and are therefore asymmetric in structure and function. The presynaptic axon terminal, or synaptic bouton, is a specialized area within the axon of the presynaptic cell that contains neurotransmitters enclosed in small membrane-bound spheres called synaptic vesicles (as well as a number of other supporting structures and organelles, such as mitochondria and endoplasmic reticulum). Synaptic vesicles are docked at the presynaptic plasma membrane at regions called active zones.
Immediately opposite is a region of the postsynaptic cell containing neurotransmitter receptors; for synapses between two neurons the postsynaptic region may be found on the dendrites or cell body. Immediately behind the postsynaptic membrane is an elaborate complex of interlinked proteins called the postsynaptic density (PSD).
Proteins in the PSD are involved in anchoring and trafficking neurotransmitter receptors and modulating the activity of these receptors. The receptors and PSDs are often found in specialized protrusions from the main dendritic shaft called dendritic spines.
Synapses may be described as symmetric or asymmetric. When examined under an electron microscope, asymmetric synapses are characterized by rounded vesicles in the presynaptic cell, and a prominent postsynaptic density. Asymmetric synapses are typically excitatory. Symmetric synapses in contrast have flattened or elongated vesicles, and do not contain a prominent postsynaptic density. Symmetric synapses are typically inhibitory.
The synaptic cleft—also called synaptic gap—is a gap between the pre- and postsynaptic cells that is about 20 nm (0.02 μ) wide.[12] The small volume of the cleft allows neurotransmitter concentration to be raised and lowered rapidly.[13]
An autapse is a chemical (or electrical) synapse formed when the axon of one neuron synapses with its own dendrites.
Signaling in chemical synapses[edit]
Overview[edit]
Here is a summary of the sequence of events that take place in synaptic transmission from a presynaptic neuron to a postsynaptic cell. Each step is explained in more detail below. Note that with the exception of the final step, the entire process may run only a few hundred microseconds, in the fastest synapses.[14]
- The process begins with a wave of electrochemical excitation called an action potential traveling along the membrane of the presynaptic cell, until it reaches the synapse.
- The electrical depolarization of the membrane at the synapse causes channels to open that are permeable to calcium ions.
- Calcium ions flow through the presynaptic membrane, rapidly increasing the calcium concentration in the interior.
- The high calcium concentration activates a set of calcium-sensitive proteins attached to vesicles that contain a neurotransmitter chemical.
- These proteins change shape, causing the membranes of some «docked» vesicles to fuse with the membrane of the presynaptic cell, thereby opening the vesicles and dumping their neurotransmitter contents into the synaptic cleft, the narrow space between the membranes of the pre- and postsynaptic cells.
- The neurotransmitter diffuses within the cleft. Some of it escapes, but some of it binds to chemical receptor molecules located on the membrane of the postsynaptic cell.
- The binding of neurotransmitter causes the receptor molecule to be activated in some way. Several types of activation are possible, as described in more detail below. In any case, this is the key step by which the synaptic process affects the behavior of the postsynaptic cell.
- Due to thermal vibration, the motion of atoms, vibrating about their equilibrium positions in a crystalline solid, neurotransmitter molecules eventually break loose from the receptors and drift away.
- The neurotransmitter is either reabsorbed by the presynaptic cell, and then repackaged for future release, or else it is broken down metabolically.
Neurotransmitter release[edit]

Release of neurotransmitter occurs at the end of axonal branches.
The release of a neurotransmitter is triggered by the arrival of a nerve impulse (or action potential) and occurs through an unusually rapid process of cellular secretion (exocytosis). Within the presynaptic nerve terminal, vesicles containing neurotransmitter are localized near the synaptic membrane. The arriving action potential produces an influx of calcium ions through voltage-dependent, calcium-selective ion channels at the down stroke of the action potential (tail current).[15] Calcium ions then bind to synaptotagmin proteins found within the membranes of the synaptic vesicles, allowing the vesicles to fuse with the presynaptic membrane.[16] The fusion of a vesicle is a stochastic process, leading to frequent failure of synaptic transmission at the very small synapses that are typical for the central nervous system. Large chemical synapses (e.g. the neuromuscular junction), on the other hand, have a synaptic release probability, in effect, of 1. Vesicle fusion is driven by the action of a set of proteins in the presynaptic terminal known as SNAREs. As a whole, the protein complex or structure that mediates the docking and fusion of presynaptic vesicles is called the active zone.[17] The membrane added by the fusion process is later retrieved by endocytosis and recycled for the formation of fresh neurotransmitter-filled vesicles.
An exception to the general trend of neurotransmitter release by vesicular fusion is found in the type II receptor cells of mammalian taste buds. Here the neurotransmitter ATP is released directly from the cytoplasm into the synaptic cleft via voltage gated channels.[18]
Receptor binding[edit]
Receptors on the opposite side of the synaptic gap bind neurotransmitter molecules. Receptors can respond in either of two general ways. First, the receptors may directly open ligand-gated ion channels in the postsynaptic cell membrane, causing ions to enter or exit the cell and changing the local transmembrane potential.[14] The resulting change in voltage is called a postsynaptic potential. In general, the result is excitatory in the case of depolarizing currents, and inhibitory in the case of hyperpolarizing currents. Whether a synapse is excitatory or inhibitory depends on what type(s) of ion channel conduct the postsynaptic current(s), which in turn is a function of the type of receptors and neurotransmitter employed at the synapse. The second way a receptor can affect membrane potential is by modulating the production of chemical messengers inside the postsynaptic neuron. These second messengers can then amplify the inhibitory or excitatory response to neurotransmitters.[14]
Termination[edit]
After a neurotransmitter molecule binds to a receptor molecule, it must be removed to allow for the postsynaptic membrane to continue to relay subsequent EPSPs and/or IPSPs. This removal can happen through one or more processes:
- The neurotransmitter may diffuse away due to thermally-induced oscillations of both it and the receptor, making it available to be broken down metabolically outside the neuron or to be reabsorbed.[19]
- Enzymes within the subsynaptic membrane may inactivate/metabolize the neurotransmitter.
- Reuptake pumps may actively pump the neurotransmitter back into the presynaptic axon terminal for reprocessing and re-release following a later action potential.[19]
Synaptic strength[edit]
The strength of a synapse has been defined by Sir Bernard Katz as the product of (presynaptic) release probability pr, quantal size q (the postsynaptic response to the release of a single neurotransmitter vesicle, a ‘quantum’), and n, the number of release sites. «Unitary connection» usually refers to an unknown number of individual synapses connecting a presynaptic neuron to a postsynaptic neuron.
The amplitude of postsynaptic potentials (PSPs) can be as low as 0.4 mV to as high as 20 mV.[20] The amplitude of a PSP can be modulated by neuromodulators or can change as a result of previous activity. Changes in the synaptic strength can be short-term, lasting seconds to minutes, or long-term (long-term potentiation, or LTP), lasting hours. Learning and memory are believed to result from long-term changes in synaptic strength, via a mechanism known as synaptic plasticity.
Receptor desensitization[edit]
Desensitization of the postsynaptic receptors is a decrease in response to the same neurotransmitter stimulus. It means that the strength of a synapse may in effect diminish as a train of action potentials arrive in rapid succession – a phenomenon that gives rise to the so-called frequency dependence of synapses. The nervous system exploits this property for computational purposes, and can tune its synapses through such means as phosphorylation of the proteins involved.
Synaptic plasticity[edit]
Synaptic transmission can be changed by previous activity. These changes are called synaptic plasticity and may result in either a decrease in the efficacy of the synapse, called depression, or an increase in efficacy, called potentiation. These changes can either be long-term or short-term. Forms of short-term plasticity include synaptic fatigue or depression and synaptic augmentation. Forms of long-term plasticity include long-term depression and long-term potentiation. Synaptic plasticity can be either homosynaptic (occurring at a single synapse) or heterosynaptic (occurring at multiple synapses).
Homosynaptic plasticity[edit]
Homosynaptic plasticity (or also homotropic modulation) is a change in the synaptic strength that results from the history of activity at a particular synapse. This can result from changes in presynaptic calcium as well as feedback onto presynaptic receptors, i.e. a form of autocrine signaling. Homosynaptic plasticity can affect the number and replenishment rate of vesicles or it can affect the relationship between calcium and vesicle release. Homosynaptic plasticity can also be postsynaptic in nature. It can result in either an increase or decrease in synaptic strength.
