The Immunological Synapse Part A
Yu Li, Jordan S. Orange, in Methods in Cell Biology, 2023
6 Data analysis
To manually quantify lipid ordering of presynaptic membranes using the Fiji version of ImageJ:
- 1.
-
Download and open software (https://imagej.net/software/fiji/).
- 2.
-
Import and open acquired images with Fiji (at least including three channels: two ratiometric channels for Di-4-ANEPPDHQ probe and one for CellMask deep red).
- 3.
-
Generate binary masks to identify cell membrane region from images of CellMask deep red channel via the Fiji “Threshold” function (via “Image › Adjust › Threshold” or “Ctrl + Shift + T”).
- 4.
-
Calculate GP values from images of the two ratiometric channels via the Fiji “image calculator” function (via “Image › Image Calculator…”). The equation used to calculate the GP value is: GP value = (Di4_A − Di4_B)/(Di4_A + Di4_B)
- 5.
-
Multiply the resulting images with corresponding binary masks to remove background and only preserve the GP values of the membranes of interest via the Fiji “image calculator” function.
- 6.
-
To visualize GP values, choose and apply a lookup table in Fiji and set the display scale manually to “0 to 1” in the “Brightness and Contrast” window. Select a lookup table and range that is consistent with the dynamic range of the GP values to emphasize
- 7.
-
To quantitatively analyze of GP values, obtain measurements via the “Analyze ›Measure” function of Fiji.
- 8.
-
For time-lapse imaging series apply this procedure individually to each frame.
To perform batched imaging analysis automatically using the Fiji version of ImageJ:
- 1.
-
Download and open software (https://imagej.net/software/fiji/).
- 2.
-
Import and open acquired images with Fiji (at least including three channels: two ratiometric channels for Di-4-ANEPPDHQ probe and one for CellMask deep red).
- 3.
-
Rename the images from 500 to 580 nm and 620–750 nm channels as “Di4_A_1, Di4_A_2, Di4_A_3, etc.” and “Di4_B_1, Di4_B_2, Di4_B_3, etc.”, respectively.
- 4.
-
To run auto-analysis script for batch processing, paste and run the source code below via the “Process › Batch › Macro” function of Fiji. User also needs to specify the input directory in the opened dialog window as the location where all Di4 images are located.
Source code of auto-analysis script:
-
selectWindow(«Di4_A.tif»);
-
run(«8-bit»);
-
selectWindow(«Di4_B.tif»);
-
run(«8-bit»);
-
imageCalculator(«Subtract create 32-bit stack», » Di4_A.tif»,» Di4_B.tif»);
-
selectWindow(«Result of Di4_A.tif»);
-
imageCalculator(«Add create 32-bit», «Di4_B.tif»,»Di4_A.tif»);
-
selectWindow(«Result of Di4_B.tif»);
-
imageCalculator(«Divide create 32-bit», » Result of Di4_A.tif»,»Result of Di4_B.tif»);
-
selectWindow(«Result of Di4_A.tif»);
-
close();
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/S0091679X2200098X
Synaptic transmission
Paul Johns BSc BM MSc FRCPath, in Clinical Neuroscience, 2014
Release of neurotransmitter (Fig. 7.4)
Once loaded with neurotransmitter, synaptic vesicles are docked at the presynaptic membrane awaiting release. Docking takes place at active zones. These consist of multi-protein complexes that tether the synaptic vesicle to the presynaptic membrane and contain high concentrations of voltage-gated calcium channels. This leads to brisk calcium influx in response to axon terminal depolarization, since the concentration of calcium is 10,000 times higher in the extracellular fluid. The focal rise in free calcium causes a number of synaptic vesicles to fuse with the presynaptic membrane, emptying their contents into the synaptic cleft by exocytosis.
Release is said to be ‘packeted’ (or quantized) since the total amount of transmitter entering the synaptic cleft is a whole-number multiple of the amount stored in a single vesicle. Transmitter release is regulated by presynaptic autoreceptors which exert negative feedback.
Mechanism of membrane fusion
The mechanism by which synaptic vesicles fuse with the presynaptic membrane is complex and relies on a group of proteins belonging to the SNARE (SNAP receptor) family. The vesicle and presynaptic membranes ‘kiss’ and the interaction between complimentary proteins creates a small fusion pore. This quickly expands as the lipid membranes unite to form a large opening through which the contents of the vesicle are discharged into the synaptic cleft. After exocytosis the vesicle membrane thus becomes part of the presynaptic membrane. However, a similar amount of membrane is reclaimed from the axon terminal to make new vesicles, so there is no net increase in the size of the axon terminal.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780443103216000072
Advances in Cellular Neurobiology
D.G. Jones, in Advances in Cellular Neurobiology, 1983
B Sequences of Synaptogenesis
The respective sequences of appearance of the pre- and postsynaptic membranes, and the differential roles of the membrane thickenings and synaptic vesicles in the elaboration of the presynaptic contact area are depicted in Fig. 80.

Fig. 8. Five alternative schemes by which synaptic junctions may develop. For details, see text.
When both pre- and postsynaptic densities are present early in development, as in Fig. 8a, they appear as continuous plaques (Aghajanian and Bloom, 1967; Jones and Revell, 1970a,b) which, later in development, undergo gradual differentiation and focalization to produce the typical paramembranous densities of the adult synaptic junction. Early workers repeatedly stressed the presence of membrane thickenings prior to the appearance of synaptic vesicles (Glees and Sheppard, 1964; Hamori and Dyachkova, 1964; Meller, 1964; Bunge et al., 1967; Larramendi, 1969; Tennyson, 1970; Alley, 1973; Stelzner et al., 1973). In all these instances, however, caution is necessary to ensure that the membrane specializations actually are precursors of synaptic junctions rather than early components of desmosome-like contacts. This also applies to the idea that puncta adhaerenta are precursors of synaptic junctions (McGraw and McLaughlin, 1980).
An alternative, therefore, is Fig. 8b with synaptic vesicles preceding membrane thickenings. While the presence of presynaptically located vesicles may not constitute an obligatory feature of immature synapses, it remains a helpful one and should not be lightly discarded. Unfortunately, once the presence of vesicles becomes an essential diagnostic feature of early synapses, it also to some extent prejudges the question of whether vesicles or membrane thickenings appear first in synaptic development.
A third alternative is Fig. 8c, in which increases in vesicle numbers and maturation of the paramembranous densities are simultaneous events (Ochi, 1967; Bodian, 1968; Jones and Revell, 1970a,b). For instance, Jones and Revell noted in rat cerebral cortex that vesicles are present when both synaptic membranes are plaquelike and undifferentiated, suggesting that synaptic vesicles appear with (or even after) undifferentiated membrane thickenings but before their differentiation into recognizable dense projections. Comparable observations have been made in other situations, including chick retina (McLaughlin, 1976) and kitten spinal trigeminal nucleus (Dunn and Westrum, 1978).
The discussion this far has assumed that the pre- and postsynaptic membranes appear concurrently (Aghajanian and Bloom, 1967; Jones and Revell, 1970a,b; Adinolfi, 1972a,b; Bloom, 1972). However, this is far from a universal phenomenon. In some studies, as shown in Fig. 8d, the postsynaptic density appears as a well-established entity before the presynaptic region with its dense projections (Poppe et al., 1973; Jones et al., 1974; West and Del Cerro, 1976; Hinds and Hinds, 1976). This phenomenon is best seen in E-PTA-stained material and has so far been described in developing guinea pig cerebral cortex (Jones et al., 1974) and in some synaptic junctions in fetal rat cerebellum (West and Del Cerro, 1976). It is possible that this appearance reflects features of the E-PTA staining reaction, in that it is not staining the presynaptic components of at least some of the immature synapses. Even if this is the case, however, other evidence points to the legitimacy of “naked” postsynaptic sites during early neuronal development.
The converse arrangement, whereby the presynaptic membrane appears prior to the postsynaptic, is far less frequently encountered (Fig. 8e). A commonly quoted illustration of this is the membrane-vesicle clusters described in the cervical spinal cord of Xenopus laevis embryos (Hayes and Roberts, 1973). A similar sequence of events has been described as part of the pattern for photoreceptor synaptogenesis in developing chick retina (McLaughlin, 1976).
These diverse developmental patterns reflect the range of ways by which pre- and postsynaptic junctional elements establish their adult organization. It is not known whether a particular pattern characterizes a particular synaptic population, or whether similar synapses are put together in different ways under differing environmental pressures.
