ОСНОВЫ
ЖИЗНЕДЕЯТЕЛЬНОСТИ НЕЙРОНОВ
СТРУКТУРО-ФУНКЦИОНАЛЬНАЯ
ХАРАКТЕРИСТИКА НЕРВНЫХ КЛЕТОК
А. Нейрон – это
структурно – функциональная единица
нервной ткани. Выделяют тело нейрона
и его отростки. Оболочка нейрона
(клеточная мембрана) образует замкнутое
пространство, содержащее протоплазму
(цитоплазма и ядро). Цитоплазма состоит
из основного вещества (цитозоль,
гиалоплазма) и органелл. Гиалоплазма
под электронными микроскопом выглядит
относительно гомогенным веществом и
является внутренней средой нейрона.
Большинство органелл и ядро нейрона,
как и любой другой клетки, заключены в
свои отсеки (компартменты), образуемые
собственными (внутриклеточными)
мембранами, обладающими избирательной
проницаемостью к отдельным ионам и
частицам, находящимся в гиалоплазме и
органеллах. Это определяет отличительный
состав их друг от друга.
Мозг человека
содержит около 25 млрд. нервных клеток,
взаимодействие между которыми
осуществляется посредством множества
синапсов (межклеточные соединения),
число которых в тысячи раз больше самих
клеток (1015 – 1016), так как их аксоны
многократно делятся дихотомически.
Нейроны оказывают свое влияние на
органы и ткани также посредством
синапсов. Нервные клетки имеются и вне
ЦНС : периферический отдел вегетативной
нервной системы, афферентные нейроны
спинномозговых ганглиев и ганглиев
черепных нервов. Периферических нервных
клеток намного меньше, чем центральных,
— всего около 25 млн. Важную роль в
деятельности нервной системы играют
глиальные клетки (см. раздел 2.1, Д).
Отростки нейрона
представляют собой большое число
дендритов и один аксон (рис. 2.1). Нервные
клетки имеют электрический заряд, как
и другие клетки животного организма и
даже растений (рис. 2.2). Потенциал покоя
(ПП) нейрона составляет 60-80 мВ, ПД –
нервный импульс – 80-110 мВ. Сома и дендриты
покрыты нервными окончаниями –
синаптическими бутонами и отростками
глиальных клеток. На одном нейроне
число синаптических
бутонов может достигать 10 000. Аксон
начинается от тела клетки
аксонным холмиком. Диаметр тела клетки
составляет 10-100 мкм, аксона — 1-6 мкм, на
периферии длина аксона может достигать
1 м и более.. Нейроны мозгаобразуют
колонки, ядра и слой, выполняющие
определенные функции. Клеточные
скопления
составляют серое вещество мозга. Между
клетками проходят немиелинизированные
и миелинизированные нервные волокна
(соответственно
дендриты и аксоны нейронов).
Б.
Классификация нейронов.
Нейроны делят на следующие группы.
-
По
медиатору, выделяющемуся
в окончаниях аксонов, раз
личают
нейроны адренергические, холинергические,
серотонинергические
и т.д., -
В
зависимости от отдела ЦНС выделяют
нейроны соматической
и вегетативной нервной системы. -
По
направлению информации различают
следующие нейроны:
-
афферентные,
воспринимающие с помощью рецепторов
ин
формацию о внешней и внутренней
среде организма и пере
дающие
ее в вышележащие отделы ЦНС; -
эфферентные,
передающие информацию к рабочим органам
—
эффекторам
(нервные клетки, иннервирующие
эффекторы,
иногда
называют эффекторными);
«вставочные
(интернейроны), обеспечивающие
взаимодействие
между нейронами ЦНС.
4. По
влиянию выделяют
возбуждающие и тормозящие нейроны.
