Как работает наш мозг или как смоделировать душу?
Время на прочтение
11 мин
Количество просмотров 50K

Здравствуй, Geektimes! В ранее опубликованной статье, была представлена модель нервной системы, опишу теорию и принципы, которые легли в её основу.
Теория основана на анализе имеющейся информации о биологическом нейроне и нервной системе из современной нейробиологии и физиологии мозга.
Сначала приведу краткую информацию об объекте моделирования, вся информация изложена далее, учтена и использована в модели.
НЕЙРОН

Нейрон является основным функциональным элементом нервной системы, он состоит из тела нервной клетки и её отростков. Существуют два вида отростков: аксоны и дендриты. Аксон – длинный покрытый миелиновой оболочкой отросток, предназначенный для передачи нервного импульса на далекие расстояния. Дендрит – короткий, ветвящийся отросток, благодаря которым происходит взаимосвязь с множеством соседних клеток.
ТРИ ТИПА НЕЙРОНОВ
Нейроны могут сильно отличаться по форме, размерам и конфигурации, не смотря на это, отмечается принципиальное сходство нервной ткани в различных участках нервной системе, отсутствуют и серьезные эволюционные различия. Нервная клетка моллюска Аплизии может выделять такие же нейромедиаторы и белки, что и клетка человека.
В зависимости от конфигурации выделяют три типа нейронов:

а) рецепторные, центростремительные, или афферентные нейроны, данные нейроны имеют центростремительный аксон, на конце которого имеются рецепторы, рецепторные или афферентные окончания. Эти нейроны можно определить, как элементы, передающие внешние сигналы в систему.
б) интернейроны (вставочные, контактные, или промежуточные) нейроны, не имеющие длинных отростков, но имеющие только дендриты. Таких нейронов в человеческом мозгу больше чем остальных. Данный вид нейронов является основным элементом рефлекторной дуги.
в) моторные, центробежные, или эфферентные, они имеют центростремительный аксон, который имеет эфферентные окончания передающий возбуждение мышечным или железистым клеткам. Эфферентные нейроны служат для передачи сигналов из нервной среды во внешнюю среду.
Обычно в статьях по искусственным нейронным сетям оговаривается наличие только моторных нейронов (с центробежным аксоном), которые связаны в слои иерархической структуры. Подобное описание применимо к биологической нервной системе, но является своего рода частным случаем, речь идет о структурах, базовых условных рефлексов. Чем выше в эволюционном значении нервная система, тем меньше в ней превалируют структуры типа «слои» или строгая иерархия.
ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ
Передача возбуждения происходит от нейрона к нейрону, через специальные утолщения на концах дендритов, называемых синапсами. По типу передачи синапсы разделяют на два вида: химические и электрические. Электрические синапсы передают нервный импульс непосредственно через место контакта. Таких синапсов в нервных системах очень мало, в моделях не будут учитываться. Химические синапсы передают нервный импульс посредством специального вещества медиатора (нейромедиатора, нейротрансмиттера), данный вид синапса широко распространен и подразумевает вариативность в работе.
Важно отметить, что в биологическом нейроне постоянно происходят изменения, отращиваются новые дендриты и синапсы, возможны миграции нейронов. В местах контактов с другими нейронами образуются новообразования, для передающего нейрона — это синапс, для принимающего — это постсинаптическая мембрана, снабжаемая специальными рецепторами, реагирующими на медиатор, то есть можно говорить, что мембрана нейрона — это приемник, а синапсы на дендритах — это передатчики сигнала.
СИНАПС

При активации синапса он выбрасывает порции медиатора, эти порции могут варьироваться, чем больше выделится медиатора, тем вероятнее, что принимаемая сигнал нервная клетка будет активирована. Медиатор, преодолевая синоптическую щель, попадает на постсинаптическую мембрану, на которой расположены рецепторы, реагирующие на медиатор. Далее медиатор может быть разрушен специальным разрушающим ферментом, либо поглощен обратно синапсом, это происходит для сокращения времени действия медиатора на рецепторы.
Так же помимо побудительного воздействия существуют синапсы, оказывающие тормозящее воздействие на нейрон. Обычно такие синапсы принадлежат определенным нейронам, которые обозначаются, как тормозящие нейроны.
Синапсов связывающих нейрон с одной и той же целевой клеткой, может быть множество. Для упрощения примем, всю совокупность, оказываемого воздействия одним нейроном, на другой целевой нейрон за синапс с определённой силой воздействия. Главной характеристикой синапса будет, является его сила.
СОСТОЯНИЕ ВОЗБУЖДЕНИЯ НЕЙРОНА
В состоянии покоя мембрана нейрона поляризована. Это означает, что по обе стороны мембраны располагаются частицы, несущие противоположные заряды. В состоянии покоя наружная поверхность мембраны заряжена положительно, внутренняя – отрицательно. Основными переносчиками зарядов в организме являются ионы натрия (Na+), калия (K+) и хлора (Cl-).
Разница между зарядами поверхности мембраны и внутри тела клетки составляет мембранный потенциал. Медиатор вызывает нарушения поляризации – деполяризацию. Положительные ионы снаружи мембраны устремляются через открытые каналы в тело клетки, меняя соотношение зарядов между поверхностью мембраны и телом клетки.

Изменение мембранного потенциала при возбуждении нейрона
Характер изменений мембранного потенциала при активации нервной ткани неизменен. Независимо от того кокой силы воздействия оказывается на нейрон, если сила превышает некоторое пороговое значение, ответ будет одинаков.
Забегая вперед, хочу отметить, что в работе нервной системы имеет значение даже следовые потенциалы (см. график выше). Они не появляются, вследствие каких-то гармонических колебаний уравновешивающих заряды, являются строгим проявлением определённой фазы состояния нервной ткани при возбуждении.
ТЕОРИЯ ЭЛЕКТРОМАГНИТНОГО ВЗАИМОДЕЙСТВИЯ
Итак, далее приведу теоретические предположения, которые позволят нам создавать математические модели. Главная идея заключается во взаимодействии между зарядами формирующихся внутри тела клетки, во время её активности, и зарядами с поверхностей мембран других активных клеток. Данные заряды являются разноименными, в связи этим можно предположить, как будут располагаться заряды в теле клетки под воздействием зарядов других активных клеток.