It has long been argued that axodendritic synapses are formed prior to axo-somatic ones (Voeller et al., 1963; Jones, 1975), the synaptic vesicles being spherical rather than flattened (Oppenheim and Foelix, 1972). More recent studies substantiate these general principles in a wide range of situations, e.g., chick optic tectum (McGraw and McLaughlin, 1980), human cervical spinal cord (Okado, 1980), and mouse spinal motor neurons (Vaughn et al., 1977). Occasional exceptions do occur however, including the study of Smolen and Raisman (1980) on rat superior cervical ganglion. In this instance, axosomatic synapses are present transiently for the first 2 weeks of life. It is possible that such synapses may facilitate dendritic development by providing directive cues to the location of the developing synaptogenic field (Vaughn et al., 1977).
Early axodendritic synapses may be of two types: those with asymmetrical junctions and spherical vesicles, and those with symmetrical junctions and spherical vesicles (Devon and Jones, 1981). Neocortical development is characterized by a decrease in the symmetrical/spherical group. This trend may represent a developmental continuum of symmetrical/spherical to asymmetrical/spherical synapses (Bunge et al., 1967; Cragg, 1972; Hinds and Hinds, 1976; Dunn and Westrum, 1978), although the possibility that the asymmetrical/axodendritic terminals represent an intermediate synaptic type cannot be excluded (Adams and Jones, 1982a). The recognition of the symmetrical/spherical terminal type may provide a means of distinguishing immature terminals in adult, and especially in retarded adult, cortical tissue (Devon and Jones, 1981). In this connection it would be interesting to know whether these symmetrical/spherical terminals correspond to the immature type E synaptic junction described by Dyson and Jones (1976a) in E-PTA-stained cortical material.
Terminals with spherical vesicles are repeatedly described as being the first to appear, with flattened or pleomorphic vesicles appearing in later-developing terminals, e.g., in chick optic tectum (McGraw and McLaughlin, 1980), human cervical spinal cord (Okado, 1980), and rabbit superior colliculus and visual cortex (Mathers et al., 1978).
Quantitative approaches to the morphological categorization of developing synaptic junctions are currently confined to the characterization of E-PTA-stained junctions. The quantitative morphogenetic scheme of Dyson and Jones (1976a) assigns synaptic junctions to five categories, A–E, on the basis of variations in the organization of their presynaptic densities. Of these descriptive characteristics, type A represents a mature junctional form with well-defined and discrete dense projections, whereas, at the other end of the scale, type E is an immature form lacking recognizable presynaptic densities; types B–D probably represent intermediate forms (Fig. 3).
The approach to synaptogenesis illustrated by this study gives some idea of the manner in which synaptic ultrastructure can be traced at varying stages during maturation and adult life. By permitting quantitation of the contribution a particular morphogenetic form makes to the population at developmental intervals, it provides a model system for assessing the maturity of both individual junctions and synaptic populations.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780120083046500129
Drosophila model of amyotrophic lateral sclerosis targeting FUS and ubiquilin
Masamitsu Yamaguchi, … Hideki Yoshida, in Handbook of Animal Models in Neurological Disorders, 2023
Mini-dictionary of terms
Active zone, a specialized area on the presynaptic membrane of a nerve cell at which synaptic vesicles fuse to release neurotransmitters into the synaptic cleft and transmit signals to the next synapse or muscle.
Amyotrophic lateral sclerosis, a rare neurodegenerative disease that primarily affects the neurons responsible for controlling voluntary muscle movement.
D. melanogaster, a species of fly in the family Drosophilidae.
Frontotemporal lobar degeneration, a clinically and pathologically heterogeneous syndrome, characterized by progressive declines in behavior or language associated with the degeneration of the frontal and anterior temporal lobes.
FUS, the FET family of RNA-binding proteins that regulate gene expression, genomic integrity, and RNA processing.
GAL4-UAS-targeted expression system, a system using the yeast GAL4 protein that interacts with UAS to induce the expression of genes from any organism in tissue- and temporal-specific manners.
Long non-coding RNA, a group of non-coding RNAs longer than 200 nucleotides.
Neuromuscular junction, a synaptic connection between the terminal end of a motor nerve and a muscle.
RNA interference, a post-transcriptional genetic mechanism of eukaryotes that suppresses gene expression and in which double-stranded RNA cleaved into small fragments initiates the degradation of a complementary mRNA sequence.
Ubiquilin, a protein that regulates protein homeostasis (proteostasis).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780323898331000331
Pharmacology for the Interventional Pain Physician
B. Todd Sitzman M.D., M.P.H., … Honorio Benzon M.D., in Essentials of Pain Medicine and Regional Anesthesia (Second Edition), 2005
Structure and Mechanism of Action of BTs:
BTs are synthesized as single-chain polypeptides and are activated by proteolytic enzymes in a cleaving process (Fig. 18-4). The cleaved heavy chain is responsible for high-affinity docking of the neurotoxin to the presynaptic nerve terminal receptor, enabling the internalization of the bound toxin into the cell.50 The light chain is a zinc-dependent endopeptidase that cleaves membrane proteins responsible for docking acetylcholine vesicles on the inner side of the nerve terminal membrane. The cleavage of these proteins irreversibly precludes fusion of the vesicles with the nerve membrane, thereby preventing release of neurotransmitters into the neuromuscular junction.51
Type A BT appears to be the most potent of the subtypes, and, when injected clinically, has the longest duration of action. While type B and F have limited clinical use, and others are the subject of further study, the multiple differences thus far observed suggest that the subtypes are not interchangeable.
Pharmacological effect of BTs occurs in three stages: binding, internalization, and proteolysis.52
BINDING:
The binding of BTs to the motor endplate presynaptic membrane is a two-stage process.53,54 The C-terminal region of the heavy chain binds in a serotype-specific manner to receptors on the axon terminals of cholinergic neurons. Binding is irreversible and is independent of nerve activity.55 Specific gangliosides have been proposed as the receptors, as well as specific proteins.56,57
INTERNALIZATION:
Binding of the neurotoxin induces the formation of an endosome that carries the toxin into the axon terminal. Internalization of the bound toxin occurs by receptor-mediated endocytosis. Once formed, the contents of the endosome become increasingly acidic, most likely by normal cellular mechanisms. The decrease in pH within the endosome prompts a configurational change in the toxin, which then forms a channel through the membrane. The channel allows all or part of the toxin to enter the cytosol.58 This process is pH dependent.
PROTEOLYSIS (FIG. 18-5):
The exceptional potency of the botulinum neurotoxins has long suggested that a catalytic effect is involved. In the cytosol the proteolytic effects occur. BT types A, E, and C cleave synaptosome-associated protein-25 (SNAP-25). Types B, D, F, and G cleave the synaptic protein synaptobrevin, also known as vesicle-associated membrane protein (VAMP). Type C also cleaves syntaxin.59 Each of these protein substrates participates in the formation of the exocytotic SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) complex, which is essential for fusion of acetylcholine-containing vesicles with the presynaptic membrane, a prerequisite for acetylcholine release. SNAREs form coiled bundles that bridge the membrane of the synaptic vesicle with the plasma membrane by an interaction between VAMP, which is anchored in the vesicle membrane, and syntaxin, which is anchored in the plasma membrane, perhaps preceded by an interaction between VAMP and SNAP-25.60
The effects of the botulinum neurotoxins are due to irreversible inhibition of the release of acetylcholine from cholinergic nerve terminals, including those of motor neurons, preganglionic sympathetic and parasympathetic neurons, and postganglionic parasympathetic nerves.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780443066511500228
Cytology of the nervous system
Jahangir Moini MD, MPH, … Mohtashem Samsam MD, PhD, in Epidemiology of Brain and Spinal Tumors, 2021
Synapses
Effective messages move along an axon and are transferred to another cell, occurring at a synapse, a specialized area where a neuron communicates with another cell. Synapses are also referred to as synaptic junctions. At a synapse, information moves from the presynaptic neuron to the postsynaptic neuron. Synapses can involve various postsynaptic cells. An example is the neuromuscular junction, a synapse in which the postsynaptic cell is a skeletal muscle fiber. There are more than 100 trillion synapses in the human brain alone. The types of synapses in the CNS are summarized in Table 2.4.
Table 2.4. Types of synapses in the central nervous system.
| Type | Presynaptic Structure | Postsynaptic Structure | Functions |
|---|---|---|---|
| Axodendritic | Axon terminal | Dendrite | Mostly excitatory |
| Axosomatic | Axon terminal | Cell body | Mostly inhibitory |
| Axoaxonic | Axon terminal | Axon terminal | Presynaptic inhibition, which regulates transmitter release in the postsynaptic axon |
| Dendrodendritic | Dendrite | Dendrite | Local interactions that can be excitatory or inhibitory, in neurons that lack axons, such as the retina |
Structure
A synapse consists of three components: the presynaptic membrane formed by the terminal button of an axon, the postsynaptic membrane formed from a segment of dendrite or cell body, and the space in between these two structures, the synaptic cleft. Some neural cells have up to two hundred thousand synaptic connections.