One example is neurons of the sympathetic nervous system (SNS), which release noradrenaline, which, besides affecting postsynaptic receptors, also affects presynaptic α2-adrenergic receptors, inhibiting further release of noradrenaline.[21] This effect is utilized with clonidine to perform inhibitory effects on the SNS.
Heterosynaptic plasticity[edit]
Heterosynaptic plasticity (or also heterotropic modulation) is a change in synaptic strength that results from the activity of other neurons. Again, the plasticity can alter the number of vesicles or their replenishment rate or the relationship between calcium and vesicle release. Additionally, it could directly affect calcium influx. Heterosynaptic plasticity can also be postsynaptic in nature, affecting receptor sensitivity.
One example is again neurons of the sympathetic nervous system, which release noradrenaline, which, in addition, generates an inhibitory effect on presynaptic terminals of neurons of the parasympathetic nervous system.[21]
Integration of synaptic inputs[edit]
In general, if an excitatory synapse is strong enough, an action potential in the presynaptic neuron will trigger an action potential in the postsynaptic cell. In many cases the excitatory postsynaptic potential (EPSP) will not reach the threshold for eliciting an action potential. When action potentials from multiple presynaptic neurons fire simultaneously, or if a single presynaptic neuron fires at a high enough frequency, the EPSPs can overlap and summate. If enough EPSPs overlap, the summated EPSP can reach the threshold for initiating an action potential. This process is known as summation, and can serve as a high pass filter for neurons.[22]
On the other hand, a presynaptic neuron releasing an inhibitory neurotransmitter, such as GABA, can cause an inhibitory postsynaptic potential (IPSP) in the postsynaptic neuron, bringing the membrane potential farther away from the threshold, decreasing its excitability and making it more difficult for the neuron to initiate an action potential. If an IPSP overlaps with an EPSP, the IPSP can in many cases prevent the neuron from firing an action potential. In this way, the output of a neuron may depend on the input of many different neurons, each of which may have a different degree of influence, depending on the strength and type of synapse with that neuron. John Carew Eccles performed some of the important early experiments on synaptic integration, for which he received the Nobel Prize for Physiology or Medicine in 1963.
Volume transmission[edit]
When a neurotransmitter is released at a synapse, it reaches its highest concentration inside the narrow space of the synaptic cleft, but some of it is certain to diffuse away before being reabsorbed or broken down. If it diffuses away, it has the potential to activate receptors that are located either at other synapses or on the membrane away from any synapse. The extrasynaptic activity of a neurotransmitter is known as volume transmission.[23] It is well established that such effects occur to some degree, but their functional importance has long been a matter of controversy.[24]
Recent work indicates that volume transmission may be the predominant mode of interaction for some special types of neurons. In the mammalian cerebral cortex, a class of neurons called neurogliaform cells can inhibit other nearby cortical neurons by releasing the neurotransmitter GABA into the extracellular space.[25] Along the same vein, GABA released from neurogliaform cells into the extracellular space also acts on surrounding astrocytes, assigning a role for volume transmission in the control of ionic and neurotransmitter homeostasis.[26] Approximately 78% of neurogliaform cell boutons do not form classical synapses. This may be the first definitive example of neurons communicating chemically where classical synapses are not present.[25]
Relationship to electrical synapses[edit]
An electrical synapse is an electrically conductive link between two abutting neurons that is formed at a narrow gap between the pre- and postsynaptic cells, known as a gap junction. At gap junctions, cells approach within about 3.5 nm of each other, rather than the 20 to 40 nm distance that separates cells at chemical synapses.[27][28] As opposed to chemical synapses, the postsynaptic potential in electrical synapses is not caused by the opening of ion channels by chemical transmitters, but rather by direct electrical coupling between both neurons. Electrical synapses are faster than chemical synapses.[13] Electrical synapses are found throughout the nervous system, including in the retina, the reticular nucleus of the thalamus, the neocortex, and in the hippocampus.[29] While chemical synapses are found between both excitatory and inhibitory neurons, electrical synapses are most commonly found between smaller local inhibitory neurons. Electrical synapses can exist between two axons, two dendrites, or between an axon and a dendrite.[30][31] In some fish and amphibians, electrical synapses can be found within the same terminal of a chemical synapse, as in Mauthner cells.[32]
Effects of drugs[edit]
One of the most important features of chemical synapses is that they are the site of action for the majority of psychoactive drugs. Synapses are affected by drugs, such as curare, strychnine, cocaine, morphine, alcohol, LSD, and countless others. These drugs have different effects on synaptic function, and often are restricted to synapses that use a specific neurotransmitter. For example, curare is a poison that stops acetylcholine from depolarizing the postsynaptic membrane, causing paralysis. Strychnine blocks the inhibitory effects of the neurotransmitter glycine, which causes the body to pick up and react to weaker and previously ignored stimuli, resulting in uncontrollable muscle spasms. Morphine acts on synapses that use endorphin neurotransmitters, and alcohol increases the inhibitory effects of the neurotransmitter GABA. LSD interferes with synapses that use the neurotransmitter serotonin. Cocaine blocks reuptake of dopamine and therefore increases its effects.
History and etymology[edit]
During the 1950s, Bernard Katz and Paul Fatt observed spontaneous miniature synaptic currents at the frog neuromuscular junction.[33] Based on these observations, they developed the ‘quantal hypothesis’ that is the basis for our current understanding of neurotransmitter release as exocytosis and for which Katz received the Nobel Prize in Physiology or Medicine in 1970.[34] In the late 1960s, Ricardo Miledi and Katz advanced the hypothesis that depolarization-induced influx of calcium ions triggers exocytosis.
Sir Charles Scott Sherringtonin coined the word ‘synapse’ and the history of the word was given by Sherrington in a letter he wrote to John Fulton:
‘I felt the need of some name to call the junction between nerve-cell and nerve-cell… I suggested using «syndesm»… He [ Sir Michael Foster ] consulted his Trinity friend Verrall, the Euripidean scholar, about it, and Verrall suggested «synapse» (from the Greek «clasp»).’–Charles Scott Sherrington[4]
See also[edit]
- Acclimatisation (neurones)
- Neuroscience
- Ribbon synapse
Notes[edit]
- ^ Drachman D (2005). «Do we have brain to spare?». Neurology. 64 (12): 2004–5. doi:10.1212/01.WNL.0000166914.38327.BB. PMID 15985565. S2CID 38482114.
- ^ Alonso-Nanclares L, Gonzalez-Soriano J, Rodriguez JR, DeFelipe J (September 2008). «Gender differences in human cortical synaptic density». Proc. Natl. Acad. Sci. U.S.A. 105 (38): 14615–9. Bibcode:2008PNAS..10514615A. doi:10.1073/pnas.0803652105. PMC 2567215. PMID 18779570.
- ^ Brain Facts and Figures Washington University.
- ^ a b Cowan, W. Maxwell; Südhof, Thomas C.; Stevens, Charles F. (2003). Synapses. JHU Press. p. 11. ISBN 9780801871184. Retrieved 9 June 2020.
- ^
Rapport, Richard L. (2005). Nerve Endings: The Discovery of the Synapse. W. W. Norton & Company. pp. 1–37. ISBN 978-0-393-06019-5.
- ^
Squire, Larry R.; Floyd Bloom; Nicholas Spitzer (2008). Fundamental Neuroscience. Academic Press. pp. 425–6. ISBN 978-0-12-374019-9. - ^
Hyman, Steven E.; Eric Jonathan Nestler (1993). The Molecular Foundations of Psychiatry. American Psychiatric Pub. pp. 425–6. ISBN 978-0-88048-353-7.
- ^
Smilkstein, Rita (2003). We’re Born to Learn: Using the Brain’s Natural Learning Process to Create Today’s Curriculum. Corwin Press. p. 56. ISBN 978-0-7619-4642-7.
- ^ a b Lytton, William W. (2002). From Computer to Brain: Foundations of Computational Neuroscience. Springer. p. 28. ISBN 978-0-387-95526-1. Axons connecting dendrite to dendrite are dendrodendritic synapses. Axons which connect axon to dendrite are called axodendritic synapses
- ^ a b
Garber, Steven D. (2002). Biology: A Self-Teaching Guide. John Wiley and Sons. p. 175. ISBN 978-0-471-22330-6.
synapses connect axons to cell body.