Можно сказать, что нейрон чувствует активность других нейронов на расстоянии, стремится направить распространения возбуждения в направлении других активных участков.
В момент активности нейрона можно рассчитать определённую точку в пространстве, которая определялась бы, как сумма масс зарядов, расположенных на поверхностях других нейронов. Указанную точку назовем точкой паттерна, её месторождение зависит от комбинации фаз активности всех нейронов нервной системы. Паттерном в физиологии нервной системы называется уникальная комбинация активных клеток, то есть можно говорить о влиянии возбуждённых участков мозга на работу отдельного нейрона.
Нужно представлять работу нейрона не просто как вычислителя, а своего рода ретранслятор возбуждения, который выбирает направления распространения возбуждения, таким образом, формируются сложные электрические схемы. Первоначально предполагалось, что нейрон просто избирательно отключает/включает для передачи свои синапсы, в зависимости от предпочитаемого направления возбуждения. Но более детальное изучение природы нейрона, привело к выводам, что нейрон может изменять степень воздействия на целевую клетку через силу своих синапсов, что делает нейрон более гибким и вариативным вычислительным элементом нервной системы.

Какое же направление для передачи возбуждения является предпочтительным? В различных экспериментах связанных с образованием безусловных рефлексов, можно определить, что в нервной системе образуются пути или рефлекторные дуги, которые связывают активируемые участки мозга при формировании безусловных рефлексов, создаются ассоциативные связи. Значит, нейрон должен передавать возбуждения к другим активным участкам мозга, запоминать направление и использовать его в дальнейшем.
Представим вектор начало, которого находится в центре активной клети, а конец направлен в точку паттерна определённую для данного нейрона. Обозначим, как вектор предпочитаемого направления распространения возбуждения (T, trend). В биологическом нейроне вектор Т может проявляться в структуре самой нейроплазмы, возможно, это каналы для движения ионов в теле клетки, или другие изменения в структуре нейрона.
Нейрон обладает свойством памяти, он может запоминать вектор Т, направление этого вектора, может меняться и перезаписываться в зависимости от внешних факторов. Степень с которой вектор Т может подвергается изменениям, называется нейропластичность.
Этот вектор в свою очередь оказывает влияние на работу синапсов нейрона. Для каждого синапса определим вектор S начало, которого находится в центре клетки, а конец направлен в центр целевого нейрона, с которым связан синапс. Теперь степень влияния для каждого синапса можно определить следующим образом: чем меньше угол между вектором T и S, тем больше синапс будет, усиливается; чем меньше угол, тем сильнее синапс будет ослабевать и возможно может прекратить передачу возбуждения. Каждый синапс имеет независимое свойство памяти, он помнит значение своей силы. Указанные значения изменяются при каждой активизации нейрона, под влиянием вектора Т, они либо увеличиваются, либо уменьшаются на определённое значение.
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ

Входные сигналы (x1, x2,…xn) нейрона представляют собой вещественные числа, которые характеризуют силу синапсов нейронов, оказывающих воздействие на нейрон.
Положительное значение входа означает побудительное воздействие, оказываемое на нейрон, а отрицательное значение – тормозящее воздействие.
Для биологического нейрона не имеет значение, откуда поступил возбуждающий его сигнал, результат его активности будет идентичен. Нейрон будет активизирован, когда сумма воздействий на него будет превышать определённое пороговое значение. Поэтому, все сигналы проходят через сумматор (а), а поскольку нейроны и нервная система работают в реальном времени, следовательно, воздействие входов должно оцениваться в короткий промежуток времени, то есть воздействие синапса имеет временный характер.
Результат сумматора проходит пороговую функцию (б), если сумма превосходит пороговое значение, то это приводит к активности нейрона.
При активации нейрон сигнализирует о своей активности системе, передовая информацию о своём положении в пространстве нервной системы и заряде, изменяемом во времени (в).
Через определённое время, после активации нейрон передает возбуждение по всем имеющимся синапсам, предварительно производя пересчет их силы. Весь период активации нейрон перестает реагировать на внешние раздражители, то есть все воздействия синапсов других нейронов игнорируются. В период активации входит так же период восстановления нейрона.
Происходит корректировка вектора Т (г) с учётом значения точки паттерна Pp и уровнем нейропластичности. Далее происходит переоценка значений всех сил синапсов в нейроне(д).
Обратите внимание, что блоки (г) и (д) выполняются параллельно с блоком (в).
ЭФФЕКТ ВОЛНЫ
Если внимательно проанализировать предложенную модель, то можно увидеть, что источник возбуждения должен оказывать большее влияние на нейрон, чем другой удалённый, активный участок мозга. Следовательно возникает вопрос: почему же все равно происходит передача в направлении другого активного участка?
Данную проблему я смог определить, только создав компьютерную модель. Решение подсказал график изменения мембранного потенциала при активности нейрона.

Усиленная реполяризация нейрона, как говорилось ранее, имеет важное значение для нервной системы, благодаря ей создается эффект волны, стремление нервного возбуждения распространятся от источника возбуждения.
При работе с моделью я наблюдал два эффекта, ели пренебречь следовым потенциалом или сделать его недостаточно большим, то возбуждение не распространяется от источников, а в большей степени стремится к локализации. Если сделать следовой потенциал сильно большим, то возбуждение стремится «разбежаться» в разные стороны, не только от своего источника, но и от других.
КОГНИТИВНАЯ КАРТА
Используя теорию электромагнитного взаимодействия, можно объяснить многие явления и сложные процессы, протекающие в нервной системе. К примеру, одним из последних открытий, которое широко обсуждается в науках о мозге, является открытие когнитивных карт в гиппокампе.
Гиппокамп – это отдел мозга, которому отвечает за кратковременную память. Эксперименты на крысах выявили, что определённому месту в лабиринте соответствует своя локализованная группа клеток в гиппокампе, причем, не имеет значение, как животное попадает в это место, все равно будет активирован соответствующий этому месту участок нервной ткани. Естественно, животное должно помнить данный лабиринт, не стоит рассчитывать на топологическое соответствие пространства лабиринта и когнитивной карты.