Classification
There are two types of synapses: electrical and chemical, with unique functions. At electrical synapses, there is direct physical contact between cells. Presynaptic and postsynaptic membranes of cells are joined at gap junctions (see Fig. 2.6A). Lipid areas of nearby membranes are separated by just two nanometers (nm), kept in position by essential membrane proteins called connexons. Pores formed by these proteins allow ions to pass between the cells. Changes in membrane potential of one cell cause local currents to affect another. An electrical synapse propagates action potentials between cells efficiently and quickly because of this. In adults, electrical synapses are not common in the CNS or PNS. They occur in brain areas such as the vestibular nuclei, involved in balance, also in the eyes, and in one or several pairs of PNS ganglia (ciliary ganglia). They are also present in embryonic structures.

Figure 2.6. (A) Electrical synapse; (B) Chemical synapse.
In a chemical synapse (Fig. 2.6B), one neuron signals another. This uses the axon terminal of the presynaptic neuron, which sends the message, and the postsynaptic neuron, which receives it. The cells are separated by the narrow synaptic cleft. A presynaptic cell is most often a neuron. Specialized receptor cells create synaptic connections with dendrites. Postsynaptic cells are neurons or other forms of cells. Communications between neurons occur at synapses on dendrites, cell bodies, or along axons of receiving cells. Axoaxonic synapses are between axons of two neurons. Axosomatic synapses have junctions, at an axon terminal of a neuron, and the cell body of another neuron. In an axodendritic synapse, synaptic contact is between an axon terminal of one neuron and a dendrite of another. Chemical synapses are very common compared to electrical synapses. Communications across chemical synapses occur from presynaptic membranes to postsynaptic membranes, and not in the reverse direction.
A neuromuscular junction is a synapse between a neuron and a skeletal muscle cell. At a neuroglandular junction, a neuron regulates activities of a secretory cell. Neurons innervate many other types of cells, such as adipocytes. Axon terminals of presynaptic cells release neurotransmitters into the synaptic cleft. The neurotransmitters are within synaptic vesicles in the axon terminal. Synaptic vesicles contain a small collection, or quanta, of neurotransmitter. When the terminal is depolarized from an action potential of its “parent” axon, there is a calcium influx that leads to phosphorylation of proteins called synapsins. After this phosphorylation, the vesicle links to the presynaptic membrane that faces the synaptic cleft, and the neurotransmitter is released. Another method of neurotransmitter release is based on transporter molecules, which usually act by taking up neurotransmitters from the synaptic cleft.
Each postsynaptic cell has its own type of axon terminal. A round axon terminal is present if a postsynaptic cell is another neuron. At a neuromuscular junction, the axon terminal is of much more complex. Axon terminal structures include mitochondria and thousands of vesicles containing neurotransmitters. Axon terminals reabsorb breakdown molecules of neurotransmitters at the synapse, then reproduce the neurotransmitters. Axon terminals also constantly receive neurotransmitters created by the cell body, along with enzymes and lysosomes, via anterograde flow.
Function of electrical synapses
In electrical synapses, signal transmission occurs as electrical signals, without using molecules. Signals are not modified during transmission. Electrical signals pass over gap junctions. The space between presynaptic and postsynaptic neurons is very small. Signal transmission occurs in both directions. It happens without use of energy and is therefore passive. Signal transmission via electrical synapses is very fast.
Function of chemical synapses
Electrical events trigger release of neurotransmitter release, flooding the synaptic cleft, and binding to receptors on the postsynaptic plasma membrane. This changes membrane permeability, producing graded potentials. The process is similar to operation of neuromuscular junctions. Chemical synapses do not use direct cellular joining, so there is more variation in results. At a chemical synapse, arriving action potentials may or may not release enough neurotransmitter to bring the postsynaptic neuron to threshold.
Cholinergic synapses release acetylcholine (ACh) at all neuromuscular junctions involving skeletal muscle fibers. They release ACh at many CNS synapses, all PNS neuron-to-neuron synapses, and all neuromuscular and neuroglandular junctions in the parasympathetic nervous system. At cholinergic synapses between neurons, a presynaptic cleft lies between presynaptic and postsynaptic membranes. Most ACh in an axon terminal collects in synaptic vesicles, each having thousands of neurotransmitter molecules. One axon terminal may contain a million such vesicles. When an action potential arrives at the presynaptic axon terminal, it depolarizes the membrane, opening its voltage-gated calcium ion channels for a short time. Extracellular calcium ions enter through these calcium channels. The ions attach to the vesicles containing ACh. Attachment of calcium ions to the vesicles causes release of ACh in the synaptic cleft. The ACh is released in groups of nearly 3000 molecules, the average number of molecules in just one vesicle. Release of ACh stops quickly, since active transport removes calcium ions quickly from cytoplasm, in the axon terminal, back to the extracellular space. Ions are pumped out of the cell, or moved to the mitochondria, awaiting another action potential.
ACh binds to receptors on the postsynaptic membrane, depolarizing it. Across the synaptic cleft, ACh diffuses to postsynaptic membrane receptors. The ACh receptors have chemically gated sodium and potassium ion channels. The main response is increased permeability to sodium ions, causing a depolarization in the postsynaptic membrane of about 20 msec. The cation channels move potassium ions outward from the cell. Sodium ions are moved by a stronger electrochemical gradient. A slight depolarization of the postsynaptic membrane occurs, which is a graded potential. The more ACh released at the presynaptic membrane, the more open cation channels in the postsynaptic membrane. Therefore, there is more depolarization. If depolarization brings an adjoining section of excitable membrane such as the initial axon segment to threshold, an action potential occurs in the postsynaptic neuron. The enzyme Acetylcholinesterase removes ACh from the synaptic cleft. The effects of ACh on the postsynaptic membrane are temporary because acetylcholinesterase (also called AChE or cholinesterase) is contained in the synaptic cleft and postsynaptic membrane. About half of all ACh released at the presynaptic membrane is degraded before reaching the postsynaptic membrane receptors. Only about 20 msec are needed for ACh molecules that bind to receptor sites to be broken down. Through hydrolysis, AChE breaks down ACh into acetate and choline. The water-soluble, vitamin-like nutrient choline is easily absorbed by axon terminals, then used to synthesize more ACh, via acetate provided by coenzyme A (CoA). Coenzymes from vitamins are needed for many enzymatic reactions. Acetate moving away from the synapse can be absorbed and metabolized by postsynaptic cells, or by various cells and tissues.
Synaptic delay
A synaptic delay is the time needed for a signal to cross a synapse between two neurons. There is only 0.2−0.5 msec between arrival of an action potential at the axon terminal and its effect upon the postsynaptic membrane. Most of the delay is due to the time needed for calcium ion influx and release of the neurotransmitters. The delay is not due to neurotransmitter diffusion. The synaptic cleft is thin, and neurotransmitters diffuse across it quickly.
If a delay of 0.5 msec occurs, an action potential can travel over 7 cm (about 3 inches) along a myelinated axon. When information is passed down CNS interneurons, increased synaptic delay may exceed propagation time along the axons. When fewer synapses are involved, cumulative synaptic delay is shorter, and responses are faster. The quickest reflexes have one synapse, with a sensory neuron directly controlling a motor neuron.
Synaptic fatigue
Synaptic fatigue, or short-term synaptic depression, is a temporary inability of neurons to fire, and to transmit input signals. Synaptic fatigue is a type of synaptic plasticity, a form of negative feedback. It is mostly presynaptic in its function. Since ACh molecules are recycled, axon terminals do not completely depend upon ACh delivered through axonal transport from the cell body. With intense stimulation, resynthesis and transport mechanisms may be unable to handle neurotransmitter demands. Then, synaptic fatigue occurs. The response of the synapse is weakened until ACh is replenished.
