Skip to content
- Главная
- БашГМУ
- Гистология
- Периферическая нервная система
Башкирский государственный медицинский университет
Периферическая нервная система
тест к зачету
- 1
- 2
- 3
- 4
- 5
- 6
- 7
- 8
- 9
- 10
- 11
- 12
- 13
- 14
- 15
- 16
- 17
- 18
- 19
- 20
- 21
- 22
- 23
- 24
- 25
- 26
- 27
- 28
- 29
- 30
- 31
- 32
- 33
- 34
- 35
- 36
- 37
- 38
- 39
- 40
- 41
- 42
- 43
- 44
- 45
- 46
- 47
- 48
- 49
- 50
- 17
- 18
- 19
- 20
- 21
- 22
- 23
- 24
- 25
- 26
- 27
- 28
- 29
- 30
- 31
- 32
- 33
- 34
- 35
- 36
- 37
- 38
- 39
- 40
- 41
- 42
- 43
- 44
- 45
- 46
- 47
- 48
- 49
- 50
- 51
- 52
- 53
- 54
- 55
-
Задание 1 из ——
Укажите основные функции скелета человека
- ———-
- ———-
- ———-
- ———-
Перед отправкой формы внимательно прочитайте то, что находится под каждым из них. В противном случае, ваше обращение может не дойти до нас.
-
Особенности строения и функции аксонов, аксонный транспорт.
Аксон
(греч.
ἀξον —
ось) — нейрит, осевой цилиндр, отросток
нервной клетки, по которому нервные
импульсы идут от тела клетки
(сомы) к иннервируемым органам и другим
нервным клеткам.
Нейрон состоит из
одного аксона, тела и нескольких
дендритов,
в зависимости от числа которых нервные
клетки делятся на униполярные, биполярные,
мультиполярные. Передача нервного
импульса происходит от дендритов (или
от тела клетки) к аксону, а затем
сгенерированный потенциал действия от
начального сегмента аксона передается
назад к дендритам . Если аксон в нервной
ткани соединяется с телом следующей
нервной клетки, такой контакт называется
аксо-соматическим, с дендритами —
аксо-дендритический, с другим аксоном —
аксо-аксональный (редкий тип соединения,
встречается в ЦНС).
В месте соединения
аксона с телом нейрона у наиболее крупных
пирамидных клеток 5-ого слоя коры
находится аксонный
холмик.
Ранее предполагалось, что здесь происходит
преобразование постсинаптического
потенциала нейрона в нервные импульсы,
но экспериментальные данные это не
подтвердили. Регистрация электрических
потенциалов выявила, что нервный импульс
генерируется в самом аксоне, а именно
в начальном сегменте на расстоянии ~50
мкм от тела нейрона. Для генерации
потенциала действия в начальном сегменте
аксона требуется повышенная концентрация
натриевых каналов (до ста раз по сравнению
с телом нейрона[3]).
Питание и рост
аксона зависят от тела нейрона: при
перерезке аксона его периферическая
часть отмирает, а центральная сохраняет
жизнеспособность. При диаметре в
несколько микронов
длина аксона может достигать у крупных
животных 1 метра и более (например,
аксоны, идущие от нейронов спинного
мозга
в конечности).
У многих животных (кальмаров,
рыб,
кольчатых
червей,
форонид,
ракообразных)
встречаются гигантские аксоны толщиной
в сотни мкм (у кальмаров — до 2—3 мм).
Обычно такие аксоны отвечают за проведение
сигналов к мышцам. обеспечивающим
«реакцию бегства» (втягивание в норку,
быстрое плавание и др.). При прочих равных
условиях с увеличением диаметра аксона
увеличивается скорость проведения по
нему нервных импульсов.
В протоплазме
аксона — аксоплазме — имеются
тончайшие волоконца — нейрофибриллы,
а также микротрубочки,
митохондрии
и агранулярная (гладкая) эндоплазматическая
сеть.
В зависимости от того, покрыты ли аксоны
миелиновой
(мякотной) оболочкой или лишены её, они
образуют мякотные или безмякотные
нервные
волокна.
Миелиновая оболочка
аксонов имеется только у позвоночных.
Её образуют «накручивающиеся» на аксон
специальные шванновские
клетки,
между которыми остаются свободные от
миелиновой оболочки участки —
перехваты
Ранвье.
Только на перехватах присутствуют
потенциал-зависимые натриевые каналы
и заново возникает потенциал
действия.
При этом нервный импульс распространяется
по миелинизированным волокнам ступенчато,
что в несколько раз повышает скорость
его распространения.
Концевые участки
аксона — терминали — ветвятся и
контактируют с другими нервными,
мышечными или железистыми клетками. На
конце аксона находится синаптическое
окончание —
концевой участок терминали, контактирующий
с клеткой-мишенью. Вместе с постсинаптической
мембраной клетки-мишени синаптическое
окончание образует синапс.
Через синапсы
передаётся возбуждение.
Специфическая
функция аксона — проведение потенциала
действия
от тела клетки к другим клеткам или
периферическим органам. Другая его
функция — аксонный
транспорт
веществ.
Помимо своей
специфической функции в качестве
проводника потенциалов
действия
аксон
является каналом для транспорта веществ.
Аксонный транспорт
— это перемещение веществ по аксону
. Белки, синтезированные в теле клетки,
синаптические
медиаторные вещества
и низкомолекулярные соединения
перемещаются по аксону вместе с клеточными
органеллами, в частности, митохондриями
. Для большинства веществ и органелл
обнаружен также транспорт в обратном
направлении. Вирусы
и токсины
могут проникать в аксон на его периферии
и перемещаться по нему. Аксонный транспорт
— активный процесс.
Аксонный транспорт
зависит от достаточного снабжения
энергией, при снижении уровня АТФ вдвое
аксонный транспорт блокируется, при
возобновлении доступа энергии —
возобновляется.
Белки цитоскелета
доставляются из тела клетки, двигаясь
по аксону со скоростью от 1 до 5 мм в
сутки. Это медленный
аксонный транспорт
(похожий на него транспорт имеется и в
дендритах). Многие ферменты и другие
белки цитозоля также переносятся при
помощи этого типа транспорта.
Нецитозольные
материалы, которые необходимы в синапсе,
такие как секретируемые белки и
мембраносвязанные молекулы, двигаются
по аксону с гораздо большей скоростью.
Эти вещества переносятся из места их
синтеза, эндоплазматического ретикулума,
к аппарату Гольджи, который часто
располагается у основания аксона. Затем
эти молекулы, упакованные в мембранные
пузырьки, переносятся вдоль
рельсов-микротрубочек путем быстрого
аксонного транспорта
со скоростью до 400 мм в сутки. Таким
образом по аксону транспортируются
митохондрии, различные белки, включая
нейропептиды (нейромедиаторы пептидной
природы), непептидные нейромедиаторы.
Транспорт материалов
от тела нейрона к синапсу называется
антероградным,
а в обратном направлении — ретроградным.
Транспорт по аксону
на большие расстояния происходит с
участием микротрубочек. Микротрубочки
в аксоне обладают присущей им полярностью
и ориентированны быстрорастущим
(плюс-)концом к синапсу, а медленнорастущим
(минус-) — к телу нейрона. Белки-моторы
аксонного транспорта принадлежат к
кинезиновому и динеиновому суперсемействам.