-
По
активности различают
фоново-активные и «молчащие» нейроны,
возбуждающиеся только в ответ на
раздражение. Фоново-
активные
нейроны отличаются общим рисунком
генерации им
пульсов,
так как одни нейроны разряжаются
непрерывно (ритмично
или аритмично), другие — пачками
импульсов. Интервал между импульсами
в пачке составляет миллисекунды, между
пачками —
секунды. Фоново-активные
нейроны играют важную роль в под
держании
тонуса ЦНС и особенно коры большого
мозга. -
По
воспринимаемой сенсорной информации
нейроны
делят на моно-, би- и полимодальные.
Мономодальными являются нейроны
центра
слуха в коре большого мозга. Бимодальные
нейроны встречаются
во вторичных зонах анализаторов в коре
(нейроны вторичной
зоны зрительного анализатора в коре
большого мозга реагируют
на световые и звуковые раздражители).
Полимодальные нейроны
— это нейроны ассоциативных зон мозга,
моторной коры;
они
реагируют на раздражения рецепторов
кожного, зрительного,слухового
и других анализаторов.

Рис. 2.1. Мотонейрон
спинного мозга. Указаны функции отдельных
структурных элементов нейрона[Эккерт
Р., Рэнделл Д., Огастин Дж., 1991]
В.
Функциональные структуры нейрона. 1.
Структуры,
обеспечивающие
синтез макромолекул, которые
транспортируются по аксону
и дендритам, — это сома (тело нейрона),
выполняющая трофическую функцию по
отношению к
отросткам
(аксону и дендритам) и клеткам-эффекторам.
Отросток, лишенный
связи
с телом нейрона, дегенерирует.
2.
Структуры, воспринимающие импульсы от
других нервных клеток,
— это тело и
дендриты нейрона
с расположенными на
них
шипиками,
занимающие
до 40% от поверхности сомы нейрона и
дендритов.ЛЕсли
шипики
не получают импульсацию, то они исчезают.
Импульсы могут
поступать
и к окончанию аксона — аксо-аксонные
синапсы.
Это происходит, например, в случае
пресинап-тического
торможения.
3.
Структуры, в которых обычно возникает
ПД (генераторный пункт
ПД),
—
аксонный
холмик.
-
Структуры,
проводящие возбуждение к другому
нейрону или
к
эффектору,
— аксон. -
Структуры,
передающие импульсы на другие клетки,
— си
напсы.
Г.
Классификация
синапсов ЦНС. Основу
классификации составляет
несколько признаков. 1.
По
способу передачи сигналов различают
химические синапсы (наиболее
распространенные в ЦНС),
в которых посредником (медиатором)
передачи является химическое
вещество; электрические, в которых
сигналы передаются электрическим
током, и
смешанные
синапсы — электрохимические.
2. В
зависимости от местоположения выделяют
аксосоматиче-
ские,
аксодендритные, аксо-аксонные,
дендросоматические, денд-
родендритные
синапсы.
3. По
эффекту различают
возбуждающие и тормозящие синапсы.
В
процессе деятельности нервной системы
отдельные нейроны
объединяются
в ансамбли (модули), нейронные сети.
Последние могут
включать несколько нейронов, десятки,
тысячи нейронов,
при
этом совокупность нейронов, образующих
модуль, обеспечивает появление у
модуля новых свойств, которыми не
обладают отдельные
нейроны. Деятельность каждого нейрона
в составе модуля
становится функцией не только поступающих
к
нему
сигналов,
но и
функцией
процессов, обусловленных той или иной
конструкцией модуля (П.Г.Костюк).
Д.
Глиальные клетки (нейроглия — «нервный
клей»). Эти
клетки более
многочисленны, чем нейроны, составляют
около 50% от объема
ЦНС.
Они способны к делению в течение всей
жизни. По размеру
глиальные клетки в 3-4 раза меньше
нервных, их число огромно
— достигает 14 х 10″, с возрастом
увеличивается (число нейронов
уменьшается). Тела нейронов, как и их
аксоны, окружены глиальными
клетками. Глиальные клетки выполняют
несколько
функций:
опорную,
защитную, изолирующую, обменную
(снабжение
нейронов питательными веществами).