Point to remember
Diphenylhydantoin, antidepressants classified as selective serotonin reuptake inhibitors (SSRIs), and caffeine can affect synaptic transmission. Diphenylhydantoin reduces frequency of action potentials reaching the axon terminal. The SSRIs block serotonin transport into presynaptic cells, increasing stimulation of postsynaptic cells. Caffeine stimulates activity of the nervous system by lowering synaptic thresholds, causing postsynaptic neurons to be excited more easily.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128217368000121
Nicotine
Tursun Alkam, … Toshitaka Nabeshima, in Reference Module in Biomedical Sciences, 2022
Future directions
Nicotine’s toxicity is mediated by nAChR subunits located in pre- or postsynaptic membranes. In addition to membrane internalization, nAChRs have a half-life of up to 10 days when membrane-bound (Benowitz et al., 2009; Benowitz et al., 1982). Nicotine exerts its various toxicities through different subunits of nAChRs, which are located in the pre- or post-synaptic membranes. Nicotine increases the expression and half-life of surface membrane nAChRs (Kuryatov et al., 2005; Nashmi et al., 2003). However, the process or mechanism involved in the degradation or elimination of nAChRs-binding or free nicotine in the synaptic cleft, as well as its half-life, is unknown. The role of nAChRs subunits in nicotine intoxication in various nervous systems is unclear. Future research should focus on synaptic nicotine degradation and nAChR subunits involved in nicotine toxicity.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128243152001482
Assembly of Synapses in the Vertebrate Central Nervous System
LEORA GOLLAN, PETER SCHEIFFELE, in Protein Trafficking in Neurons, 2007
C Nucleation of Synaptic Scaffolds by Adhesion Molecules
The mechanical stability of synapses and the precise apposition of pre- and postsynaptic membrane domains implies that adhesion molecules might bridge the synaptic cleft and thereby keep both sides of the synapse in register. Ultrastructural studies revealed the presence of different sites of cell-cell adhesion at the synapse—including the linkage across the synaptic cleft, as well as at puncta adherentia that flank the synaptic cleft. A third site of cellular adhesion at central synapses are the end feet of glial cells that tightly enclose the synaptic junctions.
Most attention in recent years has been focused on neuronal adhesion molecules that directly bridge the synaptic cleft and that might instruct the assembly of synaptic structures. Several such adhesion molecules have been identified. The synaptogenic activity of these adhesive factors is thought to rely on the nucleation of cytoplasmic scaffolds at the pre- and postsynaptic side, which subsequently facilitate the recruitment of other cytoplasmic and cell surface molecules to an incipient synaptic connection.
In vitro experiments with cultured neurons revealed that exposing axonal or dendritic processes to specific adhesive triggers is sufficient to induce pre- and postsynaptic structures. One extensively studied adhesion system with synapse-inducing function is the neuroligin-neurexin complex. Neuroligins and neurexins are heterophilic adhesion proteins that form a trans-synaptic complex (Ichtchenko et al. 1995; Nguyen and Südhof 1997; Song et al. 1999). Cytoplasmic sequences in neuroligin-1 target this protein exclusively to the synaptic sites in dendrites and ensure exclusion from the axons (Dresbach et al. 2004; Iida et al. 2004; Rosales et al. 2005). This strict polarity of neuroligin-1 provides this heterophilic adhesion system with the required directionality for the assembly of asymmetric synaptic structures. Contact of axons with neuroligin-1 ectopically expressed in a nonneuronal cell induces clustering of axonal neurexins and the cytoplasmic scaffolding molecule CASK (Scheiffele et al. 2000; Dean et al. 2003). These neuroligin-induced axonal contacts then mature into functional presynaptic release sites, including active zone components and clusters of synaptic vesicles. Conversely, contact of dendrites with neurexin-1 expressing cells leads to the formation of neuroligin clusters and postsynaptic structures, such as the accumulation of the scaffolding protein PSD95 and NMDA receptors (Graf et al. 2004). These experiments suggest that the neuroligin-neurexin adhesion system can nucleate pre- and postsynaptic structures. To a large extent these structures are thought to assemble around scaffolding components that bind to the cytoplasmic tails of neuroligins and neurexins (see Figure 4.1). For example, postsynaptic neuroligin-1 binds to PSD95, which in turn can recruit NMDA-receptors and other scaffolding components such as GKAP, Homer, and Shank (Irie et al. 1997; Meyer et al. 2004; Prange et al. 2004; Chih et al. 2005).
Interestingly, the neuroligin-2 isoform, which differs in its cytoplasmic sequences from neuroligin-1, is found primarily at GABAergic synapses. Extracellular aggregation of neuroligin-2 results in not only the recruitment of PSD95, but also of gephrin, a scaffolding protein specific to inhibitory synapses (Graf et al. 2004; Varoqueaux et al. 2004). Selective interaction of excitatory and inhibitory postsynaptic scaffolding proteins with different neuroligin cytoplasmic tail sequences therefore might contribute to the nucleation of specific postsynaptic scaffolds. However, interactions between extracellular domains that may contribute to this process have not been ruled out.
Similar experiments revealed synaptogenic activities for the cell adhesion molecule SynCAM, a homophilic Ig-domain protein (Biederer et al. 2002). Interestingly, SynCAM and neuroligin-1 differ in their cytoplasmic interactions with scaffolding proteins. These differences may explain the differential recruitment of NMDA and AMPA-type glutamate receptors to neuroligin-1- and SynCAM-induced synapses, respectively (Sara et al. 2005). Neuroligin-1 recruits PSD95 and NMDA-receptors through the type I PDZ-binding motif in neuroligin, whereas SynCAM contains a type II PDZ-binding motif that contributes to the recruitment of functional AMPA-receptors to synapses. The importance of these selective scaffolding interactions was confirmed in experiments showing that a chimaeric protein, which includes neuroligin-1 containing the cytoplasmic sequences of SynCAM, is capable of stimulating AMPA receptor recruitment (Sara et al. 2005).
Another presumptive cell adhesion molecule that has been specifically linked to the synaptic recruitment of AMPA receptors through interaction with scaffolding molecules is Dasml (Shi et al. 2004). However, the extracellular binding partners of this protein are currently unknown.
It is likely that several adhesion molecules that act through mechanisms similar to the neuroligin-neurexin complex and SynCAM will cooperate in the nucleation of synaptic scaffolds and the subsequent recruitment of neurotransmitter receptors. Candidate molecules include the nectins, a family of homophilic Ig-domain proteins that are similar to SynCAM (Satoh-Horikawa et al. 2000; Mizoguchi et al. 2002). The integration of multiple adhesive signals likely occurs at the level of the cytoplasmic scaffold. There is substantial crosstalk between different scaffolding components; for example, the CASK/Mint/MALS complex that binds to neurexins is linked to beta-catenin, the cytoplasmic binding partner of cadherins (see Figure 4.1; Hata et al. 1996; Perego et al. 2000; Bamji et al. 2003). Such crosstalk might be employed for the sequential recruitment of different adhesive factors to a growing adhesion site as has been observed for cadherins and nectins in epithelial cells (Takahashi et al. 1999; Tachibana et al. 2000).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123694379500074
Neurotransmitter release
Constance Hammond, … Michael Seagar, in Cellular and Molecular Neurophysiology (Fourth Edition), 2015
When does Ca2+ enter the presynaptic element in response to a presynaptic spike?
In order to record the presynaptic Ca2+ current, the presynaptic membrane is clamped at VH = −80 mV by means of two whole-cell electrodes. To isolate the presynaptic Ca2+ current, voltage-dependent Na+ and K+ currents are blocked with TTX and tetraethlyammonium chloride (TEA), respectively. An action potential waveform is injected into the presynaptic terminal through one of the whole-cell electrodes. It evokes a presynaptic Ca2+ current that is recorded by the second whole-cell electrode. Recordings show that Ca2+ influx is tightly associated with the repolarizing phase of the action potential (Figure 7.4): it is essentially a tail current (see Appendix 5.2) that activates shortly after the peak of the action potential and ends before repolarization is complete. It has a peak amplitude of 2.6 ± 0.2 nA and a half-width of about 350 ms. The delay between the beginning of the action potential and that of Ca2+ current is about 500 μs at 23–24°C.

Figure 7.4. Ca2+ current flows into presynaptic terminal during the repolarizing phase of the presynaptic spike.