Каждое место в лабиринте представляется в мозге, как совокупность раздражителей различного характера: запахи, цвет стен, возможные примечательные объекты, характерные звуки и т. д. Указанные раздражители отражаются на коре, различных представительствах органов чувств, в виде всплесков активности в определённых комбинациях. Мозг одновременно обрабатывает информацию в нескольких отделах, зачастую информационные каналы разделяются, одна и та же информация поступает в различные участки мозга.

Активация нейронов места в зависимости от положения в лабиринте (активность разных нейронов показана разным цветом). источник
Гиппокамп расположен в центре мозга, вся кара и её области удалены от него, на одинаковые расстояния. Если определить для каждой уникальной комбинации раздражителей точку масс зарядов поверхностей нейронов, то можно увидеть, что указанные точки будут различны, и будут находиться примерно в центре мозга. К этим точкам будет стремиться и распространятся возбуждение в гиппокампе, формируя устойчивые участки возбуждения. Более того, поочередная смена комбинаций раздражителей, будет приводить к смещению точки паттерна. Участки когнитивной карты будут ассоциативно связываться друг с другом последовательно, что приведет к тому, что животное, помещенное в начало знакомого ей лабиринта, может вспомнить весь последующий путь.
Заключение
У многих возникнет вопрос, где в данной работе предпосылки к элементу разумности или проявления высшей интеллектуальной деятельности?
Важно отметить, что феномен человеческого поведения, есть следствие функционирования биологической структуры. Следовательно, чтобы имитировать разумное поведение, необходимо хорошо понимать принципы и особенности функционирования биологических структур. К сожалению, в науке биологии пока не представлен четкий алгоритм: как работает нейрон, как понимает, куда необходимо отращивать свои дендриты, как настроить свои синапсы, что бы в нервной системе смог сформироваться простой условный рефлекс, на подобие тех, которые демонстрировал и описывал в своих работах академик И.П. Павлов.
С другой стороны в науке об искусственном интеллекте, в восходящем (биологическом) подходе, сложилось парадоксальная ситуация, а именно: когда используемые в исследованиях модели основаны на устаревших представлениях о биологическом нейроне, консерватизм, в основе которого берётся персептрон без переосмысления его основных принципов, без обращения к биологическому первоисточнику, придумывается все более хитроумные алгоритмы и структуры, не имеющих биологических корней.
Конечно, никто не уменьшает достоинств классических нейронных сетей, которые дали множество полезных программных продуктов, но игра с ними не является путем к созданию интеллектуально действующей системы.
Более того, не редки заявления, о том, что нейрон подобен мощной вычислительной машине, приписывают свойство квантовых компьютеров. Из-за этой сверхсложности, нервной системе приписывается невозможность её повторения, ведь это соизмеримо с желанием смоделировать человеческую душу. Однако, в реальности природа идет по пути простоты и элегантности своих решений, перемещение зарядов на мембране клетки может служить, как для передачи нервного возбуждения, так и для трансляции информации о том, где происходит данная передача.
Несмотря на то, что указанная работа демонстрирует, как образуются элементарные условные рефлексы в нервной системе, она приближает к пониманию того, что такое интеллект и разумная деятельность.
Существуют еще множество аспектов работы нервной системы: механизмы торможения, принципы построения эмоций, организация безусловных рефлексов и обучение, без которых невозможно построить качественную модель нервной системы. Есть понимание, на интуитивном уровне, как работает нервная система, принципы которой возможно воплотить в моделях.
Создание первой модели помогли отработать и откорректировать представление об электромагнитном взаимодействии нейронов. Понять, как происходит формирование рефлекторных дуг, как каждый отдельный нейрон понимает, каким образом ему настроить свои синапсы для получения ассоциативных связей.
На данный момент я начал разрабатывать новую версию программы, которая позволит смоделировать многие другие аспекты работы нейрона и нервной системы.
Прошу принять активное участие в обсуждении выдвинутых здесь гипотез и предположений, так как я могу относиться к своим идеям предвзято. Ваше мнение очень важно для меня.
Модель(Windows PC) + туториал
Статья на конкурс «био/мол/текст»: Клеточные процессы, обеспечивающие обмен информацией между нейронами, требуют много энергии. Высокое энергопотребление способствовало в ходе эволюции отбору наиболее эффективных механизмов кодирования и передачи информации. В этой статье вы узнаете о теоретическом подходе к изучению энергетики мозга, о его роли в исследованиях патологий, о том, какие нейроны более продвинуты, почему синапсам иногда выгодно не «срабатывать», а также, как они отбирают только нужную нейрону информацию.

Конкурс «био/мол/текст»-2017
Эта работа опубликована в номинации «Свободная тема» конкурса «био/мол/текст»-2017.

Генеральный спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.

Спонсором приза зрительских симпатий и партнером номинации «Биомедицина сегодня и завтра» выступила фирма «Инвитро».

«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Происхождение подхода
С середины ХХ века известно, что головной мозг потребляет значительную часть энергоресурсов всего организма: четверть всей глюкозы и ⅕ всего кислорода в случае высшего примата [1–5]. Это вдохновило Уильяма Леви и Роберта Бакстера из Массачусетского технологического института (США) на проведение теоретического анализа энергетической эффективности кодирования информации в биологических нейронных сетях (рис. 1) [6]. В основе исследования лежит следующая гипотеза. Поскольку энергопотребление мозга велико, ему выгодно иметь такие нейроны, которые работают наиболее эффективно — передают только полезную информацию и затрачивают при этом минимум энергии.
Это предположение оказалось справедливым: на простой модели нейронной сети авторы воспроизвели экспериментально измеренные значения некоторых параметров [6]. В частности, рассчитанная ими оптимальная частота генерации импульсов варьирует от 6 до 43 имп./с — почти так же, как и у нейронов основания гиппокампа. Их можно подразделить на две группы по частоте импульсации: медленные (~10 имп./с) и быстрые (~40 имп./с). При этом первая группа значительно превосходит по численности вторую [7]. Аналогичная картина наблюдается и в коре больших полушарий: медленных пирамидальных нейронов (~4—9 имп./с) в несколько раз больше, чем быстрых ингибиторных интернейронов (>100 имп./с) [8], [9]. Так, видимо, мозг «предпочитает» использовать поменьше быстрых и энергозатратных нейронов, чтобы те не израсходовали все ресурсы [6], [9–11].