- ^ a b
Weiss, Mirin; Dr Steven M. Mirin; Dr Roxanne Bartel (1994). Cocaine. American Psychiatric Pub. p. 52. ISBN 978-1-58562-138-5. Retrieved 2008-12-26. Axons terminating on the postsynaptic cell body are axosomatic synapses. Axons that terminate on axons are axoaxonic synapses
- ^ Widrow, Bernard; Kim, Youngsik; Park, Dookun; Perin, Jose Krause (2019). «Nature’s Learning Rule». Artificial Intelligence in the Age of Neural Networks and Brain Computing. Elsevier. pp. 1–30. doi:10.1016/b978-0-12-815480-9.00001-3. ISBN 978-0-12-815480-9. S2CID 125516633.
- ^ a b Kandel, Schwartz & Jessell 2000, p. 182
- ^ a b c Bear, Mark F; Connors, Barry W; Paradiso, Michael A (2007). Neuroscience: exploring the brain. Philadelphia, PA: Lippincott Williams & Wilkins. pp. 113–118.
- ^
Llinás R, Steinberg IZ, Walton K (1981). «Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse». Biophysical Journal. 33 (3): 323–351. Bibcode:1981BpJ….33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850. - ^ Chapman, Edwin R. (2002). «Synaptotagmin: A Ca2+ sensor that triggers exocytosis?». Nature Reviews Molecular Cell Biology. 3 (7): 498–508. doi:10.1038/nrm855. ISSN 1471-0080. PMID 12094216. S2CID 12384262.
- ^ Craig C. Garner and Kang Shen. Structure and Function of Vertebrate and Invertebrate Active Zones. Structure and Functional Organization of the Synapse. Ed: Johannes Hell and Michael Ehlers. Springer, 2008.
- ^ Romanov, Roman A.; Lasher, Robert S.; High, Brigit; Savidge, Logan E.; Lawson, Adam; Rogachevskaja, Olga A.; Zhao, Haitian; Rogachevsky, Vadim V.; Bystrova, Marina F.; Churbanov, Gleb D.; Adameyko, Igor; Harkany, Tibor; Yang, Ruibiao; Kidd, Grahame J.; Marambaud, Philippe; Kinnamon, John C.; Kolesnikov, Stanislav S.; Finger, Thomas E. (2018). «Chemical synapses without synaptic vesicles: Purinergic neurotransmission through a CALHM1 channel-mitochondrial signaling complex». Science Signaling. 11 (529): eaao1815. doi:10.1126/scisignal.aao1815. ISSN 1945-0877. PMC 5966022. PMID 29739879.
- ^ a b Sherwood L., stikawy (2007). Human Physiology 6e: From Cells to Systems
- ^ Díaz-Ríos M, Miller MW (June 2006). «Target-specific regulation of synaptic efficacy in the feeding central pattern generator of Aplysia: potential substrates for behavioral plasticity?». Biol. Bull. 210 (3): 215–29. doi:10.2307/4134559. JSTOR 4134559. PMID 16801496. S2CID 34154835.
- ^ a b Rang, H.P.; Dale, M.M.; Ritter, J.M. (2003). Pharmacology (5th ed.). Edinburgh: Churchill Livingstone. p. 129. ISBN 978-0-443-07145-4.
- ^ Bruce Alberts; Alexander Johnson; Julian Lewis; Martin Raff; Keith Roberts; Peter Walter, eds. (2002). «Ch. 11. Section: Single Neurons Are Complex Computation Devices». Molecular Biology of the Cell (4th ed.). Garland Science. ISBN 978-0-8153-3218-3.
- ^ Zoli M, Torri C, Ferrari R, et al. (1998). «The emergence of the volume transmission concept». Brain Res. Brain Res. Rev. 26 (2–3): 136–47. doi:10.1016/S0165-0173(97)00048-9. PMID 9651506. S2CID 20495134.
- ^ Fuxe K, Dahlström A, Höistad M, et al. (2007). «From the Golgi-Cajal mapping to the transmitter-based characterization of the neuronal networks leading to two modes of brain communication: wiring and volume transmission» (PDF). Brain Res Rev. 55 (1): 17–54. doi:10.1016/j.brainresrev.2007.02.009. hdl:10447/9980. PMID 17433836. S2CID 1323780.
- ^ a b Oláh S, Füle M, Komlósi G, et al. (2009). «Regulation of cortical microcircuits by unitary GABA-mediated volume transmission». Nature. 461 (7268): 1278–81. Bibcode:2009Natur.461.1278O. doi:10.1038/nature08503. PMC 2771344. PMID 19865171.
- ^ Rózsa M, Baka J, Bordé S, Rózsa B, Katona G, Tamás G, et al. (2015). «Unitary GABAergic volume transmission from individual interneurons to astrocytes in the cerebral cortex» (PDF). Brain Structure and Function. 222 (1): 651–659. doi:10.1007/s00429-015-1166-9. PMID 26683686. S2CID 30728927.
- ^ Kandel, Schwartz & Jessell 2000, p. 176
- ^ Hormuzdi et al. 2004
- ^ Connors BW, Long MA (2004). «Electrical synapses in the mammalian brain». Annu. Rev. Neurosci. 27 (1): 393–418. doi:10.1146/annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Veruki ML, Hartveit E (December 2002). «Electrical synapses mediate signal transmission in the rod pathway of the mammalian retina». J. Neurosci. 22 (24): 10558–66. doi:10.1523/JNEUROSCI.22-24-10558.2002. PMC 6758447. PMID 12486148.
- ^ Bennett MV, Pappas GD, Aljure E, Nakajima Y (March 1967). «Physiology and ultrastructure of electrotonic junctions. II. Spinal and medullary electromotor nuclei in mormyrid fish». J. Neurophysiol. 30 (2): 180–208. doi:10.1152/jn.1967.30.2.180. PMID 4167209.
- ^ Pereda AE, Rash JE, Nagy JI, Bennett MV (December 2004). «Dynamics of electrical transmission at club endings on the Mauthner cells». Brain Res. Brain Res. Rev. 47 (1–3): 227–44. CiteSeerX 10.1.1.662.9352. doi:10.1016/j.brainresrev.2004.06.010. PMID 15572174. S2CID 9527518.
- ^ Augustine, George J.; Kasai, Haruo (2007-02-01). «Bernard Katz, quantal transmitter release and the foundations of presynaptic physiology». The Journal of Physiology. 578 (Pt 3): 623–625. doi:10.1113/jphysiol.2006.123224. PMC 2151334. PMID 17068096.
- ^ «Nobel prize». British Medical Journal. 4 (5729): 190. 1970-10-24. doi:10.1136/bmj.4.5729.190. PMC 1819734. PMID 4320287.
References[edit]
- Carlson, Neil R. (2007). Physiology of Behavior (9th ed.). Boston, MA: Pearson Education. ISBN 978-0-205-59389-7.
- Kandel, Eric R.; Schwartz, James H.; Jessell, Thomas M. (2000). Principles of Neural Science (4th ed.). New York: McGraw-Hill. ISBN 978-0-8385-7701-1.
- Llinás R, Sugimori M, Simon SM (April 1982). «Transmission by presynaptic spike-like depolarization in the squid giant synapse». Proc. Natl. Acad. Sci. U.S.A. 79 (7): 2415–9. Bibcode:1982PNAS…79.2415L. doi:10.1073/pnas.79.7.2415. PMC 346205. PMID 6954549.
- Llinás R, Steinberg IZ, Walton K (1981). «Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse». Biophysical Journal. 33 (3): 323–352. Bibcode:1981BpJ….33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850.
- Bear, Mark F.; Connors, Barry W.; Paradiso, Michael A. (2001). Neuroscience: Exploring the Brain. Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-3944-3.
- Hormuzdi, SG; Filippov, MA; Mitropoulou, G; Monyer, H; Bruzzone, R (March 2004). «Electrical synapses: a dynamic signaling system that shapes the activity of neuronal networks». Biochim Biophys Acta. 1662 (1–2): 113–137. doi:10.1016/j.bbamem.2003.10.023. PMID 15033583.
- Karp, Gerald (2005). Cell and Molecular Biology: concepts and experiments (4th ed.). Hoboken, NJ: John Wiley & Sons. ISBN 978-0-471-46580-5.