Кинезины являются,
в основном, плюс-концевыми моторными
белками, транспортирующими такие грузы,
как предшественники синаптических
везикул и мембранные органеллы. Этот
транспорт идет в направлению к синапсу
(антероградно). Цитоплазматические
динеины — это минус-концевые моторные
белки, транспортирующие нейротрофные
сигналы, эндосомы и другие грузы
ретроградно к телу нейрона. Ретроградный
транспорт осуществляется динеинами не
эксклюзивно: обнаружены несколько
кинезинов, перемещающихся в ретроградном
направлении.
-
Миенилизированные
и немиенилизированные волокна. Процесс
миелинизации.
Миелин
(в некоторых изданиях употребляется
некорректная теперь форма миэлин) —
вещество, образующее миелиновую
оболочку
нервных
волокон.
Миелиновая
оболочка —
электроизолирующая оболочка, покрывающая
аксоны
многих нейронов. Миелиновую оболочку
образуют глиальные клетки: в периферической
нервной системе — Шванновские
клетки,
в центральной нервной системе —
олигодендроциты.
Миелиновая оболочка формируется из
плоского выроста тела глиальной клетки,
многократно оборачивающего аксон
подобно изоляционной ленте. Цитоплазма
в выросте практически отсутствует, в
результате чего миелиновая оболочка
представляет собой, по сути, множество
слоёв клеточной мембраны.
Миелин прерывается
только в области перехватов Ранвье,
которые встречаются через правильные
промежутки длиной примерно 1 мм. В
связи с тем, что ионные токи не могут
проходить сквозь миелин, вход и выход
ионов осуществляется лишь в области
перехватов. Это ведёт к увеличению
скорости проведения нервного импульса.
Таким образом, по миелинизированным
волокнам импульс проводится приблизительно
в 5—10 раз быстрее, чем по немиелинизированным.
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Безмиелиновые
нервные волокна находятся преимущественно
в составе вегетативной нервной системы.
Клетки олигодендроглии оболочек
безмиелиновых нервных волокон,
располагаясь плотно, образуют тяжи, в
которых на определенном расстоянии
друг от друга видны овальные ядра. В
нервных волокнах внутренних органов,
как правило, в таком тяже располагается
не один, а несколько (10-20) осевых цилиндров,
принадлежащих различным нейронам. Они
могут, покидая одно волокно, переходить
в смежное, такие волокна, содержащие
несколько осевых цилиндров, называются
волокнами кабельного типа. При электронной
микроскопии безмиелиновых нервных
волокон видно, что по мере погружения
осевых цилиндров в тяж леммоцитов
последние одевают их как муфтой.
Оболочки леммоцитов
при этом прогибаются, плотно охватывают
осевые цилиндры и, смыкаясь над ними,
образуют глубокие складки, на дне которых
и располагаются отдельные осевые
цилиндры. Сближенные в области складки
участки оболочки нейролеммоцита образуют
сдвоенную мембрану — мезаксон, на которой
как бы подвешен осевой цилиндр. Оболочки
нейролеммоцитов очень тонкие, поэтому
ни мезаксона, ни границ этих клеток под
световым микроскопом нельзя рассмотреть,
и оболочка безмиелиновых нервных волокон
в этих условиях выявляется как однородный
тяж цитоплазмы, «одевающий» осевые
цилиндры. С поверхности каждое нервное
волокно покрыто базальной мембраной.
-
Миелиновые
– более толстые, т.к. содержат миелин-
находятся внутри под оболочкой; входят
в состав соматической нервной системы.
Миелин через несколько участков резко
истончается, образуя таким образом
перехват. Он называется перехват Ранвье.
Отросток между перехватами называется
межузловой сигмент. Миелин на подобии
жира. -
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
А. Нейроны
- Мультиполярные нейроны
а. Поперечный срез спинного мозга, импрегнированный нитратом серебра
(рис. 8-24). В сером веществе спинного мозга, кроме клеток нейроглии, находятся мультиполярные нервные клетки, особенно крупные в передних рогах. Это двигательные нейроны. Так как мы имеем дело со срезом, то видны только наиболее толстые, ближайшие к клетке участки отростков. Ядро нейрона имеет вид светлого пузырька. Иногда видно ядрышко. В нейроплазме располагается сеть тонких тёмноокрашенных нитей — нейрофибрилл.
б. Поперечный срез спинного мозга, окрашенный на тигроидное вещество (рис. 8-25). В перикарионах мультиполярных нейронов присутствует тигроидное вещество в виде многочисленных глыбок. Часть перикариона, обращённая к аксону, лишена тигроидного вещества. Это аксонный холмик. Аксон также не содержит тигроидного вещества.
- Псевдоуниполярные нейроны спинномозговых узлов (рис. 8-26). Узел покрыт соединительнотканной капсулой. Внутри узла находятся группы нервных клеток, между которыми проходят пучки миелиновых нервных волокон. Тела псевдоуниполярных нейронов имеют округлую форму. Местами можно видеть толстый отросток (иногда Т-образно ветвящийся), отходящий от тела клетки. Тела нервных клеток окружены клетками-сателлитами. Популяция нейронов ганглия неоднородна. Нейроны с небольшим перикарионом содержат вещество Р, соматостатин и холецистокинин, половина нейронов — глутамин. От 35 до 65% небольших нейронов содержат одновременно вещество P и глутамин. Оба эти нейромедиатора секретируются из одних и тех же терминалей в пределах пластинки II серого вещества спинного мозга.
Б. Проводники
- Миелиновые нервные волокна (расщеплённый препарат седалищного нерва, рис. 8-27) имеют вид длинных неветвящихся цилиндров, окружённых миелиновой оболочкой. Миелин интенсивно окрашен тетраоксидом осмия. Миелиновая оболочка прерывается перетяжками, или перехватами Ранвьё, разбивающими миелиновый футляр осевого цилиндра на сегменты. Каждому сегменту соответствует одна шванновская клетка. В миелиновой оболочке имеются светлые косонаправленные полосы — насечки Шмидта-Лантермана.