Рисунок 1. Представлены два нейрона. В одном из них фиолетовым цветом окрашен пресинаптический белок синаптофизин. Другой нейрон полностью окрашен зеленым флуоресцентным белком. Мелкие светлые крапинки — синаптические контакты между нейронами [12]. Во вставке одна «крапинка» представлена ближе.
Группы нейронов, связанных между собой синапсами, называются нейронными сетями [13], [14]. Например, в коре больших полушарий пирамидальные нейроны и интернейроны образуют обширные сети. Слаженная «концертная» работа этих клеток обусловливает наши высшие когнитивные и другие способности. Аналогичные сети, только из других типов нейронов, распределены по всему мозгу, определенным образом связаны между собой и организуют работу всего органа.
Работа Леви и Бакстера [6] развивает концепцию «экономии импульсов» Горация Барлоу из Университета Калифорнии (США), который, кстати, является потомком Чарльза Дарвина [17]. Согласно ей, при развитии организма нейроны стремятся работать только с наиболее полезной информацией, фильтруя «лишние» импульсы, ненужную и избыточную информацию. Однако эта концепция не дает удовлетворительных результатов, так как не учитывает метаболические затраты, связанные с нейрональной активностью [6]. Расширенный подход Леви и Бакстера, в котором внимание уделено обоим факторам, оказался более плодотворным [6], [18–20]. И энергозатраты нейронов, и потребность в кодировании только полезной информации являются важными факторами, направляющими эволюцию мозга [6], [21–24]. Поэтому, чтобы лучше разобраться в том, как устроен мозг, стоит рассматривать обе эти характеристики: сколько нейрон передает полезной информации и сколько энергии при этом тратит.
За последнее время этот подход нашел множество подтверждений [10], [22], [24–26]. Он позволил по-новому взглянуть на устройство мозга на самых разных уровнях организации — от молекулярно-биофизического [20], [26] до органного [23]. Он помогает понять, каковы компромиссы между выполняемой функцией нейрона и ее энергетической ценой и в какой степени они выражены.
Как же работает этот подход?
Положим, у нас есть модель нейрона, описывающая его электрофизиологические свойства: потенциал действия (ПД) и постсинаптические потенциалы (ПСП) (об этих терминах — ниже). Мы хотим понять, эффективно ли он работает, не тратит ли неоправданно много энергии. Для этого нужно вычислить значения параметров модели (например, плотность каналов в мембране, скорость их открывания и закрывания), при которых: (а) достигается максимум отношения полезной информации к энергозатратам и в то же время (б) сохраняются реалистичные характеристики передаваемых сигналов [6], [19].
Эти «оптимальные» значения параметров затем нужно сравнить с измеренными экспериментально и определить, насколько они отличаются. Общая картина отличий укажет на степень оптимизации данного нейрона в целом: насколько реальные, измеренные экспериментально, значения параметров совпадают с рассчитанными. Чем слабее выражены отличия, тем нейрон более близок к оптимуму и работает энергетически более эффективно, оптимально. С другой стороны, сопоставление конкретных параметров покажет, в каком конкретно качестве этот нейрон близок к «идеалу».
Далее, в контексте энергетической эффективности нейронов рассмотрены два процесса, на которых основано кодирование и передача информации в мозге. Это нервный импульс, или потенциал действия, благодаря которому информация может быть отправлена «адресату» на определенное расстояние (от микрометров до полутора метров) и синаптическая передача, лежащая в основе собственно передачи сигнала от одного нейрона на другой.
Потенциал действия
Потенциал действия (ПД) — сигнал, которые отправляют друг другу нейроны. ПД бывают разные: быстрые и медленные, малые и большие [28]. Зачастую они организованы в длинные последовательности (как буквы в слова), либо в короткие высокочастотные «пачки» (рис. 2).

Рисунок 2. Разные типы нейронов генерируют различные сигналы. В центре — продольный срез мозга млекопитающего. Во вставках представлены разные типы сигналов, зарегистрированные методами электрофизиологии [15], [38]. а — Кортикальные (Cerebral cortex) пирамидальные нейроны могут передавать как низкочастотные сигналы (Regular firing), так и короткие взрывные, или пачечные, сигналы (Burst firing). б — Для клеток Пуркинье мозжечка (Cerebellum) характерна только пачечная активность на очень высокой частоте. в — Релейные нейроны таламуса (Thalamus) имеют два режима активности: пачечный и тонический (Tonic firing). г — Нейроны средней части поводка (MHb, Medial habenula) эпиталамуса генерируют тонические сигналы низкой частоты.
[14], рисунок адаптирован
Большое разнообразие сигналов обусловлено огромным количеством комбинаций разных типов ионных каналов, синаптических контактов, а также морфологией нейронов [28], [29]. Поскольку в основе сигнальных процессов нейрона лежат ионные токи, стоит ожидать, что разные ПД требуют различных энергозатрат [20], [27], [30].
Анализ разных типов нейронов (рис. 4) показал, что нейроны беспозвоночных не очень энергоэффективны, а некоторые нейроны позвоночных почти совершенны [20]. По результатам этого исследования, наиболее энергоэффективными оказались интернейроны гиппокампа, участвующего в формировании памяти и эмоций, а также таламокортикальные релейные нейроны, несущие основной поток сенсорной информации от таламуса к коре больших полушарий.