Two whole-cell electrodes are positioned in the calyx of Held; one measures the membrane potential (V) and the other one injects current (I). It is a two-electrode voltage clamp configuration. The preparation is bathed in 2 mM external Ca2+ with TTX and TEA to block the voltage-gated Na+ and K+ currents. (a) The voltage clamp command (VCmd) is an action potential waveform (V) from a holding potential of −80 mV. A reduced and inverted action potential waveform is also applied, scaled and re-inverted to measure the passive current. Top, recorded voltages. The two action potential waveforms are superimposed. Due to series resistance, the repolarization is somewhat slower during the full action potential than during the scaled action potential. Middle, currents. The current flowing during the full-sized action potential has a larger inward component (labeled ICa). The two passive transients overlay well. Bottom, calcium current. The calcium current is obtained by subtracting the passive current from the current measured during the full action potential. All traces are the average of 11. Vertical dotted lines denote peak of the action potential waveform and of the calcium current. (b) This Ca2+ current is reduced in 1 or 0.5 mM external Ca2+ ([Ca2+ ]o)
Part (a) from Borst JG, Sakmann B (1998) Calcium current during a single action potential in a large presynaptic terminal of the ra t brainstem. J. Physiol. 506, 143–157, with permission. Part (b) adapted from Borst JGG, Sakmann B (1996) Calcium influx and transmitter release in a fast CNS synapse. Nature 383, 431–434, with permission.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123970329000078
The Neuromuscular Junction and Excitation-Contraction Coupling
Joseph Feher, in Quantitative Human Physiology, 2012
Acetylcholine Is Degraded and Then Recycled
Each action potential fuses perhaps 300 or so synaptic vesicles with the presynaptic membrane. Continuous excitation and subsequent fusion of these large numbers of vesicles with the presynaptic membrane would expand the area of the membrane and deplete it of vesicles. To avoid this, the presynaptic cell rapidly recycles the vesicles. Through an elaborate mechanism involving specialized proteins such as clathrin and dynamin, vesicles are pinched off and endocytosed. The empty vesicles are refilled with acetylcholine through a transport channel that exchanges acetylcholine for H+ ions. The H+ ions are accumulated within the vesicles through a vacuolar H-ATPase in the vesicle membrane.
Acetylcholine is also partially recycled. Acetylcholine is degraded to acetate and choline in the synapse through the action of acetylcholinesterase. The acetate is not recycled, but the choline is taken back up into the presynaptic cell through a transporter on the presynaptic cell membrane. In the cell, acetylcholine is resynthesized from acetyl CoA and choline through the enzyme choline acetyltransferase. The events occurring in the axon terminal that initiate and shut off neuromuscular transmission are shown in Figure 3.6.4.

Figure 3.6.4. Release of neurotransmitter at the neuromuscular junction. The action potential propagates along the axon and opens voltage-gated Ca2+ channels, leading to an increase in cytoplasmic [Ca2+] in the terminal. The [Ca2+] binds to proteins that dock synaptic vesicles at the active zone, leading to fusion of the vesicles with the presynaptic membrane and release of neurotransmitter, acetylcholine, into the synapse. ACh diffuses across the gap to bind to ACh receptors located on the postsynaptic membrane. ACh binding to this receptor increases the K+ and Na+ conductance, which in turn depolarizes the muscle cell membrane. This depolarization is called the end-plate potential. It is conveyed passively to a nearby patch of muscle membrane where, if the end-plate potential is sufficient, it initiates an action potential. Vesicle release is shut off by removal of the synaptic Ca2+ by Ca2+ pumps or Na+/Ca2+ exchange. The acetylcholine signal is terminated by hydrolysis of acetylcholine in the synapse by acetylcholinesterase. VDCC=voltage-dependent calcium channel.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123821638000293
Нервная ткань — основная ткань, формирующая нервную систему и создающая условия для реализации ее многочисленных функций. Нервная ткань имеет эктодермальное происхождение, не принято делить нервную ткань на какие-либо виды тканей. Обладает двумя основными свойствами: возбудимостью и проводимостью.
Нейрон
Структурно-функциональной единицей нервной ткани является нейрон (от др.-греч. νεῦρον — волокно, нерв) — клетка с одним
длинным отростком — аксоном (греч. axis — ось), и одним/несколькими короткими — дендритами (греч. dendros — дерево).
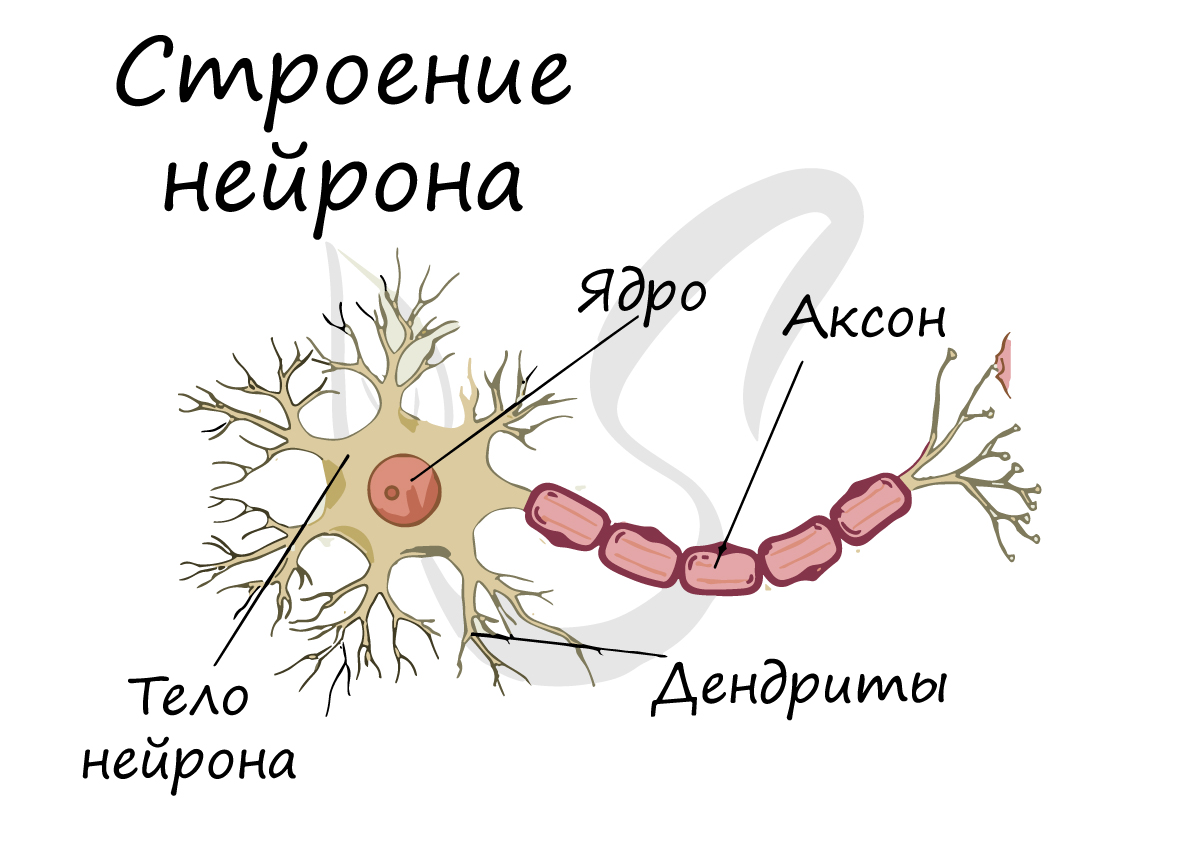
Спешу сообщить, что представление, будто короткий отросток нейрона — всегда дендрит, а длинный — всегда аксон, в корне неверно. С точки
зрения физиологии правильнее дать следующие определения: дендрит — отросток нейрона, по которому нервный импульс перемещается к телу нейрона, аксон — отросток нейрона, по которому импульс перемещается от тела нейрона.