Рис. 8-24. Мультиполярный нейрон спинного мозга. Крупное ядро расположено в теле клетки центрально. Хорошо видно ядрышко. В цитоплазме перикариона различимы нейрофибриллы — скопления элементов цитоскелета. В перикарионе они проходят в различных направлениях, а в отростках имеют направленную ориентацию [из Stohr Р, Mdllendorf W, 1933]
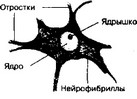
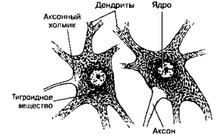
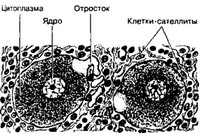 Рис. 8-25. Тигроидное вещество. Глыб- ки тигроидного вещества (субстанции Ниссля) располагаются исключительно в перикарионе и начальных отделах дендритов. Свободный от тигроидного вещества участок перикариона просматривается в том месте, где начинается аксон. Этот участок называют аксонным холмиком [из Voss H, 1957]
Рис. 8-25. Тигроидное вещество. Глыб- ки тигроидного вещества (субстанции Ниссля) располагаются исключительно в перикарионе и начальных отделах дендритов. Свободный от тигроидного вещества участок перикариона просматривается в том месте, где начинается аксон. Этот участок называют аксонным холмиком [из Voss H, 1957]
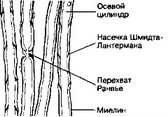 Рис. 8-26. Псевдоуниполярные нейроны спинномозгового узла. Сферической формы перикарионы содержат ядра. От тела клетки отходит сравнительно толстый отросток, который на некотором расстоянии от перикариона Т-образно разветвляется на центральную и периферическую ветви. Перикарион окружён клетками небольшой величины — клет- ками-сателлитами [из Voss Я, 1957]
Рис. 8-26. Псевдоуниполярные нейроны спинномозгового узла. Сферической формы перикарионы содержат ядра. От тела клетки отходит сравнительно толстый отросток, который на некотором расстоянии от перикариона Т-образно разветвляется на центральную и периферическую ветви. Перикарион окружён клетками небольшой величины — клет- ками-сателлитами [из Voss Я, 1957]
Рис. 8-27. Миелиновые волокна. В
центральной части волокна в виде светлой ленты проходит осевой цилиндр. Снаружи к нему прилегает миелино- вая оболочка, образованная шваннов- ской клеткой. Проходящие под углом к осевому цилиндру светлые линии в миелиновой оболочке называют насечками Шмидта-Лантермана. Участок волокна, соответствующий месту контакта соседних шванновских клеток, — перехват Ранвье [из Ком Н, 1957]
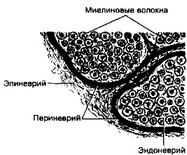
- Нервный ствол в поперечном разрезе (рис. 8-28). Нервный ствол состоит из миели- новых и безмиелиновых нервных волокон и соединительнотканных оболочек. Миелиновые нервные волокна имеют вид округлых профилей, центральная их часть занята осевым цилиндром. Миелиновая оболочка окрашена слабо, шванновская оболочка в виде тонкой линии очерчивает снаружи миелиновую оболочку. Нервные волокна собираются в пучки разного диаметра, окружённые периневрием; от периневрия между нервными волокнами отходят нежные прослойки соединительной Ткани — эндоневрий. Весь нерв с поверхности одет соединительнотканной оболочкой — эпиневрием.
В. Нервные окончания
- Двигательное нервное окончание (нервно-мышечный синапс, рис. 8-29). К параллельно расположенным мышечным волокнам подходят пучки миелиновых нервных волокон, импрегнированных нитратом серебра. Пучки постепенно распадаются на отдельные тонкие волокна, подходят к поперечнополосатым мышечным волокнам, где оканчиваются нервно-мышечными синапсами, или двигательными бляшками. В области контакта терминален нервного окончания с сарколеммой мышечного волокна образуется т.н. подошва окончания, образованная, во-первых, за счёт сарколеммы и саркоплазмы, содержащей ядра мышечного волокна (поперечная исчерченность отсутствует), и, во-вторых, за счёт скопления в этой области шванновских клеток.
- Свободное нервное окончание (рис. 8-30). Миелиновое нервное волокно теряет миелин и образует кустик. В зависимости от количества и протяжённости ветвей, кустиковидные рецепторы могут быть компактными или распространёнными.
-
 Механорецептор — тельце Пачини (рис. 8-31). В тельце различают наружную слоистую капсулу и внутреннюю колбу. Наружная капсула состоит из концентрически расположенных уплощённых фибробластов. Внутренняя колба представлена специальными клетками нейроэктодермального происхождения. В центре внутренней колбы проходит осевой цилиндр.
Механорецептор — тельце Пачини (рис. 8-31). В тельце различают наружную слоистую капсулу и внутреннюю колбу. Наружная капсула состоит из концентрически расположенных уплощённых фибробластов. Внутренняя колба представлена специальными клетками нейроэктодермального происхождения. В центре внутренней колбы проходит осевой цилиндр.
 Механорецептор — тельце Пачини (рис. 8-31). В тельце различают наружную слоистую капсулу и внутреннюю колбу. Наружная капсула состоит из концентрически расположенных уплощённых фибробластов. Внутренняя колба представлена специальными клетками нейроэктодермального происхождения. В центре внутренней колбы проходит осевой цилиндр.
Механорецептор — тельце Пачини (рис. 8-31). В тельце различают наружную слоистую капсулу и внутреннюю колбу. Наружная капсула состоит из концентрически расположенных уплощённых фибробластов. Внутренняя колба представлена специальными клетками нейроэктодермального происхождения. В центре внутренней колбы проходит осевой цилиндр.
Рис. 8-28. Нервный ствол в поперечном разрезе. Нервные волокна, окружённые эндоневрием, образуют пучки. Каждый пучок окружён соединительнотканной оболочкой — периневрием. Соединительная ткань, покрывающая нерв с поверхности, — эпиневрий [из Ком Н, 1957]
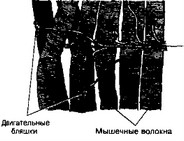

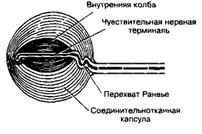 Рис. 8-30. Свободное нервное окончание. Тонкие ветвящиеся терминали афферентного волокна залегают среди клеточных элементов иннервируемой ткани [по Шмелёвой TH, 1954]
Рис. 8-30. Свободное нервное окончание. Тонкие ветвящиеся терминали афферентного волокна залегают среди клеточных элементов иннервируемой ткани [по Шмелёвой TH, 1954]
Рис. 8-31. Инкапсулированный рецептор — тельце Пачини.
Центральную часть рецептора занимает внутренняя колба, по всей длине которой проходит чувствительная нервная терминаль. Часть миелинового волокна и вся внутренняя колба окружены многослойной соединительнотканной капсулой [из Quilliam TA, Sato М, 1955]
- Мышечное веретено (рис. 8-32). Одно или несколько мышечных волокон окружены тонкой капсулой, имеющей веретеновидную форму. Мышечные волокна, лежащие под капсулой, называются интрафузальными. Миелиновые нервные волокна, проникая под капсулу, оплетают интрафузальные мышечные волокна в виде спирали. На концах интрафузальных мышечных волокон располагаются небольшие двигательные бляшки.
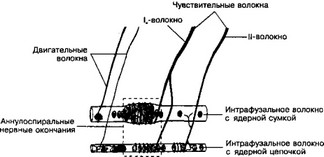
Рис. 8-32. Средняя часть двигательного веретена содержит интрафузальные мышечные волокна, к которым подходят двигательные и чувствительные нервные волокна. Терминали чувствительных волокон образуют спиральные структуры или ветвятся на поверхности интрафузальных волокон. Двигательные нервные волокна формируют нервно-мышечные синапсы [из Matthews PB, 1964]
ВОПРОСЫ
Пояснение. За каждым из перечисленных вопросов или незаконченных утверждений следуют обозначенные буквой ответы или завершения утверждений. Выберите один ответ или завершение утверждения, наиболее соответствующее каждому случаю.
- В эксперименте на эмбрионах удалили нервный гребень. Нарушено развитие всех
структур, КРОМЕ:
- чувствительных нейронов спинномозговых узлов (Б) нейронов симпатических ганглиев
- хромаффинных клеток (Г) меланоцитов
(Д) мотонейронов спинного мозга
- Клетки контактируют друг с другом и другими клетками ЦНС, содержат крупное ядро и промежуточные филаменты, состоящие из глиального фибриллярного кислого белка. О какой клетке идёт речь?