Рисунок 4. Разные нейроны эффективны по-разному. На рисунке представлено сравнение энергозатрат разных типов нейронов. Энергозатраты рассчитаны в моделях как с исходными (реальными) значениями параметров (черные столбцы), так и с оптимальными, при которых с одной стороны нейрон выполняет положенную ему функцию, с другой — затрачивает при этом минимум энергии (серые столбцы). Самыми эффективными из представленных оказались два типа нейронов позвоночных: интернейроны гиппокампа (rat hippocampal interneuron, RHI) и таламокортикальные нейроны (mouse thalamocortical relay cell, MTCR), так как для них энергозатраты в исходной модели наиболее близки к энергозатратам оптимизированной. Напротив, нейроны беспозвоночных менее эффективны. Условные обозначения: SA (squid axon) — гигантский аксон кальмара; CA (crab axon) — аксон краба; MFS (mouse fast spiking cortical interneuron) — быстрый кортикальный интернейрон мыши; BK (honeybee mushroom body Kenyon cell) — грибовидная клетка Кеньона пчелы.
[20], рисунок адаптирован
Почему они более эффективны? Потому что у них малó перекрывание Na- и К-токов. Во время генерации ПД всегда есть промежуток времени, когда эти токи присутствуют одновременно (рис. 3в). При этом переноса заряда практически не происходит, и изменение мембранного потенциала минимально. Но «платить» за эти токи в любом случае приходится, несмотря на их «бесполезность» в этот период. Поэтому его продолжительность определяет, сколько энергетических ресурсов растрачивается впустую. Чем он короче, тем более эффективно использование энергии [20], [26], [30], [43]. Чем длиннее — тем менее эффективно. Как раз в двух вышеупомянутых типах нейронов, благодаря быстрым ионным каналам, этот период очень короткий, а ПД — самые эффективные [20].
Кстати, интернейроны гораздо более активны, чем большинство других нейронов мозга. В то же время они крайне важны для слаженной, синхронной работы нейронов, с которыми образуют небольшие локальные сети [9], [16]. Вероятно, высокая энергетическая эффективность ПД интернейронов является некой адаптацией к их высокой активности и роли в координации работы других нейронов [20].
Синапс
Передача сигнала от одного нейрона к другому происходит в специальном контакте между нейронами, в синапсе [12]. Мы рассмотрим только химические синапсы (есть еще электрические), поскольку они весьма распространены в нервной системе и важны для регуляции клеточного метаболизма, доставки питательных веществ [5].
Чаще всего, химический синапс образован между окончанием аксона одного нейрона и дендритом другого. Его работа напоминает… «переброс» эстафетной палочки, роль которой и играет нейромедиатор — химический посредник передачи сигнала [12], [42], [44–48].
На пресинаптическом окончании аксона ПД вызывает выброс нейромедиатора во внеклеточную среду — к принимающему нейрону. Последний только этого и ждет с нетерпением: в мембране дендритов рецепторы — ионные каналы определенного типа — связывают нейромедиатор, открываются и пропускают через себя разные ионы. Это приводит к генерации маленького постсинаптического потенциала (ПСП) на мембране дендрита. Он напоминает ПД, но значительно меньше по амплитуде и происходит за счет открывания других каналов. Множество этих маленьких ПСП, каждый от своего синапса, «сбегаются» по мембране дендритов к телу нейрона (зеленые стрелки на рис. 3а) и достигают начального сегмента аксона, где вызывают открывание Na-каналов и «провоцируют» его на генерацию ПД.
Такие синапсы называются возбуждающими: они способствуют активации нейрона и генерации ПД. Существуют также и тормозящие синапсы. Они, наоборот, способствуют торможению и препятствуют генерации ПД. Часто на одном нейроне есть и те, и другие синапсы. Определенное соотношение между торможением и возбуждением важно для нормальной работы мозга, формирования мозговых ритмов, сопровождающих высшие когнитивные функции [49].
Как это ни странно, выброс нейромедиатора в синапсе может и не произойти вовсе — это процесс вероятностный [18], [19]. Нейроны так экономят энергию: синаптическая передача и так обусловливает около половины всех энергозатрат нейронов [25]. Если бы синапсы всегда срабатывали, вся энергия пошла бы на обеспечение их работы, и не осталось бы ресурсов для других процессов. Более того, именно низкая вероятность (20–40%) выброса нейромедиатора соответствует наибольшей энергетической эффективности синапсов. Отношение количества полезной информации к затрачиваемой энергии в этом случае максимально [18], [19]. Так, выходит, что «неудачи» играют важную роль в работе синапсов и, соответственно, всего мозга. А за передачу сигнала при иногда «не срабатывающих» синапсах можно не беспокоиться, так как между нейронами обычно много синапсов, и хоть один из них да сработает.
Еще одна особенность синаптической передачи состоит в разделении общего потока информации на отдельные компоненты по частоте модуляции приходящего сигнала (грубо говоря, частоте приходящих ПД) [50]. Это происходит благодаря комбинированию разных рецепторов на постсинаптической мембране [38], [50]. Некоторые рецепторы активируются очень быстро: например, AMPA-рецепторы (AMPA происходит от α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid). Если на постсинаптическом нейроне представлены только такие рецепторы, он может четко воспринимать высокочастотный сигнал (такой, как, например, на рис. 2в). Ярчайший пример — нейроны слуховой системы, участвующие в определении местоположения источника звука и точном распознавании коротких звуков типа щелчка, широко представленных в речи [12], [38], [51]. NMDA-рецепторы (NMDA — от N—methyl-D—aspartate) более медлительны. Они позволяют нейронам отбирать сигналы более низкой частоты (рис. 2г), а также воспринимать высокочастотную серию ПД как нечто единое — так называемое интегрирование синаптических сигналов [14]. Есть еще более медленные метаботропные рецепторы, которые при связывании нейромедиатора, передают сигнал на цепочку внутриклеточных «вторичных посредников» для подстройки самых разных клеточных процессов. К примеру, широко распространены рецепторы, ассоциированные с G-белками. В зависимости от типа они, например, регулируют количество каналов в мембране или напрямую модулируют их работу [14].