- Nicholls, J.G.; Martin, A.R.; Wallace, B.G.; Fuchs, P.A. (2001). From Neuron to Brain (4th ed.). Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-439-3.
External links[edit]
![]()
This audio file was created from a revision of this article dated 19 June 2005, and does not reflect subsequent edits.
- Synapse Review for Kids
- Synapse – Cell Centered Database
- Atlas of Ultrastructure Neurocytology An electron microscope picture gallery assembled by Kristen Harris’ lab of synapses and other neuronal structures.
From Wikipedia, the free encyclopedia

Activity at an axon terminal: Neuron A is transmitting a signal at the axon terminal to neuron B (receiving). Features: 1. Mitochondrion. 2. Synaptic vesicle with neurotransmitters. 3. Autoreceptor. 4. Synapse with neurotransmitter released (serotonin). 5.Postsynaptic receptors activated by neurotransmitter (induction of a postsynaptic potential). 6. Calcium channel. 7. Exocytosis of a vesicle. 8. Recaptured neurotransmitter.
Axon terminals (also called synaptic boutons, terminal boutons, or end-feet) are distal terminations of the telodendria (branches) of an axon. An axon, also called a nerve fiber, is a long, slender projection of a nerve cell, or neuron, that conducts electrical impulses called action potentials away from the neuron’s cell body, or soma, in order to transmit those impulses to other neurons, muscle cells or glands.
Neurons are interconnected in complex arrangements, and use electrochemical signals and neurotransmitter chemicals to transmit impulses from one neuron to the next; axon terminals are separated from neighboring neurons by a small gap called a synapse, across which impulses are sent. The axon terminal, and the neuron from which it comes, is sometimes referred to as the «presynaptic» neuron.
Nerve impulse release[edit]
Neurotransmitters are packaged into synaptic vesicles that cluster beneath the axon terminal membrane on the presynaptic side of a synapse. The axonal terminals are specialized to release the neurotransmitters of the presynaptic cell.[1] The terminals release transmitter substances into a gap called the synaptic cleft between the terminals and the dendrites of the next neuron. The information is received by the dendrite receptors of the postsynaptic cell that are connected to it. Neurons don’t touch each other, but communicate across the synapse.[2]
The neurotransmitter molecule packages (vesicles) are created within the neuron, then travel down the axon to the distal axon terminal where they sit docked. Calcium ions then trigger a biochemical cascade which results in vesicles fusing with the presynaptic membrane and releasing their contents to the synaptic cleft within 180 µs of calcium entry.[3] Triggered by the binding of the calcium ions, the synaptic vesicle proteins begin to move apart, resulting in the creation of a fusion pore. The presence of the pore allows for the release of neurotransmitter into the synaptic cleft.[4][5] The process occurring at the axon terminal is exocytosis,[6] which a cell uses to exude secretory vesicles out of the cell membrane. These membrane-bound vesicles contain soluble proteins to be secreted to the extracellular environment, as well as membrane proteins and lipids that are sent to become components of the cell membrane. Exocytosis in neuronal chemical synapses is Ca2+ triggered and serves interneuronal signalling.[7]
Mapping activity[edit]
| Neuron |
|---|
|
Dendrite Soma Axon Nucleus Node of Axon terminal Schwann cell Myelin sheath |

Wade Regehr, a Professor of Neurobiology at Harvard Medical School’s Department of Neurobiology, developed a method to physiologically see the synaptic activity that occurs in the brain. A dye alters the fluorescence properties when attached to calcium. Using fluorescence-microscopy techniques calcium levels are detected, and therefore the influx of calcium in the presynaptic neuron.[8] Regehr’s laboratory specializes in pre-synaptic calcium dynamics which occurs at the axon terminals. Regehr studies the implication of calcium Ca2+ as it affects synaptic strength.[9][self-published source?][10] By studying the physiological process and mechanisms, a further understanding is made of neurological disorders such as epilepsy, schizophrenia and major depressive disorder, as well as memory and learning.[11][12]
See also[edit]
- Endoplasmic reticulum
- Golgi apparatus
- Micelle
- Membrane nanotube
- Endocytosis
- Vesicular monoamine transporter
Further reading[edit]
- Cragg SJ, Greenfield SA (August 1997). «Differential autoreceptor control of somatodendritic and axon terminal dopamine release in substantia nigra, ventral tegmental area, and striatum». The Journal of Neuroscience. 17 (15): 5738–46. doi:10.1523/JNEUROSCI.17-15-05738.1997. PMC 6573186. PMID 9221772.
- Vaquero CF, de la Villa P (October 1999). «Localisation of the GABA(C) receptors at the axon terminal of the rod bipolar cells of the mouse retina». Neuroscience Research. 35 (1): 1–7. doi:10.1016/S0168-0102(99)00050-4. PMID 10555158. S2CID 53189471.
- Roffler-Tarlov S, Beart PM, O’Gorman S, Sidman RL (May 1979). «Neurochemical and morphological consequences of axon terminal degeneration in cerebellar deep nuclei of mice with inherited Purkinje cell degeneration». Brain Research. 168 (1): 75–95. doi:10.1016/0006-8993(79)90129-X. PMID 455087. S2CID 19618884.
- Yagi T, Kaneko A (February 1988). «The axon terminal of goldfish retinal horizontal cells: a low membrane conductance measured in solitary preparations and its implication to the signal conduction from the soma». Journal of Neurophysiology. 59 (2): 482–94. doi:10.1152/jn.1988.59.2.482. PMID 3351572.
- LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite.[13]
References[edit]
- ^ «Axon Terminal». Medical Dictionary Online. Archived from the original on 2016-03-04. Retrieved February 6, 2013.
- ^ Foster, Sally. «Axon Terminal — Synaptic Vesicle — Neurotransmitter». Retrieved February 6, 2013.
- ^ Llinás R, Steinberg IZ, Walton K (March 1981). «Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse». Biophysical Journal. 33 (3): 323–51. Bibcode:1981BpJ….33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850.
- ^ Carlson, 2007, p.56
- ^ Chudler EH (November 24, 2011). «Neuroscience for kids Neurotransmitters and Neuroactive Peptides». Archived from the original on December 18, 2008. Retrieved February 6, 2013.
- ^ Rizo, Josep (2018-07-10). «Mechanism of neurotransmitter release coming into focus». Protein Science (Review). 27 (8): 1364–1391. doi:10.1002/pro.3445. ISSN 0961-8368. PMC 6153415. PMID 29893445.
Research for three decades and major recent advances have provided crucial insights into how neurotransmitters are released by Ca2+ -triggered synaptic vesicle exocytosis, leading to reconstitution of basic steps that underlie Ca2+ -dependent membrane fusion and yielding a model that assigns defined functions for central components of the release machinery.
- ^ Südhof TC, Rizo J (December 2011). «Synaptic vesicle exocytosis». Cold Spring Harbor Perspectives in Biology. 3 (12): a005637. doi:10.1101/cshperspect.a005637. PMC 3225952. PMID 22026965.
- ^
Sauber C. «Focus October 20-Neurobiology VISUALIZING THE SYNAPTIC CONNECTION». Archived from the original on 2006-09-01. Retrieved July 3, 2013. - ^
Regehr W (1999–2008). «Wade Regehr, Ph.D.» Archived from the original on February 18, 2010. Retrieved July 3, 2013. - ^ President and Fellows of Harvard College (2008). «The Neurobiology Department at Harvard Medical School». Archived from the original on 20 December 2008. Retrieved July 3, 2013.
- ^ «NINDS Announces New Javits Neuroscience Investigator Awardees» (Press release). National Institute of Neurological Disorders and Stroke. May 4, 2005. Archived from the original on January 17, 2009. Retrieved February 6, 2013.
- ^ «Scholar Awards». The McKnight Endowment Fund for Neuroscience. Archived from the original on 2004-05-08. Retrieved July 3, 2013.
- ^ Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D (November 1999). «LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite». Nature. 402 (6760): 421–5. Bibcode:1999Natur.402..421T. doi:10.1038/46574. PMID 10586883. S2CID 205056308.
Художественная интерпретация основных элементов в химической синаптической передаче. Электрохимическая волна, называемая потенциалом действия, распространяется вдоль аксона нейрона. Когда потенциал действия достигает пресинаптического конца, он вызывает высвобождение синаптического пузырька, секретируя его кванты молекул нейромедиатора. Нейромедиатор связывается с молекулами химических рецепторов, расположенными в мембране другого нейрона, постсинаптического нейрона, на противоположной стороне синаптической щели.