- Олигодендроцит (Б) Микроглия
- Астроцит (Г) Нейрон
(Д) Шванноеская клетка
- По аксону транспортируется всё, КРОМЕ:
- рибосом (Б) везикул
- нейромедиаторов (Г) митохондрий
(Д) белковых молекул
- В аксоне присутствует всё, КРОМЕ:
- митохондрий (Б) микротрубочек
- нейрофиламентов
(Г) тигроидного вещества (Д) везикул
- У мышей с мутацией jimpy причина дрожания и судорог — нарушение миелинизации
в центральной, но не в периферической нервной системе. С нарушением функции каких клеток связан данный дефект?
- Олигодендроцит
(Б) Астроцит плазматический
- Астроцит волокнистый (Г) Микроглия
(Д) Шванноеская клетка
- Миелиновая оболочка периферических нервных волокон образована:
- уплотнённым межклеточным веществом, содержащим белки и фосфолипиды (Б) плазматической мембраной шванновских клеток
- специализированной частью периневрия
(Г) элементами цитоскелета шванновских клеток (Д) спирально закрученной мембраной аксона
- После травматического сдавления конечности в её нервном стволе найдена дегенера
ция нервных волокон. Имеются все проявления, КРОМЕ:
- распада окончаний нервных волокон (Б) тигролиза
- разрушения миелина
(Г) гибели шванновских клеток в дистальном отрезке (Д) фагоцитоза фрагментов повреждённых нервных волокон
- Медиатор в нервно-мышечном синапсе скелетной мышцы:
- адреналин (Б) норадреналин
- дофамин (Г) ацетилхолин (Д) глицин
- Какие клетки в эпидермисе кожи вместе с терминалями афферентных волокон обра
зуют тактильные рецепторы?
- Кератиноциты
(Б) Клетки Лангерханса
- Клетки Меркеля (Г) Меланоциты
(Д) Клетки-сателлиты
- Назовите чувствительное нервное окончание, ответственное за термовосприятие:
- пластинчатое тельце Пачини (Б) осязательное тельце Майсснера
- сухожильный орган Гольджи (Г) свободное нервное окончание (Д) тельце Руффйни
Пояснение. Каждый из нижеприведённых и пронумерованных вопросов 11-23 содержит четыре варианта ответов, из которых правильными могут быть один или сразу несколько. Выберите:
А — если правильны ответы I, 2 и 3 Б — если правильны ответы I и 3 В — если правильны ответы 2 и 4 Г — если правилен ответ 4 Д —¦ если правильны ответы I, 2, 3 и 4
- Производные нервного гребня:
- нейроны ресничного ганглия
- нейроны спинномозговых узлов
- нейроны стенки желудка
- нейроны спинного мозга
- Производные нервной трубки:
- псевдоуниполярные нейроны спинномозговых узлов
- пирамидные нейроны коры больших полушарий головного мозга
- меланоциты
- эпендимоциты
- Функции астроцитов:
- выделяют вещества, поддерживающие рост аксонов
- участвуют в фагоцитозе
- участвуют в метаболизме глутамина
- изолируют рецептивные поверхности нейронов
14 Для клеток микроглии характерно:
- многочисленные лизосомы
- участие в иммунном ответе в мозге
- способность к обновлению
- участие в транспорте глюкозы
- Элементы, присутствующие в аксоне:
- митохондрии
- гранулярная эндоплазматическая сеть
- промежуточные филаменты
- цистерны комплекса Гольджи
- В мнелиновом волокне ЦНС различают:
- перехват Ранвье
- осевой цилиндр
- мезаксон
- шванновские клетки
- Какие клетки синтезируют белки миелина?
- Шванновские
- Нейроны
- Олигодендроциты
- Астроциты
- Структуры периферического нерва:
- эндоневрий
- кровеносные сосуды
- фибробласты
- нервы нервов
- Периневрий:
- тонкий слой соединительной ткани вокруг каждого нервного волокна
- содержит клетки, связанные плотными контактами
- ретикулярная ткань вокруг пучков нервных волокон
- контролирует проницаемость и поддерживает гомеостаз эндоневрия
- Какие процессы происходят после локальной компрессии периферического нерва?
- Разрушение осевых цилиндров и распад миелина на всём протяжении периферического отрезка
- Дегенерация нервных волокон на небольшом протяжении центрального отрезка
- Прорастание аксонов из центрального отрезка в периферический
- Ориентация регенерирующих аксонов по цепочкам из шванновских клеток
- Укажите нервные окончания, ответственные за механорецепторную функцию:
- тельце Пачйни
- сухожильный орган Гольджи
- мышечное веретено
- комплекс клетки Меркеля с нервной терминалью
- Какие из перечисленных нервных окончаний относят к несвободным?
- Тельце Пачини
- Тельце Майсснера
- Тельце Руффйни
- Комплекс клетки Меркеля с нервной терминалью
- Нервно-мышечный синапс:
- ацетилхолин — содержимое светлых синаптических пузырьков
- постсинаптическая мембрана — область наибольшего скопления холинорецепторов
- нервная терминаль окружена шванновской клеткой
- синаптические везикулы выходят в синаптическую щель
Потенциалы действия, развивающиеся
на плазматической мембране аксона или сомадендритного комплекса, оказывают стимулирующее
действие на внутриклеточные процессы. Это связано с влиянием на ферменты клетки
проникающих в нее ионов
Na
+
и особенно Са 2+
действующих через специальный белок калъмодулин.
Таким образом, распространяющийся ПД
влечет за собой быструю волну активации внутриклеточных процессов.
Вместе с тем внутри аксона (и других
частей нервной клетки) происходят закономерные перемещения материалов (белковых
частиц, органоидов), прямо не связанные с ПД и имеющие совсем другие скорости.
Эти перемещения материалов хорошо изучены в аксонах; здесь они получили
название аксонный транспорт.
Существует два вида аксонного транспорта:
быстрый и медленный.
Быстрый аксонный транспорт —
это, например, транспорт везикул,
митохондрий и некоторых белковых частиц от тела клетки к окончаниям аксона со
скоростью у млекопитающих 250-400 мм/сут. Он осуществляется специальным
транспортным механизмом. Этот транспорт не нарушается при отделении аксона от
тела клетки, но прекращается при разрушении внутриаксонных структур —
микротрубочек и нейрофиламентов (разрушение производится колхицином,
винбластином), а также при отсутствии в аксоне АТФ и Са 2+ В связи с
этим полагают, что механизм, осуществляющий этот быстрый транспорт, подобен
механизму скольжения нитей при мышечном сокращении (см. разд. 1.2.4).
Считают, что
нейрофиламенты
перемещаются («скользят») вдоль микротрубочек, имеющих боковые выросты,
которые, видимо, своими движениями и обеспечивают скольжение нейрофиламентов.
Энергия для этого процесса извлекается из АТФ, расщепляемого ферментативно при
объединении белковых структур нейрофиламентов и выростов микротрубочек в
присутствии Са 2+ . Один из белков играет роль АТФазы.
Транспортируемые частицы прикрепляются на нейрофиламентах и как бы перевозятся
на них. Этот процесс можно наблюдать и в выдавленной из аксона аксоплазме.