Различные комбинации быстрых AMPA-, более медленных NMDA- и метаботропных рецепторов позволяют нейронам отбирать и использовать наиболее полезную для них информацию, важную для их функционирования [50]. А «бесполезная» информация отсеивается, она не «воспринимается» нейроном. В таком случае не приходится тратить энергию на обработку ненужной информации. В этом и состоит еще одна сторона оптимизации синаптической передачи между нейронами.
Что еще?
Энергетическая эффективность клеток мозга исследуется также и в отношении их морфологии [35], [52–54]. Исследования показывают, что ветвление дендритов и аксона не хаотично и тоже экономит энергию [52], [54]. Например, аксон ветвится так, чтобы суммарная длина пути, который проходит ПД, была наименьшей. В таком случае энергозатраты на проведение ПД вдоль аксона минимальны.
Снижение энергозатрат нейрона достигается также при определенном соотношении тормозящих и возбуждающих синапсов [55]. Это имеет прямое отношение, например, к ишемии (патологическому состоянию, вызванному нарушением кровотока в сосудах) головного мозга. При этой патологии, вероятнее всего, первыми выходят из строя наиболее метаболически активные нейроны [9], [16]. В коре они представлены ингибиторными интернейронами, образующими тормозящие синапсы на множестве других пирамидальных нейронов [9], [16], [49]. В результате гибели интернейронов, снижается торможение пирамидальных. Как следствие, возрастает общий уровень активности последних (чаще срабатывают активирующие синапсы, чаще генерируются ПД). За этим немедленно следует рост их энергопотребления, что в условиях ишемии может привести к гибели нейронов.
При изучении патологий внимание уделяют и синаптической передаче как наиболее энергозатратному процессу [19]. Например, при болезнях Паркинсона [56], Хантингтона [57], Альцгеймера [58–61] происходит нарушение работы или транспорта к синапсам митохондрий, играющих основную роль в синтезе АТФ [62], [63]. В случае болезни Паркинсона, это может быть связано с нарушением работы и гибелью высоко энергозатратных нейронов черной субстанции, важной для регуляции моторных функций, тонуса мышц. При болезни Хантингтона, мутантный белок хангтингтин нарушает механизмы доставки новых митохондрий к синапсам, что приводит к «энергетическому голоданию» последних, повышенной уязвимости нейронов и избыточной активации. Все это может вызвать дальнейшие нарушения работы нейронов с последующей атрофией полосатого тела и коры головного мозга. При болезни Альцгеймера нарушение работы митохондрий (параллельно со снижением количества синапсов) происходит из-за отложения амилоидных бляшек. Действие последних на митохондрии приводит к окислительному стрессу, а также к апоптозу — клеточной гибели нейронов.
Еще раз обо всем
В конце ХХ века зародился подход к изучению мозга, в котором одновременно рассматривают две важные характеристики: сколько нейрон (или нейронная сеть, или синапс) кодирует и передает полезной информации и сколько энергии при этом тратит [6], [18], [19]. Их соотношение является своего рода критерием энергетической эффективности нейронов, нейронных сетей и синапсов.
Использование этого критерия в вычислительной нейробиологии дало существенный прирост к знаниям относительно роли некоторых явлений, процессов [6], [18–20], [26], [30], [43], [55]. В частности, малая вероятность выброса нейромедиатора в синапсе [18], [19], определенный баланс между торможением и возбуждением нейрона [55], выделение только определенного рода приходящей информации благодаря определенной комбинации рецепторов [50] — все это способствует экономии ценных энергетических ресурсов.
Более того, само по себе определение энергозатрат сигнальных процессов (например, генерация, проведение ПД, синаптическая передача) позволяет выяснить, какой из них пострадает в первую очередь при патологическом нарушении доставки питательных веществ [10], [25], [56]. Так как больше всего энергии требуется для работы синапсов, именно они первыми выйдут из строя при таких патологиях, как ишемия, болезни Альцгеймера и Хантингтона [19], [25]. Схожим образом определение энергозатрат разных типов нейронов помогает выяснить, какой из них погибнет раньше других в случае патологии. Например, при той же ишемии, в первую очередь выйдут из строя интернейроны коры [9], [16]. Эти же нейроны из-за интенсивного метаболизма — наиболее уязвимые клетки и при старении, болезни Альцгеймера и шизофрении [16].
В общем, подход к определению энергетически эффективных механизмов работы мозга является мощным направлением для развития и фундаментальной нейронауки, и ее медицинских аспектов [5], [14], [16], [20], [26], [55], [64].
Благодарности
Искренне благодарен моим родителям Ольге Наталевич и Александру Жукову, сестрам Любе и Алене, моему научному руководителю Алексею Браже и замечательным друзьям по лаборатории Эвелине Никельшпарг и Ольге Слатинской за поддержку и вдохновение, ценные замечания, сделанные при прочтении статьи. Я также очень благодарен редактору статьи Анне Петренко и главреду «Биомолекулы» Антону Чугунову за пометки, предложения и замечания.
Литература
- Прожорливый мозг;