Микроглиальные клетки
способны к фагоцитозу, ритмическому
изменению своего объема
(период «сокращения» — 1,5 мин, «расслабления»
— 4 мин). 11,иклы
изменения объема повторяются через
каждые 2-20
ч.
Полагают,
что пульсация способствует продвижению
аксоплазмы в нейронах и
влияет
на ток межклеточной жидкости. Мембранный
потенциал
клеток нейроглии
составляет
70-90
мВ,
однако
ПД они не
генерируют, генерируют только локальные
токи, электротони-чески распространяющиеся
от одной клетки к другой. Процессы
возбуждения в нейронах и
электрические
явления в глиальных клетках,
по-видимому,
взаимодействуют.
Е.
Цереброспинальная жидкость (ликвор) —
бесцветная прозрачная
жидкость, заполняющая мозговые желудочки,
спинномозговой канал
и субарахноидальное пространство. Ее
происхождение связано
с интерстициальной жидкостью мозга.
Значительная часть цереброспинальной
жидкости образуется в специализированных
сплетениях желудочков мозга.
Непосредственной питательной
средой
клеток мозга является
интерстициальная жидкость, в которую
клетки выделяют также продукты своего
обмена, Цереброспинальная
жидкость
представляет
собой совокупность фильтрата
плазмы крови и интерстициальной
жидкости; она содержит
около 90%
воды
и
примерно 10%
сухого
остатка (2% -органические,
8% — неорганические вещества). От плазмы
крови она
отличается, как и
межклеточная
жидкость других тканей, низким
содержанием белка (0,1 г/л, в плазме — 75
г/л), меньшим содержанием
аминокислот (0,8 и
2 ммоль/л
соответственно)
и глюкозы (3,9
и около 5 ммоль/л соответственно). Ее
объем 100-200 мл (12-14%
от общего объема мозга), за сутки
вырабатывается около 600
мл. Обновление этой
жидкости
происходит 4-8 раз в сутки, давление
цереброспинальной жидкости составляет
7-14 мм рт. ст.,
в вертикальном положении тела — в 2 раза
больше. Цереброспинальная жидкость
выполняет также защитную
роль:
является
своеобразной гидравлической «подушкой»
мозга, обладает бактерицидными
свойствами: ликвор
содержит иммуноглобулины
классов G
и А, систему комплемента, моноциты и
лимфоциты. Отток цереброспинальной
жидкости происходит несколькими
путями: 30—40% ее оттекает через
субарахноидальное пространство
в продольный синус венозной системы
головного мозга;
10-20% — через периневральные
пространства
черепных и
спинномозговых
нервов в
лимфатическую
систему; часть жидкости
реабсорбируется сосудистыми сплетениями
мозга.
2.1. структурно-функциональная характеристика нервных клеток
А. Нейрон это структурно-функциональная единица нервной ткани. Выделяют тело нейрона и его отростки. Оболочка нейрона (клеточная мембрана) образует замкнутое пространство, содержащее протоплазму (цитоплазма и ядро). Цитоплазма состоит из основного вещества (цитозоль, гиалоплазма) и органелл. Гиалоплазма под электронным микроскопом выглядит относительно гомогенным веществом и является внутренней средой нейрона. Большинство органелл и ядро нейрона, как и любой другой клетки, заключены в свои отсеки (компартменты), образуемые собственными (внутриклеточными) мембранами, обладающими избирательной проницаемостью к отдельным ионам и частицам, находящимся в гиалоплазме и органеллах. Это определяет отличительный состав их друг от друга.
Мозг человека содержит около 25 млрд. нервных клеток, взаимодействие между которыми осуществляется посредством множества синапсов (межклеточные соединения), число которых в тысячи раз больше самих клеток (1015-1016), так как их аксоны многократно делятся дихотомически. Нейроны оказывают свое влияние на органы и ткани также, посредством синапсов. Нервные клетки имеются и вне ЦНС: периферический отдел вегетативной нервной системы, афферентные нейроны спинномозговых ганглиев и ганглиев черепных нервов. Периферических нервных клеток намного меньше, чем центральных, всего около 25 млн. Важную роль в деятельности нервной системы играют глиальные клетки (см. раздел 2.1, Д).