Нейроны обладают 4 свойствами:
- Рецепция (лат. receptio — принятие) — способны воспринимать поступающие сигналы (дендриты)
- В ответ на сигналы способны переходить в состояние возбуждения или торможения
- Проведение возбуждения (от дендрита к телу нейрона, затем — к концу аксона)
- Передача сигнала другим объектам — нейрону или эффекторному органу
В физиологии эффекторным (от лат. efferes — выносящий) органом часто называют исполнительный орган или орган-мишень воздействия (мышцы, железы). Орган-эффектор выполняет те или иные «приказы» ЦНС (центральной нервной системы) или эндокринных желёз
Отростки нейронов проводят нервные импульсы и передают их другим нейронам, эффекторам, благодаря чему
мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
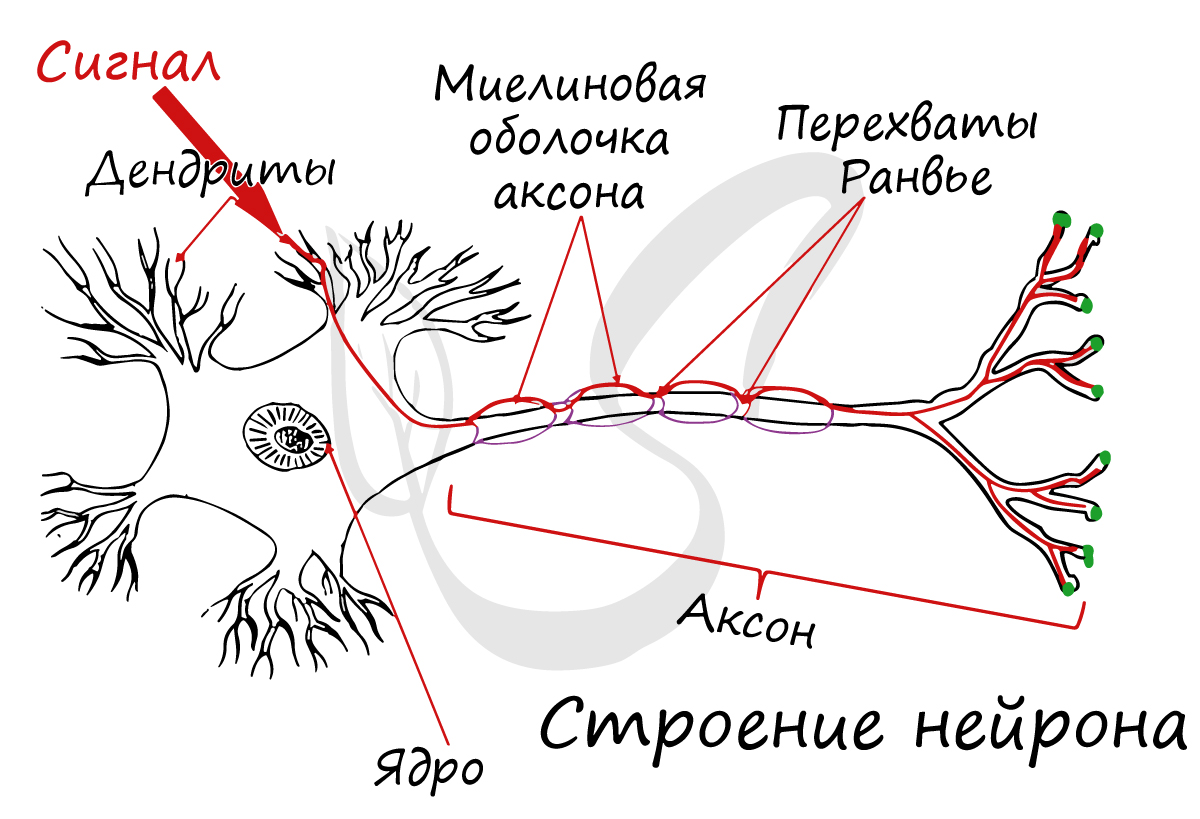
Миелиновая оболочка
Нервные волокна подразделяются на миелиновые и безмиелиновые. Нервное волокно — это один или несколько отростков нейронов (могут быть как аксоны, так и дендриты) с окружающей оболочкой.
Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы (скорость проведения 1-2 м/c). Миелиновые — образуют белое вещество головного и спинного мозга, нервные волокна соматической нервной системы (5-120 м/с).
В миелиновых нервных волокнах отростки нейронов покрыты миелиновой оболочкой (на 70-75% состоит из липидов (жиров)), которая обеспечивает изолированное проведение нервного
импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и,
когда мы хотели сделать движение рукой, то вместе с рукой двигалась бы нога.
Существует болезнь при которой собственные антитела уничтожают миелиновую оболочку нервных волокон головного и спинного мозга (случаются и такие сбои в работе организма). Эта
болезнь — рассеянный склероз, по мере прогрессирования приводит к разрушению не только миелиновой оболочки, но и нервов — а значит,
происходит атрофия мышц и человек постепенно становится обездвиженным.

Миелиновый слой представлен несколькими слоями мембраны глиальной клетки (леммоцит, шванновская клетка), которые закручиваются вокруг осевого цилиндра (отростка нейрона). Это закручивание хорошо видно на картинке, где изображен здоровый нерв, чуть выше 
Миелиновый слой оболочки волокна регулярно прерывается в местах стыка соседних леммоцитов — перехваты Ранвье. Миелиновая оболочка обеспечивает изолированное и более быстрое проведение возбуждения (сальтаторный тип, лат. salto — скачу, прыгаю).

Нейроглия (греч. νεῦρον — волокно, нерв + γλία — клей)
Вы уже убедились, насколько значимы нейроны, их высокая специализация приводит к возникновению особого окружения — нейроглии.
Нейроглия (глиальные клетки, глиоциты) — вспомогательная часть нервной системы, которая выполняет ряд важных функций:
- Опорная — поддерживает нейроны в определенном положении
- Регенераторная (лат. regeneratio — возрождение) — в случае повреждения нервных структур нейроглия способствует регенерации
- Трофическая (греч. trophe — питание) — с помощью нейроглии осуществляется питание нейронов: напрямую с кровью нейроны не контактируют
- Электроизоляционная — леммоциты (шванновские клетки) закручиваются вокруг отростков нейронов и формируют миелиновую оболочку
- Барьерная и защитная — изолируют нейроны от тканей внутренней среды организма
- Некоторые глиоциты секретируют цереброспинальную (спинномозговую) жидкость — ликвор (от лат. liquor — жидкость)
В состав нейроглии входят разные клетки, их в десятки раз больше чем самих нейронов. В периферическом отделе нервной
системы миелиновая оболочка, изученная нами, образуется именно из нейроглии — шванновских клеток (леммоцитов). Между ними хорошо
заметны перехваты Ранвье — участки, лишенные миелиновой оболочки, между двумя смежными шванновскими клетками.
Классификация нейронов
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
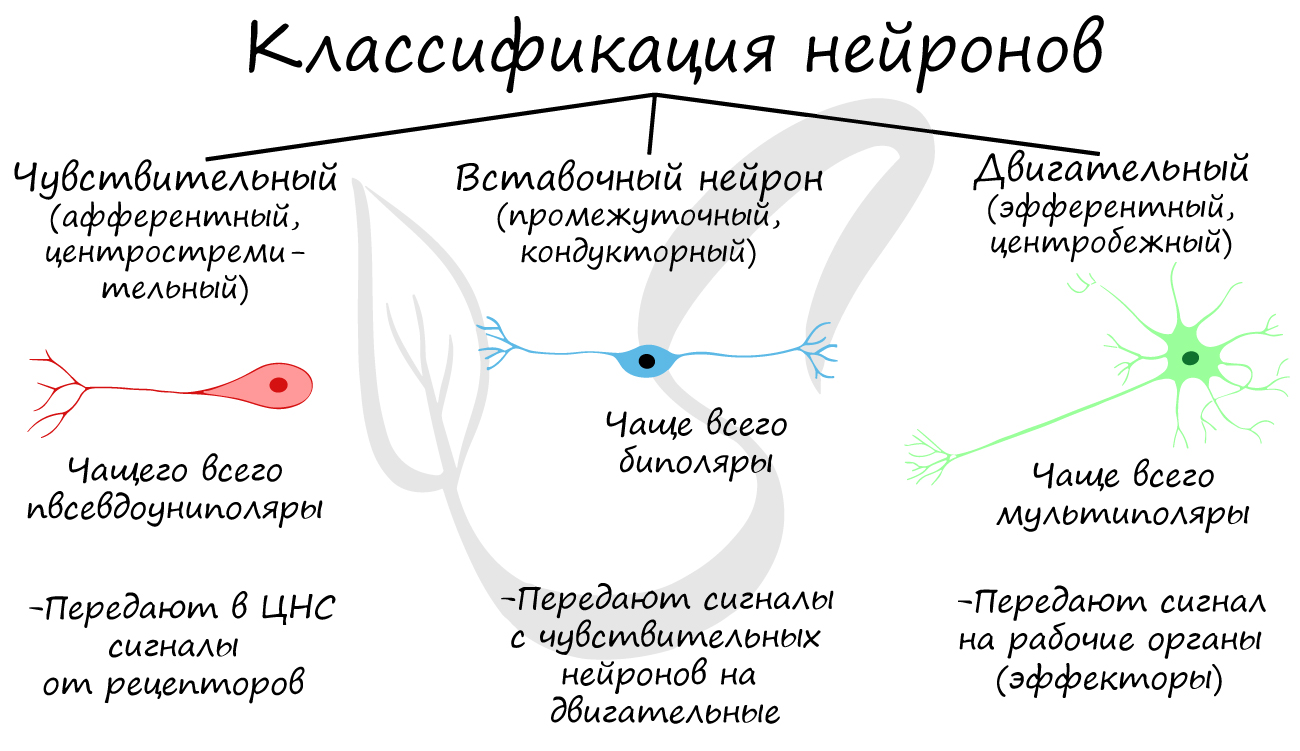
Чувствительные нейроны также называются афферентные, центростремительные, сенсорные, воспринимающие — они воспринимают раздражения, преобразуют их в нервные импульсы и передают в ЦНС. Рецептором называют концевое окончание чувствительных нервных
волокон, воспринимающих раздражитель.
Вставочные нейроны также называются промежуточные, ассоциативные — они обеспечивают связь между чувствительными и двигательными
нейронами, передают возбуждение в различные отделы ЦНС, участвуют в обработке информации и выработке команд.