- SEYMOUR S. KETY. (1957). THE GENERAL METABOLISM OF THE BRAIN IN VIVO. Metabolism of the Nervous System. 221-237;
- L. Sokoloff, M. Reivich, C. Kennedy, M. H. Des Rosiers, C. S. Patlak, et. al.. (1977). THE [14C]DEOXYGLUCOSE METHOD FOR THE MEASUREMENT OF LOCAL CEREBRAL GLUCOSE UTILIZATION: THEORY, PROCEDURE, AND NORMAL VALUES IN THE CONSCIOUS AND ANESTHETIZED ALBINO RAT. J Neurochem. 28, 897-916;
- Magistretti P.J. (2008). Brain energy metabolism. In Fundamental neuroscience // Ed by. Squire L.R., Berg D., Bloom F.E., du Lac S., Ghosh A., Spitzer N. San Diego: Academic Press, 2008. P. 271–297;
- Pierre J. Magistretti, Igor Allaman. (2015). A Cellular Perspective on Brain Energy Metabolism and Functional Imaging. Neuron. 86, 883-901;
- William B Levy, Robert A. Baxter. (1996). Energy Efficient Neural Codes. Neural Computation. 8, 531-543;
- Sharp P.E. and Green C. (1994). Spatial correlates of firing patterns of single cells in the subiculum of the freely moving rat. J. Neurosci. 14, 2339–2356;
- H. Hu, J. Gan, P. Jonas. (2014). Fast-spiking, parvalbumin+ GABAergic interneurons: From cellular design to microcircuit function. Science. 345, 1255263-1255263;
- Oliver Kann, Ismini E Papageorgiou, Andreas Draguhn. (2014). Highly Energized Inhibitory Interneurons are a Central Element for Information Processing in Cortical Networks. J Cereb Blood Flow Metab. 34, 1270-1282;
- David Attwell, Simon B. Laughlin. (2001). An Energy Budget for Signaling in the Grey Matter of the Brain. J Cereb Blood Flow Metab. 21, 1133-1145;
- Henry Markram, Maria Toledo-Rodriguez, Yun Wang, Anirudh Gupta, Gilad Silberberg, Caizhi Wu. (2004). Interneurons of the neocortical inhibitory system. Nat Rev Neurosci. 5, 793-807;
- Как происходит выделение нейромедиатора;
- От живого мозга к искусственному интеллекту;
- Kandel E.R., Schwartz J.H., Jessell T.M., Siegelbaum S.A., Hudspeth A.J. Principles of neural science (5th Edition). NY: McGraw-Hill Education / Medical, 2012. — 1760 p.;
- 12 методов в картинках: нейробиология;
- Oliver Kann. (2016). The interneuron energy hypothesis: Implications for brain disease. Neurobiology of Disease. 90, 75-85;
- H. B. Barlow. (1969). Trigger Features, Adaptation and Economy of Impulses. Information Processing in The Nervous System. 209-230;
- Levy W.B. and Baxter R.A. (2002). Energy-efficient neuronal computation via quantal synaptic failures. J. Neurosci. 22, 4746–4755;
- Julia J. Harris, Renaud Jolivet, David Attwell. (2012). Synaptic Energy Use and Supply. Neuron. 75, 762-777;
- Biswa Sengupta, Martin Stemmler, Simon B. Laughlin, Jeremy E. Niven. (2010). Action Potential Energy Efficiency Varies Among Neuron Types in Vertebrates and Invertebrates. PLoS Comput Biol. 6, e1000840;
- Simon B. Laughlin, Rob R. de Ruyter van Steveninck, John C. Anderson. (1998). Unknown title.. Nat. Neurosci.. 1, 36-41;
- S Laughlin. (2001). Energy as a constraint on the coding and processing of sensory information. Current Opinion in Neurobiology. 11, 475-480;
- J. E. Niven, S. B. Laughlin. (2008). Energy limitation as a selective pressure on the evolution of sensory systems. Journal of Experimental Biology. 211, 1792-1804;
- Jeremy E Niven. (2016). Neuronal energy consumption: biophysics, efficiency and evolution. Current Opinion in Neurobiology. 41, 129-135;
- Clare Howarth, Padraig Gleeson, David Attwell. (2012). Updated Energy Budgets for Neural Computation in the Neocortex and Cerebellum. J Cereb Blood Flow Metab. 32, 1222-1232;
- A. Hasenstaub, S. Otte, E. Callaway, T. J. Sejnowski. (2010). Metabolic cost as a unifying principle governing neuronal biophysics. Proceedings of the National Academy of Sciences. 107, 12329-12334;
- Abdelmalik Moujahid, Alicia D’Anjou, Manuel Graña. (2014). Energy demands of diverse spiking cells from the neocortex, hippocampus, and thalamus. Front. Comput. Neurosci.. 8;
- Bruce P. Bean. (2007). The action potential in mammalian central neurons. Nat Rev Neurosci. 8, 451-465;
- Izhikevich E.M. Dynamical systems in neuroscience: the geometry of excitability and bursting (computational neuroscience). The MIT Press, 2007. — 457 p.;
- Brett C. Carter, Bruce P. Bean. (2009). Sodium Entry during Action Potentials of Mammalian Neurons: Incomplete Inactivation and Reduced Metabolic Efficiency in Fast-Spiking Neurons. Neuron. 64, 898-909;
- Формирование мембранного потенциала покоя;
- Появление и эволюция клеточной мембраны;
- Липидный фундамент жизни;
- Alan Woodruff, Rafael Yuste. (2008). Of Mice and Men, and Chandeliers. PLoS Biol. 6, e243;
- György Buzsáki, Caroline Geisler, Darrell A. Henze, Xiao-Jing Wang. (2004). Interneuron Diversity series: Circuit complexity and axon wiring economy of cortical interneurons. Trends in Neurosciences. 27, 186-193;
- David Sterratt, Bruce Graham, Andrew Gillies, David Willshaw. (Unknown published date.). Introduction. Principles of Computational Modelling in Neuroscience. 1-12;
- B. C. Carter, B. P. Bean. (2011). Incomplete Inactivation and Rapid Recovery of Voltage-Dependent Sodium Channels During High-Frequency Firing in Cerebellar Purkinje Neurons. Journal of Neurophysiology. 105, 860-871;
- Камкин А.Г. и Каменский А.А. (2004). Фундаментальная и клиническая физиология. М.: «Академия», 2004. — 1073 с.;
- Hille B. (2001). Ion channels of excitable membranes. Sunderland: Sinauer Associates, 2001. — 814 p.;
- Brian D. Clark, Ethan M. Goldberg, Bernardo Rudy. (2009). Electrogenic Tuning of the Axon Initial Segment. Neuroscientist. 15, 651-668;
- Джаксон М.Б. Молекулярная и клеточная биофизика. М.: «БИНОМ», 2013. — 552 с.;
- Спокоен как GABA;
- P. Crotty. (2006). Metabolic Energy Cost of Action Potential Velocity. Journal of Neurophysiology. 96, 1237-1246;