Отростки нейрона представляют собой большое число дендритов и один аксон (рис. 2.1). Нервные клетки имеют электрический заряд, как и другие клетки животного организма и даже растений (рис. 2.2). Потенциал покоя (ПП) нейрона составляет 60-80 мВ, ПД нервный импульс 80-110 мВ. Сома и дендриты покрыты нервными окончаниями синаптическими бутонами и отростками глиальных клеток. На одном нейроне число синаптических бутонов может достигать 10 000. Аксон начинается от тела клетки аксонным холмиком. Диаметр тела клетки составляет 10-100 мкм, аксона 1-6 мкм, на периферии длина аксона может достигать 1 м и более. Нейроны мозга образуют колонки, ядра и слои, выполняющие определенные функции. Клеточные скопления составляют серое вещество мозга. Между клетками проходят немиелинизированные и миелинизированные нервные волокна (соответственно дендриты и аксоны нейронов).
Б. Классификация нейронов. Нейроны делят на следующие группы.
1. По медиатору, выделяющемуся в окончаниях аксонов, различают нейроны адренергические, холинергические, серотонинергические и т.д.
2. В зависимости от отдела ЦНС выделяют нейроны соматической и вегетативной нервной системы.
3. По направлению информации различают следующие нейроны:
• афферентные, воспринимающие с помощью рецепторов информацию о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС;
• эфферентные, передающие информацию к рабочим органам -эффекторам (нервные клетки, иннервирующие эффекторы, иногда называют эффекторными);
• вставочные (интернейроны), обеспечивающие взаимодействие между нейронами ЦНС.
4. По влиянию выделяют возбуждающие и тормозящие нейроны.
5. По активности различают фоново-активные и «молчащие» нейроны, возбуждающиеся только в ответ на раздражение. Фоново-активные нейроны отличаются общим рисунком генерации импульсов, так как одни нейроны разряжаются непрерывно (ритмично или аритмично), другие пачками импульсов. Интервал между импульсами в пачке составляет миллисекунды, между пачками секунды. Фоново-активные нейроны играют важную роль в поддержании тонуса ЦНС и особенно коры большого мозга.
6. По воспринимаемой сенсорной информации нейроны делят на моно-, бии полимодальные. Мономодальными являются нейроны центра слуха в коре большого мозга. Бимодальные нейроны встречаются во вторичных зонах анализаторов в коре (нейроны вторичной зоны зрительного анализатора в коре большого мозга реагируют на световые и звуковые раздражители). Полимодальные нейроны это нейроны ассоциативных зон мозга, моторной коры; они реагируют на раздражения рецепторов кожного, зрительного, слухового и других анализаторов.
В. Функциональные структуры нейрона. 1. Структуры, обеспечивающие синтез макромолекул, которые транспортируются по аксону и дендритам, это сома (тело нейрона), выполняющая трофическую функцию по отношению к отросткам (аксону и дендритам) и клеткам-эффекторам. Отросток, лишенный связи с телом нейрона, дегенерирует.
2. Структуры, воспринимающие импульсы от других нервных клеток, это тело и дендриты нейрона с расположенными на них шипиками, занимающие до 40% от поверхности сомы нейрона и дендритов. Если шипики не получают импульсацию, то они исчезают. Импульсы могут поступать и к окончанию аксона аксо-аксонные синапсы. Это происходит, например, в случае пресинаптического торможения.
3. Структуры, в которых обычно возникает ПД (генераторный пункт ПД), аксонный холмик.
4. Структуры, проводящие возбуждение к другому нейрону или к эффектору, аксон.
5. Структуры, передающие импульсы на другие клетки, синапсы.