Двигательные нейроны по-другому называются эфферентные, центробежные, мотонейроны — они передают нервный импульс (возбуждение) на
эффектор (рабочий орган). Наиболее простой пример взаимодействия нейронов — коленный рефлекс (однако вставочного нейрона
на данной схеме нет). Более подробно рефлекторные дуги и их виды мы изучим в разделе, посвященном нервной системе.
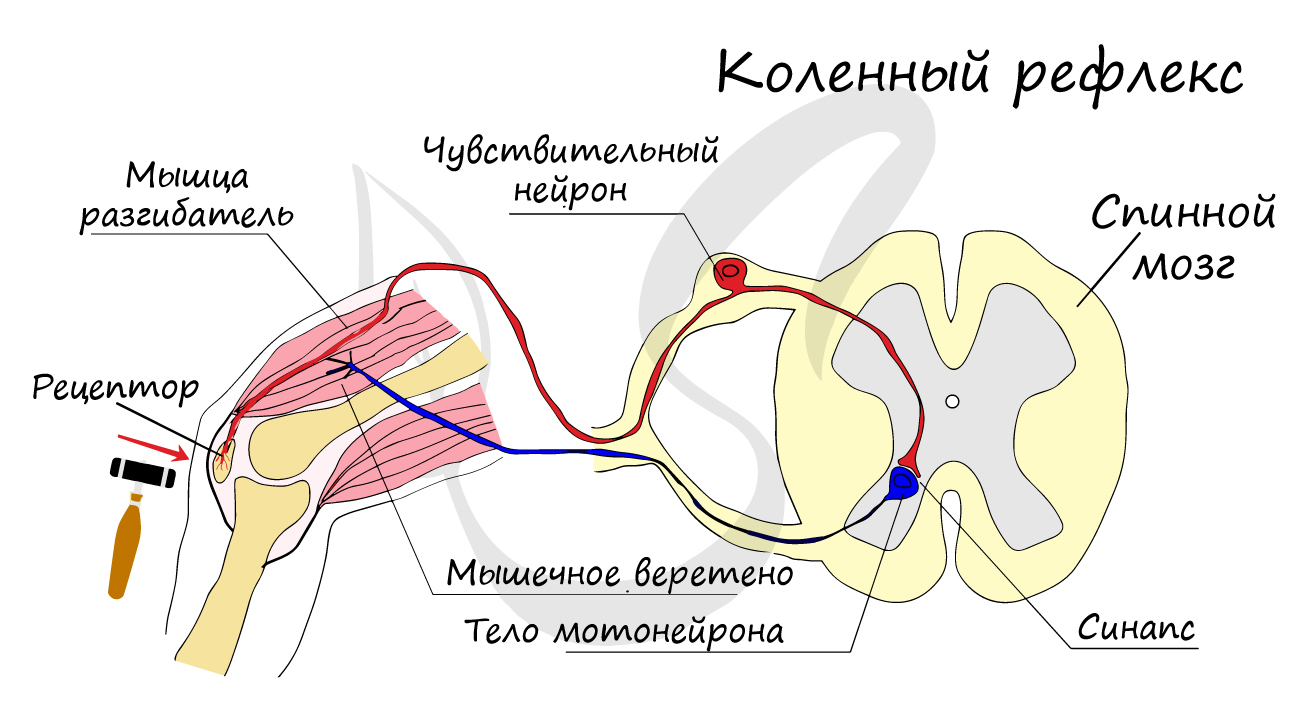
Синапс
На схеме выше вы наверняка заметили новый термин — синапс (греч. sýnapsis — соединение). Синапсом называют место контакта между двумя нейронами или между
нейроном и эффектором (органом-мишенью). В синапсе нервный импульс «преобразуется» в химический: происходит выброс особых
веществ — нейромедиаторов (наиболее известный — ацетилхолин) в синаптическую щель.
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с
нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают
сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
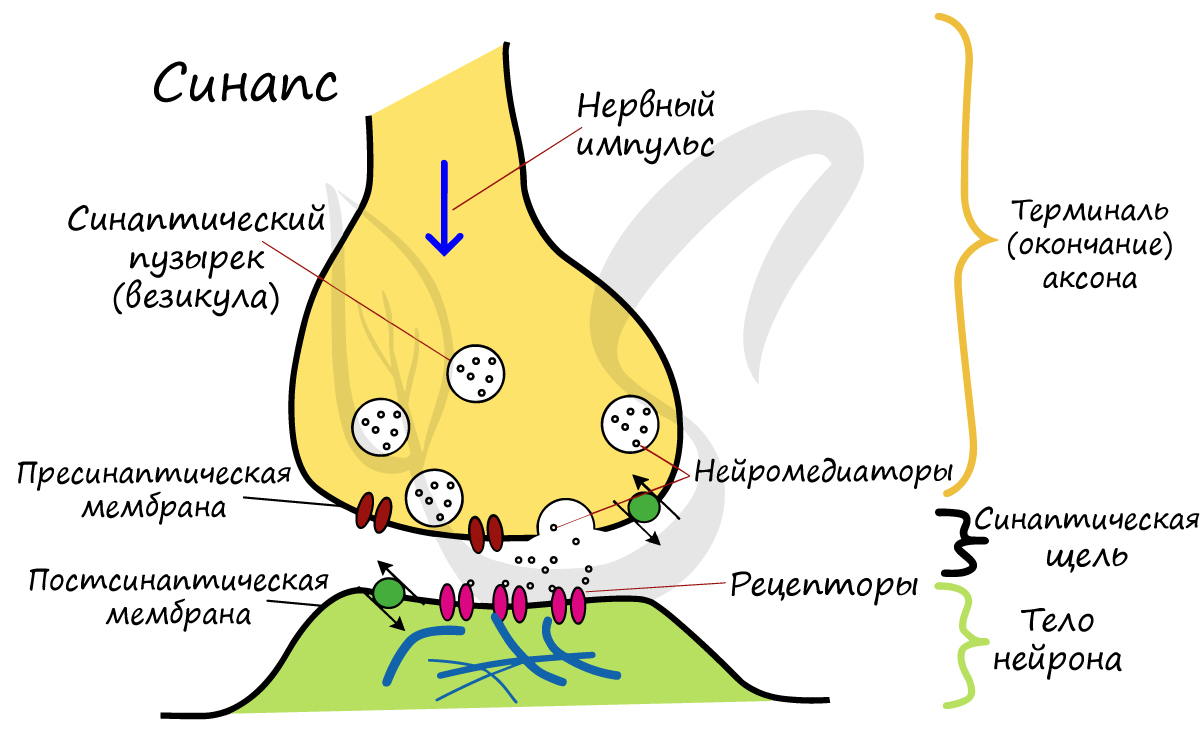
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение (нервный импульс)
передается другому нейрону. Так устроена нервная система: электрический путь передачи сменяется
химическим (в синапсе).
Яд кураре
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими Не могу утаить
историю о яде кураре, который используют индейцы для охоты с древних времен.
Этот яд блокирует ацетилхолиновые рецепторы на постсинаптической мембране, и, как следствие, химическая передача возбуждения с
одного нейрона на другой становится невозможна. Это приводит к тому, что нервные импульсы перестают поступать к эффекторам,
в том числе к дыхательным мышцам (межреберным, диафрагме), вследствие чего дыхание останавливается и наступает смерть животного.

Нервы и нервные узлы
Собираясь вместе, отростки нейронов (нервные волокна) образуют пучки нервных волокон. Нервные пучки объединяются в нервы, которые покрыты соединительнотканной оболочкой.
В случае, если тела нейронов концентрируются в одном месте за пределами центральной нервной системы, их скопления
называют нервным узлом — или ганглием (от др.-греч. γάγγλιον — узел).
В случае сложных соединений между нервными волокнами говорят о нервных сплетениях. Одно из наиболее известных —
плечевое сплетение.