Химические синапсы — это биологические соединения, через которые нейроны ‘подают сигналы могут быть отправлены друг другу и ненейрональным клеткам, таким как клетки мышц или желез. Химические синапсы позволяют нейронам образовывать цепи внутри центральной нервной системы. Они имеют решающее значение для биологических вычислений, лежащих в основе восприятия и мышления. Они позволяют нервной системе подключаться к другим системам тела и контролировать их.
В химическом синапсе один нейрон высвобождает молекулы нейромедиатора в небольшое пространство (синаптическая щель ), прилегающее к другому нейрону. Нейротрансмиттеры содержатся в небольших мешочках, называемых синаптическими пузырьками, и высвобождаются в синаптическую щель посредством экзоцитоза. Эти молекулы затем связываются с рецепторами нейротрансмиттеров на постсинаптической клетке. Наконец, нейротрансмиттеры выводятся из синапса с помощью одного из нескольких потенциальных механизмов, включая ферментативную деградацию или повторное поглощение специфическими переносчиками либо пресинаптической клеткой, либо какой-либо другой нейроглией для прекращения действие нейротрансмиттера.
Согласно оценкам, мозг взрослого человека содержит от 10 до 5 × 10 (100–500 триллионов) синапсов. Каждый кубический миллиметр коры головного мозга содержит примерно миллиард (в сокращенном масштабе, т.е. 10) их. Количество синапсов в коре головного мозга человека по отдельности оценивается в 0,15 квадриллиона (150 триллионов)
Слово «синапс» было введено сэром Чарльзом Скоттом Шеррингтоном в 1897 году. не единственный тип биологических синапсов: также существуют электрические и иммунологические синапсы. Однако без уточнения «синапс» обычно относится к химическому синапсу.
Содержание
- 1 Структура
- 2 Передача сигналов в химических синапсах
- 2.1 Обзор
- 2.2 Высвобождение нейротрансмиттера
- 2.3 Связывание рецептора
- 2.4 Прекращение действия
- 3 Синаптическая сила
- 4 Рецептор десенсибилизация
- 5 Синаптическая пластичность
- 5.1 Гомосинаптическая пластичность
- 5.2 Гетеросинаптическая пластичность
- 6 Интеграция синаптических входов
- 7 Объемная передача
- 8 Связь с электрическими синапсами
- 9 Действие лекарств
- 10 История и этимология
- 11 См. Также
- 12 Примечания
- 13 Ссылки
- 14 Внешние ссылки
Структура
| Структура типичного химического синапса |
|---|
 Постсинаптический. плотность Напряжение-. закрытый Ca. канал Синаптический. везикула Нейротрансмиттер. транспортер Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит Постсинаптический. плотность Напряжение-. закрытый Ca. канал Синаптический. везикула Нейротрансмиттер. транспортер Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит |
| Различать пре- и пост- синапс |
|---|
 ↓ К постсинаптическому. нейрону ↓ От пресинаптическому. нейрону Переносчик нейротрансмиттера Рецептор нейротрансмиттера Нейротрансмиттер передача Синаптическая щель { ↓ К постсинаптическому. нейрону ↓ От пресинаптическому. нейрону Переносчик нейротрансмиттера Рецептор нейротрансмиттера Нейротрансмиттер передача Синаптическая щель { |
| «Соединение, связывающее нейрон с нейроном, — это синапс. Сигнал проходит. в одном направлении, от пресинаптического нейрона к постсинаптическому нейрону. через синапс, который действует как переменный аттенюатор ». Вкратце,. направление потока сигнала определяет префикс для задействованного. синапсы. |
Синапсы — это функциональные связи между нейронами или между нейронами и другими типами клеток. Типичный нейрон дает начало нескольким тысячам синапсов, хотя есть некоторые типы, которые образуют намного меньше. Большинство синапсов соединяют аксоны От до дендритов, но существуют и другие типы соединений, включая аксон-к-телу клетки, аксон-аксон и дендрит-дендрит. Синапсы обычно слишком малы, чтобы их можно было распознать с помощью светового микроскопа, за исключением точек, где мембраны двух клеток, кажется, соприкасаются, но их клеточные элементы можно четко визуализировать с помощью электронного микроскопа.
Химические синапсы передают информацию направленно от пресинаптической клетки к постсинаптической клетке и, следовательно, асимметричны по структуре и функциям. Пресинаптический конец аксона, или синаптический бутон, представляет собой специализированную область в аксоне пресинаптической клетки, которая содержит нейротрансмиттеры, заключенные в небольшие связанные с мембраной сферы, называемые синаптическими пузырьками (а также ряд других поддерживающих структур и органелл, таких как митохондрии и эндоплазматический ретикулум ). Синаптические везикулы состыкованы с пресинаптической плазматической мембраной в областях, называемых активными зонами.
. Непосредственно напротив находится область постсинаптической клетки, содержащая нейротрансмиттерные рецепторы ; для синапсов между двумя нейронами постсинаптическая область может быть найдена на дендритах или теле клетки. Непосредственно за постсинаптической мембраной находится сложный комплекс взаимосвязанных белков, называемый постсинаптической плотностью (PSD).
Белки в PSD участвуют в закреплении и транспортировке рецепторов нейротрансмиттеров, а также в модуляции активности этих рецепторов. Рецепторы и PSD часто находятся в специализированных выступах от главного дендритного стержня, называемых дендритными шипами.
Синапсы могут быть описаны как симметричные или асимметричные. При исследовании под электронным микроскопом асимметричные синапсы характеризуются округлыми пузырьками в пресинаптической клетке и заметной постсинаптической плотностью. Асимметричные синапсы обычно возбуждающие. Симметричные синапсы, напротив, имеют уплощенные или удлиненные пузырьки и не содержат заметной постсинаптической плотности. Симметричные синапсы обычно тормозят.
синаптическая щель — также называемая синаптическая щель — это щель между пре- и постсинаптическими клетками шириной около 20 нм (0,02 мкм). Небольшой объем щели позволяет быстро повышать и понижать концентрацию нейромедиаторов.
autapse — это химический (или электрический) синапс, образующийся, когда аксон одного нейрона синапсируется с его собственными дендритами..
Передача сигналов в химических синапсах
Обзор
Вот краткое изложение последовательности событий, которые имеют место при синаптической передаче от пресинаптического нейрона к постсинаптической клетке. Ниже каждый шаг описан более подробно. Обратите внимание, что за исключением последнего шага, весь процесс может длиться всего несколько сотен микросекунд в самых быстрых синапсах.
- Процесс начинается с волны электрохимического возбуждения, называемой потенциалом действия, перемещающейся вдоль мембрана пресинаптической клетки, пока не достигнет синапса.
- Электрическая деполяризация мембраны в синапсе вызывает открытие каналов, проницаемых для ионов кальция.
- Ионы кальция проходят через пресинаптическую мембрану, быстро увеличивая концентрацию кальция внутри.
- Высокая концентрация кальция активирует набор кальций-чувствительных белков, прикрепленных к везикулам, которые содержат нейротрансмиттер химический.
- Эти белки изменяют форму, в результате чего мембраны некоторых «пристыкованных» везикул сливаются с мембраной пресинаптической клетки, тем самым открывая везикулы и сбрасывая их нейромедиаторное содержимое в синаптическую щель, узкое пространство будет между мембранами пре- и постсинаптических клеток.
- Нейромедиатор диффундирует в щель. Часть из них ускользает, но часть связывается с молекулами химического рецептора, расположенными на мембране постсинаптической клетки.
- Связывание нейромедиатора заставляет молекулу рецептора каким-то образом активироваться. Возможны несколько типов активации, которые более подробно описаны ниже. В любом случае, это ключевой шаг, с помощью которого синаптический процесс влияет на поведение постсинаптической клетки.
- Из-за тепловой вибрации движение атомов, колеблющихся вокруг их положений равновесия в кристаллическое твердое вещество, молекулы нейротрансмиттера в конечном итоге отрываются от рецепторов и уносятся прочь.
- Нейромедиатор либо реабсорбируется пресинаптической клеткой, а затем переупаковывается для будущего высвобождения, либо метаболически расщепляется.