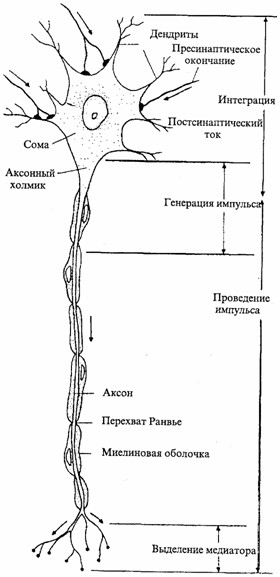
Рис. 2.1. Мотонейрон спинного мозга. Указаны функции отдельных структурных элементов нейрона [Эккерт Р., Рэнделл Д., Огастин Дж.. 1991]
Г. Классификация синапсов ЦНС. Основу классификации составляет несколько признаков. 1. По способу передачи сигналов различают химические синапсы (наиболее распространенные в ЦНС), в которых посредником (медиатором) передачи является химическое вещество; электрические, в которых сигналы передаются электрическим током, и смешанные синапсы электрохимические.
2. В зависимости от местоположения выделяют аксосоматические, аксодендритные, аксо-аксонные, дендросоматические, дендродендритные синапсы.
3. По эффекту различают возбуждающие и тормозящие синапсы. В процессе деятельности нервной системы отдельные нейроны объединяются в ансамбли (модули), нейронные сети. Последние могут включать несколько нейронов, десятки, тысячи нейронов, при этом совокупность нейронов, образующих модуль, обеспечивает появление у модуля новых свойств, которыми не обладают отдельные нейроны. Деятельность каждого нейрона в составе модуля становится функцией не только поступающих к нему сигналов, но и функцией процессов, обусловленных той или иной конструкцией модуля (П.Г.Костюк).
Д. Глиальные клетки (нейроглия «нервный клей»). Эти клетки более многочисленны, чем нейроны, составляют около 50% от объема ЦНС. Они способны к делению в течение всей жизни. По размеру глиальные клетки в 3-4 раза меньше нервных, их число огромно достигает 14 х 1010, с возрастом увеличивается (число нейронов уменьшается). Тела нейронов, как и их аксоны, окружены глиальными клетками. Глиальные клетки выполняют несколько функций: опорную, защитную, изолирующую, обменную (снабжение нейронов питательными веществами). Микроглиальные клетки способны к фагоцитозу, ритмическому изменению своего объема (период «сокращения» 1,5 мин, «расслабления» 4 мин). Циклы изменения объема повторяются через каждые 2-20 ч. Полагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Мембранный потенциал клеток нейроглии составляет 70-90 мВ, однако ПД они не генерируют, генерируют только локальные токи, электротонически распространяющиеся от одной клетки к другой. Процессы возбуждения в нейронах и электрические явления в глиальных клетках, по-видимому, взаимодействуют.
Е. Цереброспинальная жидкость (ликвор) бесцветная прозрачная жидкость, заполняющая мозговые желудочки, спинномозговой канал и субарахноидальное пространство. Ее происхождение связано с интерстициальной жидкостью мозга. Значительная часть цереброспинальной жидкости образуется в специализированных сплетениях желудочков мозга. Непосредственной питательной средой клеток мозга является интерстициальная жидкость, в которую клетки выделяют также продукты своего обмена. Цереброспинальная жидкость представляет собой совокупность фильтрата плазмы крови и интерстициальной жидкости; она содержит около 90% воды и примерно 10% сухого остатка (2% -органические, 8% неорганические вещества). От плазмы крови она отличается, как и межклеточная жидкость других тканей, низким содержанием белка (0,1 г/л, в плазме 75 г/л), меньшим содержанием аминокислот (0,8 и 2 ммоль/л соответственно) и глюкозы (3,9 и около 5 ммоль/л соответственно). Ее объем 100-200 мл (12-14% от общего объема мозга), за сутки вырабатывается около 600 мл. Обновление этой жидкости происходит 4-8 раз в сутки, давление цереброспинальной жидкости составляет 7-14 мм рт. ст., в вертикальном положении тела в 2 раза больше. Цереброспинальная жидкость выполняет также защитную роль: является своеобразной гидравлической «подушкой» мозга, обладает бактерицидными свойствами: ликвор содержит иммуноглобулины классов G и А, систему комплемента, моноциты и лимфоциты. Отток цереброспинальной жидкости происходит несколькими путями: 30-40% ее оттекает через субарахноидальное пространство в продольный синус венозной системы головного мозга; 10-20% через периневральные пространства черепных и спинномозговых нервов в лимфатическую систему; часть жидкости реабсорбируется сосудистыми сплетениями мозга.