Быстрый аксонный транспорт везикул
(с медиатором для синапсов) происходит в дистальном направлении — антероградный
транспорт. Существует и обратный — ретроградный —
быстрый транспорт
лизосом, везикул, мультивезикулярных тел, возникающих в окончаниях аксона в
ходе пиноцитоза, который протекает с захватом некоторых веществ (например,
ацетилхолинэстеразы, периферических факторов, регулирующих синтез белка в соме
нейрона, а также некоторых вирусов, токсинов и пероксидазы хрена — маркёра,
используемого в экспериментах). Скорость этого транспорта ≈ 220 мм/сут (у
млекопитающих). Скорости быстрого транспорта, и антероградного и ретроградного,
не зависят от типа и диаметра аксона, хотя у пойкилотермных (холоднокровных) позвоночных
они ниже, чем у гомойотермных (теплокровных).
Медленный аксонный транспорт —
это перемещение всей массы белков
цитоплазмы (микротрубочек, нейрофиламентов, РНК, каналов, насосов и т. п.) в
дистальном направлении, создаваемое за счет интенсивных синтетических процессов
в перикарионе. Медленный аксоток обнаруживает себя при тугой перетяжке нерва
лигатурой, сдавливающей аксоны. При этом в дистальной части аксона диаметр
уменьшается, а в проксимальной — перед перетяжкой образуется вздутие — «наплыв
цитоплазмы».
Медленный аксоток движется со скоростью около 1-4 мм/сут. Он
прекращается при отделении сомы от аксона и не нарушается факторами,
разрушающими микротрубочки (колхицином, винбластином). Медленный аксонный
транспорт имеет особое значение в процессах роста и регенерации аксонов
(дендритов) и их разветвлений.
Мембранные и цитоплазматические компоненты, которые образуются в
биосинтезирующем аппарате сомы
и проксимальной части дендритов
, должны распределяться по аксону
(особенно важно их поступление в пресинаптические структуры синапсов), чтобы восполнить потерю элементов, подвергшихся высвобождению или
инактивации. Однако многие аксоны слишком длинны, чтобы материалы могли
эффективно перемещаться из сомы к синаптическим окончаниям путем простой
диффузии. Эту задачу выполняет особый механизм — аксональный транспорт.
Существует несколько его типов. Окруженные мембранами органоиды и митохондрии
транспортируются с относительно большой скоростью посредством быстрого
аксонального транспорта. Вещества, растворенные в цитоплазме (например,
белки), перемещаются с помощью медленного аксонального транспорта. У
млекопитающих быстрый аксональный транспорт обладает скоростью 400 мм/сут,
а медленный — около 1 мм/сут. Синаптические пузырьки
могут передвигаться с номощью быстрого аксонального транспорта из сомы
мотонейрона спинного мозга человека к нервно-мышечному соединению
стопы примерно за 2,5 сут. Сравним: доставка на такое же расстояние
многих растворимых белков происходит примерно за 3 г.
Для работы аксонального транспорта требуются затрата метаболической
энергии и присутствие внутриклеточного Са2+. Элементы цитоскелета (точнее,
микротрубочки) создают систему направляющих тяжей, вдоль которых передвигаются
окруженные мембранами органоиды (
рис. 32.13). Эти органоиды прикрепляются к микротрубочкам аналогично тому, как это
происходит между толстыми и тонкими филаментами
волокон скелетных мышц
; движение органоидов вдоль микротрубочек запускается ионами Са2+.
Аксональный транспорт осуществляется в двух направлениях. Транспорт от
сомы к аксональным терминалям, называемый антероградным аксональным транспортом
(
рис. 32.14
, а), восполняет в пресинаптических окончаниях запас синаптических пузырьков
и ферментов, ответственных за синтез нейромедиатора. Транспорт в
противоположном направлении — ретроградный аксональный транспорт
(
рис. 32.14
, б), возвращает опустошенные синаптические пузырьки в сому, где эти
мембранные структуры деградируются лизосомами
.
Посредством аксонального транспорта по периферическим нервам
распространяются некоторые вирусы
и токсины
. Так, вирус, который может вызывать ветряную оспу
(
varicella-zoster virus), проникает в клетки спинальных ганглиев
. Там он пребывает в неактивной форме иногда в течение многих лет, пока не
изменится иммунный статус человека. Тогда вирус может транспортироваться по
сенсорным аксонам
к коже, и в дерматомах
соответствующих спинальных нервов
возникают болезненные высыпания —
В
нейроне, как и в других клетках организма, постоянно происходят процессы
распада молекул, органоидов, других компонентов клетки. Их необходимо постоянно
обновлять. Нейроплазматический транспорт важен для обеспечения электрических и
неэлектрических функций нейрона, для осуществления обратной связи между
отростками и телом нейрона. При повреждении нервов необходима регенерация
поврежденных участков и восстановление иннервации органов.
Разнообразные вещества транспортируются
по отросткам нейрона с разной скоростью, в разных направлениях и с
использованием разных механизмов транспорта. Выделяют два основных вида
транспорта: прямой (антероградный) – от тела клетки по отросткам к их периферии
и обратный (ретроградный) – по отросткам нейрона к телу клетки (табл. 1).
Табл. 1
Основные компоненты аксонного и дендритного транспорта в нейронах
позвоночных (по данным разных авторов)
|
Компоненты |
Скорость мм/сутки |
Что |
Морфологический |
|
Прямой |
|||
|
Быстрый (Fast) |
|||
| I |
200- 500 |
Медиаторы и их предшественники, ферменты синтеза медиаторов, белки плазматической мембраны, мембранные органоиды, нейрогормоны, |
Синаптические пузырьки, цистерны гладкого ретикулума, нейросекреторные гранулы, цитоскелетная сеть |
|
Промежуточный |
|||
| II |
50 — 100 |
Белки митохондрий, липиды мембран |
Митохондрии, цитоскелет |
| III | 15 |
Миозиновые белки, |
Цитоскелет |
|
Медленный (Slow) |
|||
| IV SCb | 2- 4 |
Актин, клатрин, актинсвязывающие белки, ферменты метаболизма нейрона, белки аксоплазмы |
|
| V SCa | 0,2- 1 |
Белки нейрофиламентов, тубулин и фрагменты микротрубочек, ферменты аксоплазмы |
Цитоскелет (микротрубочки, микро- и нейрофиламенты), микротрабекулярная сеть |
|
Прямой |
|||
| I D | 200- 400 |
Белки постсинапса, рецепторные комплексы, белки цитоплазмы и мембран дендрита и шипиков |
Цитоскелет, гладкий ретикулум, транспортные пузырьки |
|
Обратный |
|||
| I R | 100- 300 |
Отработанные лизосомы и митохондрии, ростовые и трофические факторы, вирусы. |
Мультивезикулярные и мультиламеллярные тела, цитоскелет, эндосомы |
В
осуществлении транспортных процессов в нейроне участвуют пять групп «моторных» белков, тесно связанных с цитоскелетной сетью. В их состав входят такие белки
как кинезины, денеины и миозины.
В осуществлении транспортных процессов в
нейроне участвуют пять групп т.н. «моторных» молекул (Рис. xx).
1-3 Группа. Кинезины
В составе этой группы выделяют три типа кинезиновых белков.
1. Группа. Конвекционный кинезин ( kinesin
— I
или
KIF
-5).
Он был идентифицирован в нервной системе головоногих моллюсков и
млекопитающих в 1985 году, а затем и в клетках других животных, включая низших
эукариот. Он тесно связан с микротрубочками и является одним из самых главных
транспортных белков клетки, осуществляя транспорт материалов (cargo) вдоль микротрубочек по направлению к
ее плюс концу. С его помощью транспортируются в отростках нейронов митохондрии,
лизосомы, цистерны эндоплазматического ретикулума, синаптические пузырьки, а
также ряд немембранных комполнентов клетки (молекулы и-РНК, белки и фибриллы нейрофиламентов).