Высвобождение нейротрансмиттера
Высвобождение нейротрансмиттера происходит на концах аксональных ветвей.
Высвобождение нейромедиатора запускается приходом нервного импульса (или потенциала действия ) и происходит через необычно быстрое процесс клеточной секреции (экзоцитоз ). Внутри пресинаптического нервного окончания везикулы, содержащие нейромедиатор, локализуются рядом с синаптической мембраной. Поступающий потенциал действия вызывает приток ионов кальция через зависимые от напряжения, кальций-селективные ионные каналы при нижнем ходе потенциала действия (хвостовой ток). Затем ионы кальция связываются с белками синаптотагмин, обнаруженными в мембранах синаптических везикул, позволяя везикулам сливаться с пресинаптической мембраной. Слияние пузырьков — это стохастический процесс, приводящий к частым сбоям синаптической передачи в очень маленьких синапсах, которые типичны для центральной нервной системы. С другой стороны, большие химические синапсы (например, нервно-мышечное соединение ) имеют вероятность синаптического высвобождения, равную 1. Слияние везикул управляется действием набора белков в пресинаптической терминал, известный как SNAREs. В целом, белковый комплекс или структура, которая опосредует стыковку и слияние пресинаптических везикул, называется активной зоной. Мембрана, добавленная в процессе слияния, позже извлекается посредством эндоцитоза и повторно используется для образования свежих везикул, заполненных нейротрансмиттерами.
Исключение из общей тенденции высвобождения нейромедиаторов посредством везикулярного слияния обнаруживается в рецепторных клетках типа II вкусовых рецепторов млекопитающих. Здесь нейротрансмиттер АТФ высвобождается непосредственно из цитоплазмы в синаптическую щель через каналы, управляемые напряжением.
Связывание рецептора
Рецепторы на противоположной стороне синаптической щели связывают нейротрансмиттер молекулы. Рецепторы могут реагировать одним из двух основных способов. Во-первых, рецепторы могут напрямую открывать лиганд-управляемые ионные каналы в постсинаптической клеточной мембране, заставляя ионы входить или выходить из клетки и изменяя локальный трансмембранный потенциал. Результирующее изменение напряжения называется постсинаптическим потенциалом. Как правило, результат является возбуждающим в случае деполяризующих токов и тормозящим в случае гиперполяризующих токов. Является ли синапс возбуждающим или тормозящим, зависит от того, какие типы ионных каналов проводят постсинаптические токи, которые, в свою очередь, зависят от типа рецепторов и нейротрансмиттеров, используемых в синапсе. Второй способ, которым рецептор может влиять на мембранный потенциал, — это модуляция производства химических мессенджеров внутри постсинаптического нейрона. Эти вторичные мессенджеры могут затем усилить тормозную или возбуждающую реакцию на нейротрансмиттеры.
Прерывание
После того, как молекула нейромедиатора связывается с молекулой рецептора, ее необходимо удалить, чтобы позволить постсинаптической мембране продолжить свое существование. ретранслировать последующие EPSP и / или IPSP. Это удаление может происходить в результате одного или нескольких процессов:
- Нейромедиатор может диффундировать из-за термически индуцированных колебаний как его, так и рецептора, делая его доступным для метаболического разложения вне нейрона или для реабсорбции.
- Ферменты в подсинаптической мембране могут инактивировать / метаболизировать нейромедиатор.
- Насосы обратного захвата могут активно перекачивать нейромедиатор обратно в пресинаптический конец аксона для повторной обработки и повторного высвобождения после более поздних действий
Сила синапса
Сила синапса была определена сэром Бернардом Кацем как продукт (пресинаптической) вероятности высвобождения pr, квантовой величины q (постсинаптический ответ на высвобождение одного везикулы нейромедиатора, «квант»), и n — количество мест высвобождения. «Унитарная связь» обычно относится к неизвестному количеству отдельных синапсов, соединяющих пресинаптический нейрон с постсинаптическим нейроном. Амплитуда постсинаптических потенциалов (ПСП) может составлять от 0,4 мВ до 20 мВ. Амплитуда PSP может модулироваться нейромодуляторами или может изменяться в результате предыдущей активности. Изменения в силе синапсов могут быть кратковременными, продолжительностью от секунд до минут, или долгосрочными (долгосрочное потенцирование, или LTP), продолжительностью несколько часов. Считается, что обучение и память являются результатом долговременных изменений синаптической силы посредством механизма, известного как синаптическая пластичность.
десенсибилизация рецепторов
Десенсибилизация постсинаптических рецепторов — это снижение в ответ на то же самое. стимул нейромедиатора. Это означает, что сила синапса может фактически уменьшаться по мере того, как последовательность потенциалов действия прибывает в быстрой последовательности — феномен, который приводит к так называемой частотной зависимости синапсов. Нервная система использует это свойство в вычислительных целях и может настраивать свои синапсы с помощью таких средств, как фосфорилирование задействованных белков.
Синаптическая пластичность
Синаптическая передача может быть изменена предыдущей активностью. Эти изменения называются синаптической пластичностью и могут привести либо к снижению эффективности синапса, называемой депрессией, либо к повышению эффективности, называемой потенцированием. Эти изменения могут быть долгосрочными или краткосрочными. Формы краткосрочной пластичности включают синаптическую усталость или депрессию и синаптическое увеличение. Формы долговременной пластичности включают длительную депрессию и длительную потенциацию. Синаптическая пластичность может быть либо гомосинаптической (происходящей в одном синапсе), либо гетеросинаптической (происходящей в нескольких синапсах).
Гомосинаптическая пластичность
Гомосинаптическая пластичность (или также гомотропная модуляция) — это изменение синаптической силы, которое является результатом истории активности конкретного синапса. Это может быть результатом изменений пресинаптического кальция, а также обратной связи с пресинаптическими рецепторами, то есть формой аутокринной передачи сигналов. Гомосинаптическая пластичность может влиять на количество и скорость пополнения везикул или может влиять на соотношение между высвобождением кальция и везикул. Гомосинаптическая пластичность также может иметь постсинаптическую природу. Это может привести либо к увеличению, либо к снижению синаптической силы.
Одним из примеров являются нейроны симпатической нервной системы (SNS), которые выделяют норадреналин, который, помимо воздействия на постсинаптические рецепторы, также влияет на пресинаптические α2- адренорецепторы, ингибирующие дальнейшее высвобождение норадреналина. Этот эффект используется с клонидином для оказания ингибирующего действия на SNS.
Гетеросинаптическая пластичность
Гетеросинаптическая пластичность (или также гетеротропная модуляция) — это изменение синаптической силы, которое возникает в результате активности других нейронов. Опять же, пластичность может изменять количество везикул или скорость их пополнения, или соотношение между высвобождением кальция и везикул. Кроме того, это может напрямую влиять на приток кальция. Гетеросинаптическая пластичность также может иметь постсинаптическую природу, влияя на чувствительность рецепторов.
Одним из примеров снова являются нейроны симпатической нервной системы, которые выделяют норадреналин, который, кроме того, оказывает ингибирующее действие на пресинаптические окончания нейронов парасимпатическая нервная система.
Интеграция синаптических входов
В целом, если возбуждающий синапс достаточно силен, потенциал действия в пресинаптическом нейроне сработает. потенциал действия в постсинаптической клетке. Во многих случаях возбуждающий постсинаптический потенциал (ВПСП) не достигает порога для выявления потенциала действия. Когда потенциалы действия от нескольких пресинаптических нейронов срабатывают одновременно или если один пресинаптический нейрон срабатывает с достаточно высокой частотой, ВПСП могут перекрываться и суммироваться. Если достаточное количество ВПСП перекрывается, суммарный ВПСП может достичь порога для инициирования потенциала действия. Этот процесс известен как суммирование и может служить фильтром верхних частот для нейронов.
С другой стороны, пресинаптический нейрон, высвобождающий тормозящий нейротрансмиттер, такой как ГАМК, может вызвать ингибирующий постсинаптический потенциал (IPSP) в постсинаптическом нейроне, отводящий мембранный потенциал дальше от порогового значения, снижая его возбудимость и затрудняя запуск нейроном потенциала действия. Если IPSP перекрывается с EPSP, IPSP может во многих случаях предотвратить запуск нейроном потенциала действия. Таким образом, выходной сигнал нейрона может зависеть от входа множества разных нейронов, каждый из которых может иметь разную степень влияния, в зависимости от силы и типа синапса с этим нейроном. Джон Кэрью Эклс провел несколько важных ранних экспериментов по синаптической интеграции, за которые он получил Нобелевскую премию по физиологии и медицине в 1963 году.
