Для более глубокого понимания механизма действия БТА, которое будет подробно описано ниже, необходимо вспомнить строение нервно-мышечного синапса.
Синапс (греч. synapsis – соединение) – это специализированная структура, обеспечивающая передачу возбуждающих или тормозящих влияний между двумя возбудимыми клетками.
Синапсы разделяют по виду соединяемых клеток, по эффекту, по природе медиатора (рис. 2).
Нервно-мышечный синапс образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры. Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и синаптическая мембрана, являющаяся частью плазматической мембраны постсинаптической мышечной клетки. Пресинаптической мембраной называют часть непокрытой миелином мембраны аксонной терминали, обращенной в синаптическую щель.
Пресинаптическая терминаль формируется тонкой веточкой аксона, которая, подходя к мышечному волокну, образует утолщение. В пресинаптической терминали находятся синаптические пузырьки (везикулы), содержащие запасы медиатора ацетилхолина. Везикул может быть до нескольких тысяч. Их диаметр около 40 нм, и в каждой содержится несколько тысяч молекул медиатора. Непосредственно около пресинаптической мембраны находится около 1000 везикул – оперативный запас, готовый к выделению в пресинаптическую щель. Остальная часть является мобильным запасом, готовым к быстрому перемещению к мембране взамен освободившегося медиатора. Наиболее существенным является стационарный резерв (около 300 тыс. везикул). Он находится в проксимальных отделах терминали. Освобождение медиатора в синаптическую щель происходит порциями. Минимальная порция – везикула медиатора, которая содержит около 5–10 тыс. молекул ацетилхолина и называется квантом (рис. 3).
В естественных
условиях (в нашем организме) возбуждение
мышечного волокна (или нескольких
мышечных волокон, составляющих мышцу)
возникает в результате передачи
возбуждения с нервного волокна на
мембрану мышечного в местах контакта
нерва и мышцы: нервно-мышечных синапсах.
Механизм
нервно-мышечной передачи
Синапсы представляют
собой коммуникационные структуры,
которые формируются окончанием нервного
волокна и прилегающей к нему мембраной
мышечного волокна (пресинаптической
нервной и постсинаптической мышечной
мембранами) (рис.2)
Когда нервный
импульс достигает окончания аксона, на
деполяризованной пресинаптической
мембране открываются потенциалзависимые
Са2+ каналы. Вход Са2+ в аксональное
расширение (пресинаптическую мембрану)
способствует высвобождению химических
нейромедиаторов, находящихся в виде
везикул (пузырьков) из окончания аксона.
Медиаторы (в нервно-мышечном синапсе
это всегда ацетилхолин) синтезируются
в соме нервной клетки и путем аксонального
транспорта транспортируются к окончанию
аксона, где и выполняют свою роль.
Медиатор диффундирует через синаптическую
щель и связывается со специфическими
рецепторами на постсинаптической
мембране. Так как медиатором в
нервно-мышечном синапсе является
ацетилхолин, то рецепторы постсинаптической
мембраны называют холинорецепторами.
В результате этого процесса на
постсинаптической мембране открываются
хемочувствительные Nа+-каналы, возникает
деполяризация, величина которой различна,
и зависит от количества выделенного
медиатора. Чаще всего возникает локальный
процесс, который называют потенциалом
концевой пластинки (ПКП). При повышении
частоты стимуляции нервного волокна,
усиливается деполяризация пресинаптической
мембраны, а, следовательно, возрастает
количество выделяемого медиатора и
число активированных хемочувствительных
Nа+каналов на постсинаптической мембране.
Таким образом, возникают ПКП, которые
по амплитуде деполяризации суммируются
до порогового уровня, после чего, на
мембране мышечного волокна, окружающей
синапс, возникает ПД, который обладает
способностью к распространению вдоль
мембраны мышечного волокна. Чувствительность
постсинаптической мембраны регулируется
активностью фермента – ацетилхолинэстеразы
(АЦХ-Э), который гидролизует медиатор
АЦХ на составные компоненты (ацетил и
холин) и возвращает назад – в
пресинаптическую бляшку для ресинтеза.