Состоит молекула кинезина -1 из двух
тяжелых и двух легких полипептидных цепей.
Из тяжелых и легких цепей каждая кодируются тремя генами. Легкие и
тяжелые цепи могут комбинироваться в различных сочетаниях и, как полагают,
могут, таким образом, формировать различные разновидности молекул кинезина – I,
транспортирующие при этом разные
компоненты внутри клетки.
2.Группа. Гетеродимерный
кинезин, (кинезин — II
, kinesin-II, KIF – 3C).
Свое название он получил из-за наличия
трех моторных доменов в структуре молекулы. В нервных и сенсорных клетках
позвоночных и беспозвоночных животных (например: в фоторецепторах позвоночных
или в хеморецепторных клетках С. elegans) этот белок связан с работой ресничек и
жгутиков, осуществляя транспорт крупных молекулярных комплексов вдоль их аксонемальной оси (IFT – intraflagellar transport)
В аксонах нервных клеток он выполняет транспортную функцию, перемещая синаптические пузырьки и ферментные комплексы (холинэстеразу), участвующие в
работе синапсов.
Одной из форм кинезина II типа является т.н. гомодимерный кинезин
(Osm 3, KIF-17) Найден только у многоклеточных
(метазоиных) животных. Также как и гетеродимерный кинезин II, он является важнейшей составляющей
ресничек хеморецептивных клеток. В нейронах ЦНС млекопитающих эта форма кинезина участвует в транспорте по дендритам пузырьков, содержащих NMDA – синаптические рецепторы. Участие гомодимерного кинезина в IFT — транспорте обсуждается.
3 Группа. Мономерный кинезин
(UNC
-104,
KIF
-1A, Klp-53D, kinesin-73)
Эта форма транспортных белков была
обнаружена в нервной системе C . elegans, где ее мутантная форма вызывала
паралич транспорта синаптических пузырьков по аксонам моторных нейронов.
Особенностью этой транспортной молекулы является преобладающая мономерная форма
этого белка, тогда как другие формы кинезина (как отмечено выше, являются димерами или тетрамерами). Обнаруженный у многих животных (C. elegans – Unc104, дрозофила – Klp53 D ,
kinesin -73
мышь – KIF -1А,
KIF -1В,
человек — GAKIN) он
принимает участие в транспорте синаптических пузырьков, мембранных белков, связанных с формированием клеточных
контактов.
Показано, что в результате
альтернативного сплайсинга гена KIF -1B кинезина образуются две изоформы: KIF-1 Bα, участвующая в транспорте по отросткам
митохондрий и KIF -1Bβ, транспортирующая синаптические
пузырьки в аксонную терминаль.
Еще раз необходимо подчеркнуть, что все
формы кинезинов участвуют в транспорте к плюс концу микротрубочек
(антероградный, прямой транспорт)
Табдица. Некоторые молекулярные и
функциональные характеристики кинезинов в нервной ткани (по N. Hirokava, 1997)
|
Тип молекулы |
Мол.вес |
Вторичная структура |
Направление транспорта и скорость |
Специфичность экспрессии |
Транспортируемый материал |
| KIF -1А | 192 | мономер | + конец, 1,5 мкм/сек | нейроспецифичен |
Предшественники синаптических пузырьков |
| KIF -1В | 130 | мономер | + конец, 0,66 мкм/сек | повсеместно | митохондрии |
| KIF 2 | 81 | гомодимер | + конец, 0,47 мкм/сек | Пузырьки, отделяющиеся от предсшественников син. пузырьков | |
| KIF3A | 80 | Гетеродимер с KIF3B | +конец, 0, 3 мкм/сек |
Пузырьки (90-180нм), от предшественников |
|
| KIF3B | 85 | Гетеродимер с KIF3A | +конец, 0, 3 мкм/сек |
Обычен в нейронах, но экспремсируется повсеместно |
Пузырьки (90-180нм), от предшественников синаптических пузырьков |
| KIF4 | 140 |
Гомодимер, амино концевой моторный домен |
+ конец, 0,2 мкм/сек |
Повсеместно, но в раннем развитии, во взрослых нейронах слабо |
Пузырьки |
| KIF5 | |||||
| KIF 1C2 | 86 |
Гомодимер, карбоксил концевой моторный домен |
— конец, | Нейроспецифичен |
Мультивезикулярные тела, дендритный транспорт |
4 Группа Денеины.
Эти транспортные белки участвуют в
транспорте по микротрубочкам к ее минус-концу (ретроградный, обратный транспорт).
Присутствуют во многих транспортных процессах и движениях клеток, начиная от
митоза и заканчивая миграцией нейробластов в развивающемся мозге.
Имеет довольно сложную структуру,
представленную множеством субъединиц (цепей). Эти субъединицы взаимодействуют с
различными ассоциированными с денеином белками, которые, в свою очередь, могут
определять избирательный характер, выполняемых денеином функций в клетке.
Так, белок лиссэнцефалин -1 (Lis-1) будучи ассоциированным с денеином,
определяет его роль в митозе и движении ядра в клетках развивающегося мозга,
но не в транспорте органоидов. Мутации
или отсутствие этого белка в период раннего
развития организма (пренатальный период) вызывает серьезные нарушения в
формировании ЦНС и особенно коры полушарий, приводя в конечном итоге к– лиссэнцефалии (наследственное заболеванию
внешне выражающееся в недоразвитии или
полном отсутствии в больших полушариях
извилин и борозд).
5 Группа. Миозины (myosin-Vs).
Этот транспортный белок был впервые идентифицирован биохимически в мозге
позвоночных как «миозиноподобный калмодулин связывающий белок». От мышечного
миозина он отличается большой длинной шарнирной части молекулы, которая имеет
дополнительную легкую цепь и присоединенных к ней пять молекул калмодулина – Са+2
связывающего белка.
Миозин V
широко задействован у позвоночных и беспозвоночных животных в транспортных
процессах в нервных клетках. В основном он участвует в обратном транспорте
мембранных пузырьков, мультивезикулярных тел, отработанных органоидов и их
компонентов, а также нейротрофических и нейроростовых субстанций и наконец
вирусов.
Кинезины
обеспечивают транспорт в обоих направлениях (прямой и обратный), но во всех
случаях этот транспорт идет к «+ » –
концу микротрубочки. Денеины
участвуют в транспорте по микротрубочкам к ее «- » — концу. Миозины – это
транспортные белки, которые, в основном, участвуют в обратном транспорте
мембранных пузырьков, мультивезикулярных тел, отработанных органоидов и их
компонентов, а также нейротрофических и нейроростовых субстанций и вирусов.
Кроме того, миозины принимают участие и в прямом транспорте компонентов цитоскелета по отросткам и телу нейрона (например, с его помощью перемещаются
короткие мобильные микротрубочки). Важную роль миозины играют в росте отростков
и их ретракции в процессе развития нейронов и миграции клеток.
Механизмы аксонного и дендритного транспорта
Прямой аксональный транспорт
осуществляют моторные молекулы, связанные с системой цитоскелета и
плазматической мембраной. Моторная часть молекул кинезина или денеина
связывается с микротрубочкой, а хвостовая ее часть – с транспортируемым
материалом, с аксональной мембраной или с соседними элементами цитоскелета. В
обеспечении транспорта по отросткам принимают участие и ряд вспомогательных
белков (адапторов), ассоциированных с кинезином или денеином. Все процессы идут со
значительной затратой энергии.