Объемная передача
Когда нейромедиатор высвобождается в синапсе, он достигает своей максимальной концентрации в узком пространстве синаптической щели, но некоторая его часть обязательно диффундирует, прежде чем реабсорбируется или разрушается. Если он диффундирует прочь, он может активировать рецепторы, расположенные либо в других синапсах, либо на мембране вдали от любого синапса. Внесинаптическая активность нейротрансмиттера известна как объемная передача. Хорошо известно, что такие эффекты происходят в некоторой степени, но их функциональное значение долгое время оставалось предметом споров.
Недавняя работа показывает, что передача объема может быть преобладающим способом взаимодействия для некоторых специальных типов нейронов. В коре головного мозга млекопитающих класс нейронов, называемых нейроглиаформными клетками, может подавлять другие близлежащие нейроны коры, высвобождая нейромедиатор ГАМК во внеклеточное пространство. По тому же принципу ГАМК, высвобождаемая из нейроглиаформных клеток во внеклеточное пространство, также действует на окружающие астроциты, определяя роль передачи объема в контроле гомеостаза ионов и нейротрансмиттеров. Примерно 78% бутонов нейроглиаформных клеток не образуют классических синапсов. Это может быть первым убедительным примером химического взаимодействия нейронов там, где нет классических синапсов.
Связь с электрическими синапсами
электрический синапс является электрически проводящим связь между двумя примыкающими нейронами, которая образуется в узком промежутке между пре- и постсинаптическими клетками, известном как щелевое соединение. В щелевых соединениях клетки приближаются друг к другу на расстояние примерно 3,5 нм, а не на расстояние от 20 до 40 нм, которое разделяет клетки в химических синапсах. В отличие от химических синапсов, постсинаптический потенциал в электрических синапсах вызывается не открытием ионных каналов химическими передатчиками, а скорее прямым электрическим взаимодействием между обоими нейронами. Электрические синапсы быстрее химических. Электрические синапсы обнаруживаются по всей нервной системе, в том числе в сетчатке, ретикулярном ядре таламуса, неокортексе и в гиппокампе. В то время как химические синапсы обнаруживаются между возбуждающими и тормозящими нейронами, электрические синапсы чаще всего встречаются между меньшими локальными тормозящими нейронами. Электрические синапсы могут существовать между двумя аксонами, двумя дендритами или между аксоном и дендритом. У некоторых рыб и амфибий электрические синапсы могут быть обнаружены в том же конце химического синапса, что и в клетках Маутнера.
Действие лекарств
Одна из наиболее важных особенностей химических синапсов заключается в том, что они являются местом действия большинства психоактивных веществ. На синапсы влияют такие наркотики, как кураре, стрихнин, кокаин, морфин, алкоголь, ЛСД и многие другие. Эти препараты по-разному влияют на синаптическую функцию и часто ограничиваются синапсами, которые используют конкретный нейромедиатор. Например, кураре — это яд, не позволяющий ацетилхолину деполяризовать постсинаптическую мембрану, вызывая паралич. Стрихнин блокирует ингибирующие эффекты нейротрансмиттера глицина, который заставляет организм улавливать и реагировать на более слабые и ранее игнорированные раздражители, что приводит к неконтролируемым мышечным спазмам. Морфин действует на синапсы, которые используют нейротрансмиттеры эндорфина, а алкоголь усиливает ингибирующие эффекты нейромедиатора ГАМК. LSD воздействует на синапсы, которые используют нейротрансмиттер серотонин. Кокаин блокирует обратный захват дофамина и, следовательно, усиливает его эффекты.
История и этимология
В 1950-х годах Бернард Кац и Пол Фатт наблюдали спонтанные миниатюрные синаптические токи в нервно-мышечном соединении лягушки . Основываясь на этих наблюдениях, они разработали «квантовую гипотезу», которая лежит в основе нашего нынешнего понимания высвобождения нейромедиаторов как экзоцитоза, за которое Кац получил Нобелевскую премию по физиологии и медицине в 1970. В конце 1960-х Рикардо Миледи и Кац выдвинули гипотезу о том, что приток ионов кальция, вызванный деполяризацией, запускает экзоцитоз.
Сэр Чарльз Скотт Шеррингтонин придумал слово «синапс» и история этого слова была дана Шеррингтоном в письме, которое он написал Джону Фултону:
«Я почувствовал потребность в каком-то имени, чтобы называть соединение между нервной клеткой и нервной клеткой… Я предложил использовать» синдесм «… Он [сэр Майкл Фостер ] проконсультировался по этому поводу со своим другом троицы Верролом, еврипидовым ученым, и Верралл предложил «синапс» (от греч. «застежка») »- Чарльз Скотт Шеррингтон
См. также
- Акклиматизация (нейроны)
- Нейробиология
- Ленточный синапс
Примечания
См. ences
- (2007). (9-е изд.). Бостон, Массачусетс: Образование Пирсона. ISBN 978-0-205-59389-7 . CS1 maint: ref = harv (link )
- Кандел, Эрик Р. ; Шварц, James H.; Jessell, Thomas M. (2000). Principles of Neural Science (4 ed.). New York: McGraw-Hill. ISBN 978-0 -8385-7701-1 . CS1 maint: ref = harv (ссылка )
- Ллинас Р., Сугимори М., Саймон С.М. (апрель 1982). «Передача пресинаптической шиповидной деполяризация в гигантском синапсе кальмара «. Proc. Natl. Acad. Sci. USA 79 (7): 2415–9. Bibcode : 1982PNAS… 79.2415L. doi : 10.1073 / pnas.79.7.2415. PMC 346205. PMID 6954549.
- Ллинас Р., Стейнберг И.З., Уолтон К. (1981). «Взаимосвязь между пресинаптическим кальциевым током и постсинаптическим потенциалом в гигантском синапсе кальмара». Биофизический журнал. 33 (3): 323–352. Bibcode : 1981BpJ…. 33..323L. doi : 10.1016 / S0006-3495 (81) 84899-0. PMC 1327434. PMID 62 61850.
- Медведь, Марк Ф.; Коннорс, Барри У.; Парадизо, Майкл А. (2001). Неврология: изучение мозга. Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-3944-3 .
- Хормузди, SG; Филиппов, М.А. Mitropoulou, G; Моньер, H; Бруззоне, Р. (март 2004 г.). «Электрические синапсы: динамическая сигнальная система, которая формирует активность нейронных сетей». Biochim Biophys Acta. 1662 (1–2): 113–137. doi : 10.1016 / j.bbamem.2003.10.023. PMID 15033583. CS1 maint: ref = harv (ссылка )
- Карп, Джеральд (2005). Клеточная и молекулярная биология: концепции и эксперименты (4-е изд.). Хобокен, штат Нью-Джерси: John Wiley Sons. ISBN 978-0-471-46580-5 .
- Николс, JG; Мартин, Арканзас; Уоллес, BG; Fuchs, PA (2001). From Neuron to Brain (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-439- 3 .
Внешние ссылки
Слушайте эту статью ![]() Этот аудиофайл был создан на основе редакции этой статьи от 19.06.2005 и не отражает последующих правок. (
Этот аудиофайл был создан на основе редакции этой статьи от 19.06.2005 и не отражает последующих правок. (
- Аудиопомощь
- Другие устные статьи
)
- Обзор Synapse для детей
- Synapses Biologymad.com (2004)
- Synapse — Cell Centered Database
- Атлас ультраструктурной нейроцитологии Собрана большая галерея изображений с электронного микроскопа Лаборатория синапсов и других нейронных структур Кристен Харрис.
Активность на конце аксона: нейрон A передает сигнал на конце аксона нейрону B (принимает). Функции: 1. Митохондрия. 2. Синаптический пузырек с нейротрансмиттеры. 3. Авторецептор. 4. Синапс с выпущенным нейротрансмиттером (серотонин ). 5.Постсинаптические рецепторы, активируемые нейротрансмиттером (индукция постсинаптического потенциала). 6. Кальциевый канал. 7. Экзоцитоз пузырька. 8. Повторно захваченный нейротрансмиттер.