Без удаления медиатора на постсинаптической
мембране развивается длительная
деполяризация, которая ведет к нарушению
проведения возбуждения в синапсе –
синаптической депрессии. Таким образом,
синаптическая связь обеспечивает
одностороннее проведение возбуждения
с нерва на мышцу, однако на все эти
процессы расходуется время (синаптичекая
задержка), что приводит к низкой
лабильности синапса по сравнению с
нервным волокном.
Основными этапами
передачи возбуждения в нервно-мышечном
синапсе являются:
1) возбуждение
мотонейрона, распространение потенциала
действия на пресинаптическую мембрану;
2) повышение проницаемости
пресинаптической мембраны для ионов
кальция, ток кальция в клетку, повышение
концентрации кальция в пресинаптическом
окончаниии;
3) слияние синаптических
пузырьков с пресинаптической мембраной
в активной зоне, экзоцитоз, поступление
медиатора в синаптическую щель;
4) диффузия ацетилхолина
к постсинаптической мембране, присоединение
его к Н-холинорецепторам, открытие
хемозависимых ионных каналов;
5) преобладающий
ионный ток натрия через хемозависимые
каналы, образование надпорогового
потенциала концевой пластинки;
6) возникновение
потенциалов действия на мышечной
мембране;
7) ферментативное
расщепление ацетилхолина, возвращение
продуктов расщепления в окончание
нейрона, синтез новых порций медиатора.
29. Режимы одиночного
и тетанического сокращения мышечного
волокна. Формы мышечного сокращения —
динамическая и статическая. Концентрический
и эксцентрический типы мышечного
сокращения.
Одиночное
сокращение
Режим
сокращений мышечных волокон
определяется частотой импульсации
мотонейронов. Механический
ответ мышечного волокна или отдельной
мышцы на однократное их раздражение
называется одиночным
сокращением.
При одиночном
сокращении выделяют:
1. Фазу развития
напряжения или укорочения;
2. Фазу расслабления
или удлинения.
Фаза расслабления
продолжается примерно в два раза дольше,
чем фаза напряжения. Длительность этих
фаз зависит от морфофункциональных
свойств мышечного волокна: у наиболее
быстро сокращающихся волокон глазных
мышц фаза напряжения составляет 7-10 мс,
а у наиболее медленных волокон
камбаловидной мышцы — 50-100
мс.
В естественных
условиях мышечные волокна двигательной
единицы и скелетная мышца в целом
работают в режиме одиночного сокращения
только в том случае, когда длительность
интервала между последовательными
импульсами мотонейрона равна или
превышает длительность одиночного
сокращения иннервируемых им мышечных
волокон. Так, режим одиночного сокращения
медленных волокон камбаловидной мышцы
человека обеспечивается при частоте
импульсации мотонейрона менее 10 имп/с,
а быстрых волокон глазодвигательных
мышц — при частоте импульсации мотонейрона
менее 50 имп/с.
В
режиме одиночного сокращения мышца
способна работать длительное время
без развития утомления. Однако в связи
с тем, что длительность одиночного
сокращения невелика, развиваемое
мышечными волокнами напряжение не
достигает максимально возможных
величин. При относительно высокой
частоте импульсации мотонейронов
каждый последующий раздражающий импульс
приходится на фазу предшествующего
напряжения волокона, то есть до того
момента, когда оно начинает расслабляться.
В этом случае механические эффекты
каждого предыдущего сокращения
суммируются с последующим. Причем
величина механического ответа на каждый
последующий импульс меньше, чем на
предыдущий. После
нескольких первых импульсов последующие
ответы мышечных волокон не изменяют
достигнутого напряжения, а лишь
поддерживают его. Такой режим
сокращения называется гладким
тетанусом (рис.4.6.).
В подобном режиме двигательные единицы
мышц человека работают при развитии
максимальных изометрических усилий.
При гладком тетанусе развиваемое ДЕ
напряжение в 2-4 раза больше, чем при
одиночных сокращениях.