Медиатором, образующимся в окончаниях парасимпатических нервов, а также симпатических вазодилататоров и симпатических нервов потовых желез, является ацетилхолин; медиатором, образующимся в окончаниях постганглионарных симпатических нервов (за исключением нервов потовых желез и симпатических вазодилататоров),— норадреналин (адреналин, лишенный одной метильной группы).
Медиаторы, образующиеся в окончаниях вегетативных нервных волокон, действуют на иннервируемые ими клетки дольше по сравнению со временем действия медиатора (ацетилхолина) в окончаниях соматических нервов. По-видимому, это объясняется меньшей активностью ферментов, разрушающих медиатор.
Медиаторы образуются также терминалями преганглионарных волокон в синапсах ганглиев вегетативной нервной системы. Первые доказательства этого факта были получены А. В. Кибяковым в 1933 г. в опытах, в которых он пропускал через сосуды верхнего шейного симпатического узла кошки раствор Рингера — Локка и обнаружил при раздражении преганглионарных симпатических волокон в растворе, оттекающем от узла, адреналиноподобное вещество.
В дальнейшем было показано, что возбуждающим медиатором в синапсах преганглионарных волокон является ацетилхолин. Адреналин оказался медиатором, вызывающим торможение активности нейронов симпатического ганглия. Возможно, что тормозящие волокна, в которых образуется адреналиноподобное вещество, представляют собой постганглионарные волокна, иннервирующие узел и изменяющие его функциональное состояние.
Особенностью действия ацетилхолина в синапсах ганглиев является то, что оно не прекращается после отравления узла атропином, но исчезают после отравления никотином. На этом основании считают, что существует два вида структур, чувствительных к ацетилхолину; одни из них — М-холинорецепторы — теряют чувствительность к ацетилхолину под влиянием атропина, другие – Н-холинорецепторы — под влиянием никотина и других веществ, называемых ганглиоблокаторами (гексоний и др.).
В области концевых разветвлений симпатических нервных волокон имеются расширения — варикозы, в которых находятся пузырьки — везикулы, подобные имеющимся в синапсах. Толщина этих расширений концевых нервных волокон -до 2 мкм, длина — 0,5—3 мкм. Таких варикозов может быть 15—30 на протяжении 100 мкм. В варикозах содержится в 20—100 раз больше норадреналина, чем в остальных участках постганглионарного волокна. В расширенной части концевых разветвлений, а не только в синапсах симпатических нервных волокон возможно высвобождение медиатора, действующего на иннервированную ими ткань.
В зависимости от того какой медиатор выделяется окончаниями аксонов вегетативных нейронов,п редложено разделять нейроны нахолинергические и адренергические. Холинергическими являются эфферентные нейроны интрамуральных парасимпатических ганглиев и эфферентные нейроны парасимпатических центров среднего, продолговатого и спинного мозга, а также эфферентные нейроны симпатических центров спинного мозга и те эфферентные нейроны периферических симпатических ганглиев, которые иннервируют потовые железы и обеспечивают расширение сосудов работающих мышц. Окончания аксонов этих нейронов выделяют ацетилхолин. Адренергическими являются все остальные эфферентные нейроны симпатических ганглиев. В окончаниях аксонов и в контактах, образованных этими аксонами с гладкомышечными клетками и другими структурами, выделяется норадреналин. Освобождающийся в терминалах аксонов медиатор — ацетилхолин или норадреналин взаимодействует со специфическим белком постсинаптической мембраны, образующим комплексное соединение с медиатором.
Белок, с которым взаимодействует ацетилхолин, получил название холинорецептора, а белок, взаимодействующий с адреналином или норадреналином, назван адренорецептором. Соединение медиатора с соответствующим рецепторным веществом является начальной реакцией в цепи химических превращений, возникающих в клетке под влиянием приходящих к ней нервных импульсов.