Обратный
(ретроградный) транспорт.
В
аксонах основным механизмом обратного транспорта является система денеиновых и
миозиновых моторных белков. Морфологическим субстратом этого транспорта
являются: в аксоне – мультивезикулярные тела и сигнальные эндосомы, в дендритах
– мультивезикулярные и мультиламеллярные тела.
В
дендритах обратный транспорт осуществляется молекулярными комплексами не только денеина, но и кинезина. Это связано с тем, что (как указывалось ранее) в
проксимальных участках дендритов микротрубочки ориентированы во взаимопротивоположном направлении, а транспортировку молекул и органоидов к «+ » – концу микротрубочек осуществляют
только кинезиновые комплексы. Как и в случае прямого транспорта, разные
компоненты и вещества транспортируются ретроградно в разных нейронах с разной
скоростью, и, по – видимому, разными способами.
Большую
роль в транспортных процессах в нейроне играет гладкий эндоплазматический ретикулум. Показано, что по всей длине отростков нейрона распространяется непрерывная
разветвленная сеть цистерн гладкого ретикулума. Концевые ветвления этой сети
проникают в пресинаптические участки синапсов, где от них отшнуровываются синаптические
пузырьки. Именно по его цистернам
быстро транспортируются многие медиаторы
и нейромодуляторы, нейросекреты, ферменты их синтеза и распада, ионы кальция и
другие компоненты аксотока. Молекулярные механизмы этой разновидности
транспорта пока не ясны.
Дендритный
транспорт
Долгое
время экспериментально подтвердить наличие транспорта в дендритах не удавалось
из-за значительного объема синтеза белков собственно в дендритах. Только с
появлением методики внутриклеточной инъекции меченых предшественников синтеза
белка и других компонентов цитоплазмы, удалось показать, что в дендритах, также
как и в аксонах, имеется транспорт. Скорость прямого и обратного транспорта, в
дендритах сопоставима со скоростью прямого быстрого аксонального транспорта.
По
дендритам транспортируются вещества, которые либо не транспортируются по
аксонам, либо транспортируются в очень ограниченном количестве (например:
ферменты распада медиаторов, компоненты постсинаптических утолщений, ганглиозиды (специфические гликолипиды нейрональных мембран), нейрогормоны и
нейротрофические факторы).
Наличие
одновременно прямого и обратного транспорта в отростках нейронов создает
проблему их взаимодействия друг с другом. Направление транспортных потоков в
нейроне зависит, как полагают, от баланса между прямым и обратным транспортом и
этот баланс может быть самым различным.
Состояние
цитоскелета нейрона и моторных комплексов сильно сказывается на общей
морфологии его отростков. Показано, что в зависимости от того, какие компоненты цитоскелета или моторные молекулы активированы или не работают, форма, длина и
толщина отростков сильно изменяется.
Как
и в случае прямого транспорта, разные компоненты и вещества транспортируются
ретроградно в разных нейронах с разной скоростью, и, по – видимому, разными
способами.
Таблица.
4 Скорости ретроградного аксонного
транспорта различных молекул в периферической нервной системе (по: Reynolds
et
al.,
2000 с изменениями)
|
Транспортируемое |
Скорость |
Популяции |
|
NGF |
2-5 10-13 |
Симпатические Чувствительные |
|
Фермент |
Седалищный |
|
|
Вторичные |
28-57 (8-16 |
Седалищный |

Таким
образом, в нейронах существует хорошо развитый цитоскелет и связанная с ним эффективная система прямого и обратного транспорта по отросткам
разнообразных материалов и субстанций.
5.2.5. АКСОННЫЙ ТРАНСПОРТ
Наличие у нейрона
отростков, длина которых может достигать
1 м (например, аксоны, ин-нервирующие
мускулатуру конечностей), создает
серьезную проблему внутриклеточной
связи между различными участками нейрона
и ликвидации возможных повреждений его
отростков. Основная масса веществ
(структурных белков, ферментов,
полисахаридов, липидов и др.) образуется
в трофическом центре (теле) нейрона,
расположенном преимущественно около
ядра, а используются они в различных
участках нейрона, включая его отростки.
Хотя в аксонных окончаниях существуют
синтез медиаторов, АТФ и повторное
использование мембраны пузырьков после
выделения медиатора, все же необходима
постоянная доставка ферментов и
фрагментов мембран из тела клетки.
Для транспорта этих веществ (например,
белков) путем диффузии на расстояние,
равное максимальной длине аксона
(около 1 м), потребовалось бы 50 лет! Для
решения этой задачи эволюция
сформировала специальный вид транспорта
в пределах отростков нейрона, который
более хорошо изучен в аксонах и получил
название аксонного транспорта. С помощью
этого процесса осуществляется трофическое
влияние не только в пределах различных
участков нейрона, но и на иннервируе-
мые клетки. В
последнее время появились данные о
существовании нейроплазматичес-кого
транспорта в дендритах, который
осуществляется из тела клетки со
скоростью около 3 мм в сутки. Различают
быстрый и медленный аксонный транспорт.
А.
Быстрый аксонный транспорт
идет
в двух направлениях: от тела клетки до
аксонных окончаний (антеградный
транспорт, скорость 250-400 мм/сут) и в
противоположном направлении
(ретроградный транспорт, скорость
200-300 мм/сут). Посредством анте-градного
транспорта в аксонные окончания
доставляются везикулы, образующиеся в
аппарате Гольджи и содержащие
гликопротеины мембран, ферменты,
медиаторы, липиды и другие вещества.
Посредством ретроградного транспорта
в тело нейрона переносятся везикулы,
содержащие остатки разрушенных структур,
фрагменты мембран, ацетилхоли-нэстераза,
неидентифицированные «сигнальные
вещества», регулирующие синтез белка
в соме клетки. В патологических условиях
по аксону к телу клетки могут
транспортироваться вирусы полиомиелита,
герпеса, бешенства и столбнячный
экзотоксин. Многие вещества, доставленные
путем ретроградного транспорта,
подвергаются разрушению в лизосомах.
Быстрый
аксонный транспорт осуществляется
с помощью специальных структурных
элементов нейрона: микротрубочек и
микрофиламентов, часть которых
представляет собой актиновые нити
(актин составляет 10-15 % белков нейрона).
Для транспорта необходима энергия АТФ.
Разрушение микротрубочек (например,
колхицином) и микрофиламентов
(цитохолазином В), снижение уровня АТФ
в аксоне более чем в 2 раза и падение
концентрации Са 2+
блокируют аксонный транспорт.
Б.
Медленный аксонный транспорт
осуществляется
только в антеградном направлении и
представляет собой передвижение всего
столба аксоплазмы. Он выявляется в
опытах со сдавлением (перевязкой) аксона.
При этом происходит увеличение диаметра
аксона проксимальнее перетяжки в
результате «наплыва гиалоплазмы» и
утончение аксона за местом сдавления.
Скорость медленного транспорта равна
1-2 мм/сут, что соответствует скорости
роста аксона в онтогенезе и при его
регенерации после его повреждения. С
помощью этого транспорта перемещаются
образованные в эндоплазматической сети
белки микротрубочек и микрофиламентов
(тубулин, актин и др.), ферменты цитозоля,
РНК, белки каналов, насосов и другие
вещества. Медленный аксонный транспорт
не на-
рушается при
разрушении микротрубочек, но прекращается
при отделении аксона от тела нейрона,
что свидетельствует о разных механизмах
быстрого и медленного аксонного
транспорта.