Терминалы Axon (также называемый синаптические бутоны, терминальные бутоны, или же конечности) являются дистальными окончаниями телодендрия (филиалы) аксон. Аксон, также называемый нервным волокном, представляет собой длинный тонкий выступ нервной клетки или нейрон, который проводит электрические импульсы, называемые потенциалы действия вдали от нейрона Тело клетки, или сома, чтобы передавать эти импульсы другим нейронам, мышечным клеткам или железам.
Нейроны связаны между собой сложным образом и используют электрохимические сигналы и нейротрансмиттер химические вещества для передачи импульсов от одного нейрона к другому; терминалы аксонов отделены от соседних нейронов небольшой щелью, называемой синапс, по которому посылаются импульсы. Конец аксона и нейрон, от которого он исходит, иногда называют «пресинаптическим» нейроном.
Высвобождение нервного импульса
Нейротрансмиттеры упакованы в синаптические везикулы этот кластер под терминальной мембраной аксона на пресинаптической стороне синапса. Аксональные окончания специализируются на высвобождении нейромедиаторов пресинаптической клетки.[1] Терминалы выпускают передающие вещества в щель, называемую синаптическая щель между терминалами и дендритами следующего нейрона. Информацию получают связанные с ней дендритные рецепторы постсинаптической клетки. Нейроны не касаются друг друга, а общаются через синапс.[2]
Пакеты молекул нейротрансмиттера (везикулы) создаются внутри нейрона, затем перемещаются по аксону к дистальному окончанию аксона, где они находятся. состыкованный. Затем ионы кальция запускают биохимический каскад, в результате которого везикулы сливаются с пресинаптической мембраной и высвобождают свое содержимое в синаптическую щель в течение 180 секунд.мкс поступления кальция.[3] Запущенные связыванием ионов кальция, белки синаптических везикул начинают раздвигаться, что приводит к созданию поры слияния. Наличие пор позволяет высвобождать нейромедиатор в синаптическую щель.[4][5] Процесс, происходящий на окончании аксона: экзоцитоз, который клетка использует для выделения секреторной пузырьки вне клеточная мембрана. Эти мембраносвязанные везикулы содержат растворимые белки секретироваться во внеклеточную среду, а также мембранные белки и липиды которые отправляются, чтобы стать компонентами клеточной мембраны. Экзоцитоз в нейронах химические синапсы это Ca2+ запускается и обслуживает межнейронную передачу сигналов.[6]
Картографическая деятельность
| Нейрон |
|---|
|
Дендрит Сома Аксон Ядро Узел Аксон терминал Шванновская ячейка Миелиновой оболочки |
Доктор Уэйд Регер, а Профессор из Нейробиология в Гарвардская медицинская школа с Отделение нейробиологии, разработали метод физиологического наблюдения синаптической активности, происходящей в головном мозге. Краситель изменяет флуоресцентные свойства при присоединении к кальцию. С помощью флуоресцентная микроскопия методы определения уровня кальция, и, следовательно, приток кальция в пресинаптический нейрон.[7] Лаборатория Регера специализируется на пресинаптической кальциевой динамике, которая происходит на окончаниях аксонов. Регер изучает значение кальций Ca2+ поскольку это влияет на синаптическую силу.[8][9] Изучая физиологический процесс и механизмы, мы получаем дальнейшее понимание неврологических расстройств, таких как: эпилепсия, шизофрения и сильное депрессивное расстройство, а также объем памяти и учусь.[10][11]
Смотрите также
- Эндоплазматический ретикулум
- аппарат Гольджи
- Мицелла
- Мембранная нанотрубка
- Эндоцитоз
- Транспортер везикулярных моноаминов
Рекомендации
- ^ «Аксон Терминал». Медицинский словарь онлайн. Архивировано из оригинал на 2016-03-04. Получено 6 февраля, 2013.
- ^ Фостер, Салли. «Аксонный терминал — синаптический пузырек — нейротрансмиттер». Получено 6 февраля, 2013.[самостоятельно опубликованный источник? ][ненадежный медицинский источник? ]
- ^ Ллинас Р., Стейнберг И.З., Уолтон К. (март 1981 г.). «Связь между пресинаптическим кальциевым током и постсинаптическим потенциалом в гигантском синапсе кальмаров». Биофизический журнал. 33 (3): 323–51. Bibcode:1981BpJ …. 33..323L. Дои:10.1016 / S0006-3495 (81) 84899-0. ЧВК 1327434. PMID 6261850.
- ^ Карлсон, 2007, стр.56.[требуется проверка ]
- ^ Чудлер Э.Х. (24 ноября 2011 г.). «Неврология для детей. Нейротрансмиттеры и нейроактивные пептиды». В архиве с оригинала 18 декабря 2008 г.. Получено 6 февраля, 2013.[самостоятельно опубликованный источник? ][ненадежный медицинский источник? ]
- ^ Südhof TC, Rizo J (декабрь 2011 г.). «Экзоцитоз синаптических везикул». Перспективы Колд-Спринг-Харбор в биологии. 3 (12): a005637 – a005637. Дои:10.1101 / cshperspect.a005637. ЧВК 3225952. PMID 22026965.
- ^ Заубер К. «Фокус 20 октября — Нейробиология ВИЗУАЛИЗАЦИЯ СИНАПТИЧЕСКОЙ СВЯЗИ». Архивировано из оригинал на 2006-09-01. Получено 3 июля, 2013.
- ^ Регер В. (1999–2008). «Уэйд Регер, доктор философии» Архивировано из оригинал 18 февраля 2010 г.. Получено 3 июля, 2013.[самостоятельно опубликованный источник? ]
- ^ Президент и научные сотрудники Гарвардского колледжа (2008 г.). «Отделение нейробиологии Гарвардской медицинской школы». Архивировано из оригинал 20 декабря 2008 г.. Получено 3 июля, 2013.
- ^ «NINDS объявляет новых лауреатов премии исследователя нейробиологии Джавитса» (Пресс-релиз). Национальный институт неврологических заболеваний и инсульта. 4 мая 2005 г. В архиве из оригинала 17 января 2009 г.. Получено 6 февраля, 2013.
- ^ «Scholar Awards». Благотворительный фонд нейробиологии Макнайта. Архивировано из оригинал на 2004-05-08. Получено 3 июля, 2013.
дальнейшее чтение
- Cragg SJ, Greenfield SA (август 1997 г.). «Дифференциальный ауторецепторный контроль соматодендритного и терминального высвобождения дофамина аксона в черной субстанции, вентральной тегментальной области и полосатом теле». Журнал неврологии. 17 (15): 5738–46. Дои:10.1523 / JNEUROSCI.17-15-05738.1997. ЧВК 6573186. PMID 9221772.
- Vaquero CF, de la Villa P (октябрь 1999 г.). «Локализация рецепторов ГАМК (С) на конце аксона стержневых биполярных клеток сетчатки мыши». Нейробиологические исследования. 35 (1): 1–7. Дои:10.1016 / S0168-0102 (99) 00050-4. PMID 10555158.
- Роффлер-Тарлов С., Беарт П.М., О’Горман С., Сидман Р.Л. (май 1979 г.). «Нейрохимические и морфологические последствия дегенерации терминала аксона в глубоких ядрах мозжечка мышей с наследственной дегенерацией клеток Пуркинье». Исследование мозга. 168 (1): 75–95. Дои:10.1016 / 0006-8993 (79) 90129-X. PMID 455087.
- Яги Т., Канеко А. (февраль 1988 г.). «Терминал аксона горизонтальных клеток сетчатки золотой рыбки: низкая проводимость мембраны, измеренная в одиночных препаратах, и ее влияние на передачу сигнала от сомы». Журнал нейрофизиологии. 59 (2): 482–94. Дои:10.1152 / jn.1988.59.2.482. PMID 3351572.
- LTP способствует образованию множественных шипованных синапсов между одним концом аксона и дендритом.[1]
- ^ Тони Н., Букс П.А., Никоненко И., Брон ЧР, Мюллер Д. (ноябрь 1999 г.). «LTP способствует образованию множественных синапсов позвоночника между одним концом аксона и дендритом». Природа. 402 (6760): 421–5. Bibcode:1999Натура 402..421Т. Дои:10.1038/46574. PMID 10586883.