Имеется два основных вида адренорецепторов, с которыми взаимодействует как адреналин, так и норадреналин: а- и β-адренорецепторы. Их существование установлено путем применения некоторых фармакологических препаратов, действующих избирательно на тот или другой вид адренорецепторов. В ряде органов находится оба вида адренорецепторов, которые могут вызывать либо разные, либо одинаковые реакции, или же имеется только один из адренорецепторов. В кровеносных сосудах имеются и α-и β-адренорецепторы. Показано, что соединение симпатического медиатора с α-адренорецепторами в артериальной стенке вызывает сужение артериол, а соединение с α-адренорецепторами приводит к расширению артериол. В кишечнике также имеются и а- и β-адренорецепторы; воздействие и на те и на другие тормозит сокращение гладкой мускулатуры. В сердце и бронхах нет α-адренорецепторов и здесь норадреналин и адреналин взаимодействуют только с β-адренорецепторами. В результате этого происходит усиление сердечных сокращений и расширение бронхов.
В механизме действия норадреналина и адреналина придают значение тому недавно открытому факту, что норадреналин и адреналин активируют энзим, находящийся в мембране мышечных клеток,— аденилциклазу. Этот энзим в присутствии ионов магния катализирует образование в клетке из АТФ циклического 3,5-аденозинмонофосфата. Это соединение — цАМФ вызывает ряд физиологических эффектов, в частности активирует некоторые энзимы энергетического обмена и стимулирует сердечную деятельность.
Кроме ацетилхолина и норадреналина, в вегетативной нервной системе найдены и другие медиаторы. В окончаниях симпатических нервных волокон обнаружен дофамин, выделение которого в синаптическую щель происходит под влиянием приходящих нервных импульсов. Полагают, что дофамин вступает во взаимодействие с α-адренорецепторами, расположенными на самих пресинаптических окончаниях, и тем самым тормозит выделение норадреналина.
Полагают, что на гладкую мускулатуру кишечника, матки, а возможно, и кровеносных сосудов может действовать серотонин, эффект которого напоминает действие медиатора ацетилхолина, но сохраняется после блокады М-холинорецепторов.
В желудке и кишечнике обнаружены интрамуральные эфферентные нейроны, возбуждение которых тормозит активность гладкой мускулатуры. Это торможение осуществляется путем выделения окончаниями аксонов этих нейронов пуринового нуклеотида аденозинтрифосфорной кислоты (АТФ). Медиаторный эффект принадлежит, по-видимому, самой АТФ. Указанные эфферентные нейроны получили название пуринергических.
Предполагают, что медиатором может быть и гистамин, так как в некоторых тканях обнаружены специфические H ₁- и Н ₂-гистаминорецепторы. Гистамин Является биологически активным веществом широкого спектра действия. Выявлено, что широко распространенный в синапсах ЦНС тормозной медиатор гамма-аминомасляная кислота — ГАМК тормозит проведение возбуждения в звездчатом ганглии, но облегчает передачу возбуждения в верхнем шейном, нижнем брыжеечном и в ганглиях солнечного сплетения.
После перерезки и перерождения вегетативных нервов чувствительность денервированных органов к соответствующим медиаторам возрастает. Если десимпатизировать любой орган, иннервированный симпатическими нервными волокнами (сердце, желудок, кишечник, сосуды, радужную оболочку глаза и др.), то он приобретает повышенную чувствительность к адреналину и норадреналину. Точно так же, если произвести парасимпатическую денервацию органа, он может приобрести повышенную чувствительность к ацетилхолину. Имеется ряд механизмов этой повышенной чувствительности денервированных тканей. Среди них следует указать на возрастание числа рецепторов на постсинаптической мембране, снижение активности или содержания в тканях фермента, расщепляющего адреналин (моноаминооксидаза) или расщепляющего ацетилхолин (ацетилхолинэстераза) и др.