В тех
случаях, когда промежутки между
последовательными импульсами
мотонейрона меньше времени полного
цикла одиночного сокращения, но больше
длительности фазы напряжения, сила
сокращения ДЕ колеблется. Этот режим
сокращения называется зубчатым
тетанусом (рис.
4.6.).
Гладкий тетанус
для быстрых и медленных мыши достигается
при разных частотах импульсации
мотонейронов. Зависит это от времени
одиночного сокращения. Так, гладкий
тетанус для быстрой глазодвигательной
мышцы проявляется при частотах свыше
150-200 имп/с, а у медленной камбаловидной
мышцы — при частоте около 30 имп/с. В
режиме тетанического сокращения мышца
способна работать лишь короткое время.
Это объясняется тем, что из-за отсутствия
периода расслабления она не может
восстановить свой энергетический
потенциал и работает как бы «в
долг».
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
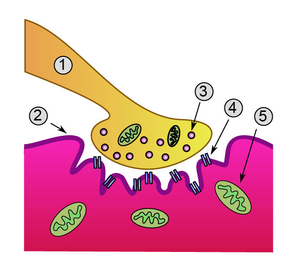
Нервно-мышечный синапс (также нейромышечный, либо мионевральный синапс) — эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в этом синапсе является ацетилхолин.
В этом синапсе нервный импульс превращается в механическое движение мышечной ткани.
Строение
Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с плазматической мембраной мышечного волокна, образующийся из выпячиваний аксона и цитолеммы мышечного волокна, создавая глубокие «карманы». Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором ацетилхолином.
![]()
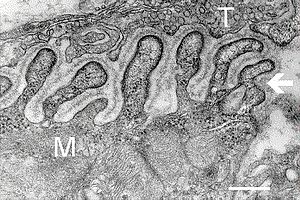
Электронная микрофотография среза нервномышечного синапса. Т — окончание аксона, М — мышечное волокно. Стрелка указывает на складки базальной мембраны. Шкала 0.3 мкм[1]
Морфология синапса в гладкомышечной ткани
Двигательные нервные окончания в гладкой мышечной ткани построены проще — безмиелиновые пучки аксонов проникают между глиоцитами к пласту гладких мышц и образуют булавовидные расширения, которые содержат холинергические и адренергические пузырьки.
Примечания
- ↑ Источник — NIMH
| |
|
|---|---|
| Нейроны (Серое вещество) |
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты) Дендрит (Вещество Ниссля, Дендритный шипик, Апикальный дендрит, Базальный дендрит) типы: Биполярные нейроны · Псевдополярные нейроны · Мультиполярные нейроны · Пирамидальный нейрон · Клетка Пуркинье · Гранулярная клетка |
| Афферентный нерв/ Сенсорный нерв/ Сенсорный нейрон |
GSA · GVA · SSA · SVA · Нервные волокна (Мышечные веретёна (Ia), Нервно-сухожильное веретено, II or Aβ, Aδ-волокна, C-волокна) |
| Эфферентный нерв/ Моторный нерв/ Моторный нейрон |
GSE · GVE · SVE · Верхний моторный нейрон · Нижний моторный нейрон (α мотонейроны, γ мотонейроны) |
| Синапс | Нейропиль · Синаптический пузырек · Нервно-мышечный синапс · Электрический синапс · Химический синапс · Интернейрон (Клетки Реншоу) |
| Сенсорный рецептор | Чувствительное тельце Мейснера · Нервное окончание Меркеля · Тельца Пачини · Окончание Руффини · Нервномышечное веретено · Свободное нервное окончание · Обонятельный нейрон · Фоторецепторные клетки · Волосковые клетки · Вкусовая луковица |
| Нейроглия | Астроциты (Радиальная глия) · Олигодендроглиоциты · Клетки эпендимы (Танициты) · Микроглия |
| Миелин (Белое вещество) |
CNS: Олигодендроцит PNS: Клетки Шванна · Невролемма · Перехват Ранвье/Межузловой сегмент · Насечка миелина |
| Соединительная ткань | Эпиневрий · Периневрий · Эндоневрий · Нервные пучки · Мозговые оболочки: твёрдая, паутинная, мягкая |