В.
Функциональная роль аксонного транспорта.
1.
Антеградный и ретроградный транспорт
белков и других веществ необходимы для
поддержания структуры и функции аксона
и его пресинаптических окончаний, а
также для таких процессов, как аксонный
рост и образование синаптических
контактов.
2. Аксонный транспорт
участвует в трофическом влиянии
нейрона на иннервируемую клетку, так
как часть транспортируемых веществ
выделяется в синаптическую щель и
действует на рецепторы постсинаптической
мембраны и близлежащих участков мембраны
иннервируемой клетки. Эти вещества
участвуют в регуляции обмена веществ,
процессов размножения и дифференцировки
ин-нервируемых клеток, формируя их
функциональную специфику. Например,
в опытах с перекрестной иннервацией
быстрых и медленных мышц показано,
что свойства мышц меняются в зависимости
от типа иннервиру-ющего нейрона, его
нейротрофического воздействия.
Передатчики трофических влияний нейрона
до сих пор точно не определены, важное
значение в этом плане придается
полипептидам и нуклеиновым кислотам.
3. Роль аксонного
транспорта особенно ярко выявляется
при повреждении нерва. Если нервное
волокно на каком-либо участке прервано,
его периферический отрезок, лишенный
контакта с телом нейрона, подвергается
разрушению, которое называется
валле-ровской дегенерацией. В течение
2-3 сут наступает распад нейрофибрилл,
митохондрий, миелина и синаптических
окончаний. Надо отметить, что распаду
подвергается участок волокна, снабжение
которого кислородом и питательными
веществами с кровотоком не прекращается.
Считают, что решающим механизмом
дегенерации является прекращение
аксонного транспорта веществ от тела
клетки до синаптических окончаний.
4. Аксонный
транспорт играет важную роль и при
регенерации нервных волокон.
Аксональный
транспорт (аксоток)
— это перемещение веществ от тела
нейрона в отростки (антероградный
аксоток)
и в обратном направлении (ретроградный
аксоток).
Различают медленный
аксональный
ток веществ (1-5 мм в сутки) и быстрый
(до
1-5 м в сутки). Обе транспортные системы
присутствуют как в аксонах, так и в
дендритах. Аксональный транспорт
обеспечивает единство нейрона. Он
создаёт постоянную связь между телом
нейрона (трофическим центром) и
отростками. Основные синтетические
процессы идут в перикарионе. Здесь
сосредоточены необходимые для этого
органеллы. В отростках синтетические
процессы протекают слабо.
Антероградная
быстрая система транспортирует
к нервным окончаниям белки и органеллы,
необходимые для синаптических функций
(митохондрии, фрагменты мембран,
пузырьки, белки-ферменты, участвующие
в обмене нейромедиаторов, а также
предшественники нейромедиаторов).
Ретроградная
система возвращает
в перикарион использованные и поврежденные
мембраны и белки для деградации в
лизосомах и обновления, приносит
информацию о состоянии периферии,
факторы роста нервов. Медленный
транспорт —
это антероградная система, проводящая
белки и другие вещества для обновления
аксоппазмы зрелых нейронов и обеспечения
роста отростков при их развитии и
регенерации.
Ретроградный
транспорт может
иметь значение в патологии. За счёт него
нейротропные вирусы (герпеса, бешенства,
полиомиелита) могут перемещаться с
периферии в центральную нервную
систему.
Нейроглия
Глиоциты
выполняют в нервной ткани вспомогательные
функции: опорную, разграничительную,
трофическую, секреторную
и
защитную. Они поддерживают постоянно
среды вокруг нейронов. Клетки нейроглии
делятся на 2 группы: макроглию и микроглию.
Клетки макроглии бывают трех типов.
Эпендимоциты.
Выстилают каналы и желудочки единого
и головного мозга, по которым циркулирует
спинномозговая жидкость (ликвор). Эти
клетки напоминают однослойный
призматический
эпителий.
На апикальных концах эпендимоцитов
расположены реснички, помогающие
движению спинномозговой жидкости. Через
апикальные концы эпендимоциты могут
выделять биологически активные вещества,
которые с лидером разносятся по всему
мозгу. От базальных концов эпендимоцитов
отходят отростки, которые могут идти
через весь мозг. В желудочках мозга
находятся сосудистые
сплетения. Они
покрыты специализированными эпендимоцитами,
участвующими в образований ликвора.
Астроциты
.
Различают протоплазматические
и
волокнистые
астроциты.
Протоплазматические
астроциты имеют
короткие толстые отростки. Они расположены
в сером веществе мозга, выполняют
разграничительную и трофическую
функции. Волокнистые
астроциты находятся
в белом веществе, имеют многочисленные
тонкие длинные отростки, которые
оплетают кровеносные сосуды мозга,
образуя периваскулярные глиальные
пограничные мембраны. Их отростки также
изолируют синапсы. Таким образом, они
изолируют нейроны и кровеносные сосуды
и участвуют в образовании
гемато-энцефалического барьера,
обеспечивают обмен веществ между кровью
и нейронами. Они также участвуют в
образовании оболочек мозга и выполняют
опорную функцию (образуют каркас мозга).
Олигодендроциты
имеют мало отростков, окружают нейроны,
выполняя трофическую (участие в питании
нейронов) и разграничительную Функции.
Олигодендроциты, расположенные вокруг
тел нейронов, называются мантийными
глиоцитами. Олигодендроциты,
расположенные в периферической нервной
системе и образующие оболочки вокруг
отростков нейронов, называют
леммоцитами
(шванновскими клетками).
Микроглия
(глиальные макрофаги). Образуйся из
костномозговых предшественников
моноцитов. Покоящиеся микроглиоциты
имеют короткие ветвящиеся отростки.
Под действием микроорганизмов и продуктов
распада нервной ткани они активируются,
теряют отростки, округляются и превращаются
в «зернистые шары» (реактивная микроглия).
При этом они, как макрофаги, уничтожают
разрушенные нервные и глиальные клетки.
Источники
развития
— нервная
трубка, нервный гребень (ганглиозные
пластинки) и
плакоды.
Нервная трубка образуется
в результате смыкания краёв нервного
желобка, развивающегося из эктодермы.
Нервные
гребни расположены
между нервной трубкой и эктодермой Они
образуются в результате выселения
клеток из утолщенных краев нервного
желобка — нервных
валиков. Плакоды представляют
собой эктодермы по бокам нервной трубки
на головном конце зародыша. Нейробласты
нервной
трубки дают начало нервным клеткам, а
глиобласты — глиальным клеткам головного
и спинного мозга. Из клеток нервного
гребня происходят нейроны и нейроглия
всех нервных ганглиев, а из плакод —
рецепторные (нейросенсорные) клетки
органа обоняния, нейроны слухового
и вестибулярного ганглиев. Клетки
микроглии образуются из промоноцитов
красного костного мозга.
В
ходе эмбриогенеза до 85% образующихся
нейронов гибнет в результате апоптоза
(генетически запрограммированной
смерти). Погибают дефектные нейроны (с
повреждённой ДНК), нейроны, которые
не нашли свои «клетки-мишени» или
оказались избыточными, «лишними».