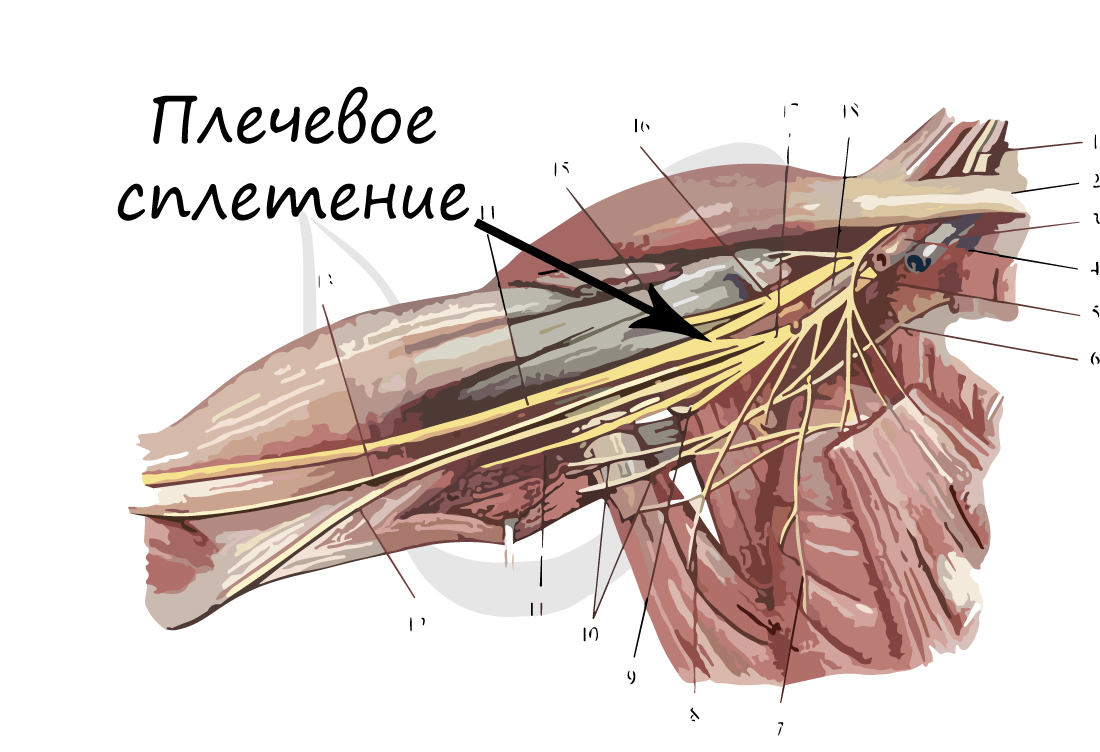
Болезни нервной системы
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения
чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом
движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет
невозможно: возникает паралич, но чувствительность может сохраняться.
Существует тяжелое мышечное заболеванием — миастения (от др.-греч. μῦς — «мышца» и ἀσθένεια — «бессилие, слабость»), при
котором собственные антитела разрушают мотонейроны (двигательные нейроны).

Постепенно любые движения мышцами становятся для пациента все труднее,
становится тяжело долго говорить, повышается утомляемость. Наблюдается характерный симптом — опущение верхнего века.
Болезнь может привести к слабости диафрагмы и дыхательных мышц, вследствие чего дыхание становится невозможным.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Для более глубокого понимания механизма действия БТА, которое будет подробно описано ниже, необходимо вспомнить строение нервно-мышечного синапса.
Синапс (греч. synapsis – соединение) – это специализированная структура, обеспечивающая передачу возбуждающих или тормозящих влияний между двумя возбудимыми клетками.
Синапсы разделяют по виду соединяемых клеток, по эффекту, по природе медиатора (рис. 2).
Нервно-мышечный синапс образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры. Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и синаптическая мембрана, являющаяся частью плазматической мембраны постсинаптической мышечной клетки. Пресинаптической мембраной называют часть непокрытой миелином мембраны аксонной терминали, обращенной в синаптическую щель.
Пресинаптическая терминаль формируется тонкой веточкой аксона, которая, подходя к мышечному волокну, образует утолщение. В пресинаптической терминали находятся синаптические пузырьки (везикулы), содержащие запасы медиатора ацетилхолина. Везикул может быть до нескольких тысяч. Их диаметр около 40 нм, и в каждой содержится несколько тысяч молекул медиатора. Непосредственно около пресинаптической мембраны находится около 1000 везикул – оперативный запас, готовый к выделению в пресинаптическую щель. Остальная часть является мобильным запасом, готовым к быстрому перемещению к мембране взамен освободившегося медиатора. Наиболее существенным является стационарный резерв (около 300 тыс. везикул). Он находится в проксимальных отделах терминали. Освобождение медиатора в синаптическую щель происходит порциями. Минимальная порция – везикула медиатора, которая содержит около 5–10 тыс. молекул ацетилхолина и называется квантом (рис. 3).
Пресинаптическая мембрана
Cтраница 1
Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других истончена и имеет отверстия для сообщения цитоплазмы аксона с синаптическим пространством. Постсинапти-ческая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
[1]
В области пресинаптической мембраны располагаются синаптические пузырьки, в которых находятся молекулы медиаторов. Количество и расположение пузырьков зависит от типа клеток, между которыми передается нервный импульс.
[2]
Ацетилхолин высвобождается квантами в зависимости от потенциала действия пресинаптической мембраны. Выделению ацетилхолина способствуют ионы Са2, находящиеся во внутриклеточной жидкости. При деполяризации пресинаптической мембраны происходит выход ионов Са2, что обеспечивает слияние синаптических пузырьков с пресинаптической мембраной.
[4]
В большинстве аксонов кальциевые каналы имеются только в пресинаптической мембране, и даже здесь их сравнительно немного. По этой причине, а также потому, что внешние и внутриклеточные концентрации Са2 относительно низки, ток, протекающий через кальциевые каналы, обычно мал по сравнению с токами, проходящими через многочисленные потенциал-зависимые натриевые и калиевые каналы в окончании аксона.
[5]
Биосинтез ацетилхолина происходит в митохондриях нейронов, ацетилхолин скапливается в виде микровезикул вблизи пресинаптической мембраны. Возникновение нервного импульса сопровождается увеличением проницаемости и деполяризацией пресинаптических мембран. Благодаря этому микровезикулы с ацетилхолином притягиваются к мембрапе и высвобождают свое содержимое через мембрану в область синапса.
[6]
Такая инактивация происходит либо путем ферментативной деградации медиатора, либо путем его обратного поглощения пресинаптической мембраной.
[7]
Наиболее хорошо изучен холинергический синапс ( рис. 3) В нервном окончании передающей нервный импульс клетки находится так называемая пресинаптическая мембрана. Воспринимает этот нервный импульс мембрана другой клетки — постсинаптическаь мембрана. Между мембранами находится синаптическая щель шириной около 50 нм.
[9]
По своему строению и действию эфедрин близок важнейшему биологическому медиатору адренергическнх синапсов — норадреналину и гормонам надпочечников адреналину и дофамину; он слабо, но продолжительно стимулирует а — и р-адренорецепторы и действует на пресинаптические мембраны, высвобождая норадре-налин и являясь симпатомиметиком.
[10]
Нервные сигналы переходят от клетки к клетке через синапсы, которые могут быть электрическими ( щелевые контакты) или химическими. В химическом синапсе деполяризация пресинаптической мембраны в результате прибытия нервного импульса открывает потенциал-зависимые кальциевые каналы, вызывая тем самым приток Са2 в клетку, что приводит к освобождению нейромедиатора из синоптических пузырьков. Медиатор диффундирует в синоптическую щель и связывается с рецептурными белками в мембране постсинаптической клетки; в конечном итоге медиатор удаляется из синоптической щели путем диффузии, ферментативного расщепления или обратного поглощения выделившей его клеткой. Через рецепторные белки, образующие лиганд-зависимые каналы, реализуется быстрый постсинаптический эффект нейромедиатора-открытие каналов приводит к возникновению возбуждающего или тормозного постсинаптического потенциала в соответствии с ионной специфичностью каналов. При участии рецепторов, сопряженных с ферментами, например с аденилатциклазой, обычно осуществляются медленные и более продолжительные эффекты.
[11]
Свободный ацетилхолин в неактивной форме, связанный с белками, накапливается в окончании нервного волокна в специальных пузырьках — везикулах. В момент достижения нервным импульсом пресинаптической мембраны везикулы лопаются и ацетилхолин изливается в синаптическую щель. Достигая постсинаптической мембраны, он воздействует на холинорецепторы и вызывает соответствующую реакцию, например сокращение в случае мышечного волокна или нервный импульс в случае нервной клетки. Затем ацетилхолин быстро разрушается ферментом белковой природы — ацетилхолинэстеразой, находящейся на внешней поверхности постсинаптической мембраны, на малоактивные холин и уксусную кислоту. Расход ацетилхолина постоянно пополняется его синтезом — ацетилированием холина.
[13]
Биосинтез ацетилхолина происходит в митохондриях нейронов, ацетилхолин скапливается в виде микровезикул вблизи пресинаптической мембраны. Возникновение нервного импульса сопровождается увеличением проницаемости и деполяризацией пресинаптических мембран. Благодаря этому микровезикулы с ацетилхолином притягиваются к мембрапе и высвобождают свое содержимое через мембрану в область синапса.
[14]
Страницы:
1
2
3