- Дофаминовые болезни;
- Серотониновые сети;
- Тайны голубого пятна;
- Молекула здравого ума;
- Очень нервное возбуждение;
- György Buzsáki, Kai Kaila, Marcus Raichle. (2007). Inhibition and Brain Work. Neuron. 56, 771-783;
- David Attwell, Alasdair Gibb. (2005). Neuroenergetics and the kinetic design of excitatory synapses. Nat Rev Neurosci. 6, 841-849;
- Nace L. Golding, Donata Oertel. (2012). Synaptic integration in dendrites: exceptional need for speed. The Journal of Physiology. 590, 5563-5569;
- Ashish Raj, Yu-hsien Chen. (2011). The Wiring Economy Principle: Connectivity Determines Anatomy in the Human Brain. PLoS ONE. 6, e14832;
- B. L. Chen, D. H. Hall, D. B. Chklovskii. (2006). Wiring optimization can relate neuronal structure and function. Proceedings of the National Academy of Sciences. 103, 4723-4728;
- Dmitri B. Chklovskii, Thomas Schikorski, Charles F. Stevens. (2002). Wiring Optimization in Cortical Circuits. Neuron. 34, 341-347;
- Biswa Sengupta, Simon B. Laughlin, Jeremy E. Niven. (2013). Balanced Excitatory and Inhibitory Synaptic Currents Promote Efficient Coding and Metabolic Efficiency. PLoS Comput Biol. 9, e1003263;
- Болезнь Паркинсона: что изучать? как изучать?;
- Как спасти Тринадцатую? (Перспективы лечения болезни Хантингтона);
- Болезнь Альцгеймера: ген, от которого я без ума;
- Альцгеймеровский нейротоксин: ядовиты не только фибриллы;
- Возможно, β-амилоид болезни Альцгеймера — часть врождённого иммунитета;
- Новый шаг к пониманию болезни Альцгеймера: возможно, недосыпание является одним из факторов риска;
- Болезни и изменения клеточного метаболизма;
- Сон и старение II: Чем отличается сон пожилых и больных от сна молодых и здоровых?;
- Adelbert Ames. (2000). CNS energy metabolism as related to function. Brain Research Reviews. 34, 42-68.
From Wikipedia, the free encyclopedia
This article is about synapses of the nervous system. For other uses, see Synapse (disambiguation).
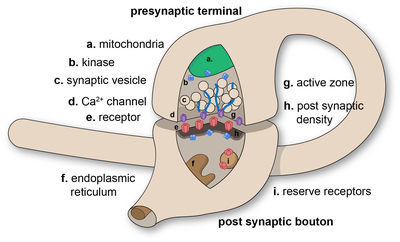
Diagram of a chemical synaptic connection.
In the nervous system, a synapse[1] is a structure that permits a neuron (or nerve cell) to pass an electrical or chemical signal to another neuron or to the target effector cell.
Synapses are essential to the transmission of nervous impulses from one neuron to another. Neurons are specialized to pass signals to individual target cells, and synapses are the means by which they do so. At a synapse, the plasma membrane of the signal-passing neuron (the presynaptic neuron) comes into close apposition with the membrane of the target (postsynaptic) cell. Both the presynaptic and postsynaptic sites contain extensive arrays of molecular machinery that link the two membranes together and carry out the signaling process. In many synapses, the presynaptic part is located on an axon and the postsynaptic part is located on a dendrite or soma. Astrocytes also exchange information with the synaptic neurons, responding to synaptic activity and, in turn, regulating neurotransmission.[2] Synapses (at least chemical synapses) are stabilized in position by synaptic adhesion molecules (SAMs) projecting from both the pre- and post-synaptic neuron and sticking together where they overlap; SAMs may also assist in the generation and functioning of synapses.[3]
History[edit]
Santiago Ramón y Cajal proposed that neurons are not continuous throughout the body, yet still communicate with each other, an idea known as the neuron doctrine.[4] The word «synapse» was introduced in 1897 by the English neurophysiologist Charles Sherrington in Michael Foster’s Textbook of Physiology.[1] Sherrington struggled to find a good term that emphasized a union between two separate elements, and the actual term «synapse» was suggested by the English classical scholar Arthur Woollgar Verrall, a friend of Foster.[5][6] The word was derived from the Greek synapsis (σύναψις), meaning «conjunction», which in turn derives from synaptein (συνάπτειν), from syn (σύν) «together» and haptein (ἅπτειν) «to fasten».[5][7]
However, while the synaptic gap remained a theoretical construct, and sometimes reported as a discontinuity between contiguous axonal terminations and dendrites or cell bodies, histological methods using the best light microscopes of the day could not visually resolve their separation which is now known to be about 20nm. It needed the electron microscope in the 1950s to show the finer structure of the synapse with its separate, parallel pre- and postsynaptic membranes and processes, and the cleft between the two.[8][9][10]
Chemical and electrical synapses[edit]

There are two fundamentally different types of synapses:
- In a chemical synapse, electrical activity in the presynaptic neuron is converted (via the activation of voltage-gated calcium channels) into the release of a chemical called a neurotransmitter that binds to receptors located in the plasma membrane of the postsynaptic cell. The neurotransmitter may initiate an electrical response or a secondary messenger pathway that may either excite or inhibit the postsynaptic neuron. Chemical synapses can be classified according to the neurotransmitter released: glutamatergic (often excitatory), GABAergic (often inhibitory), cholinergic (e.g. vertebrate neuromuscular junction), and adrenergic (releasing norepinephrine). Because of the complexity of receptor signal transduction, chemical synapses can have complex effects on the postsynaptic cell.
- In an electrical synapse, the presynaptic and postsynaptic cell membranes are connected by special channels called gap junctions that are capable of passing an electric current, causing voltage changes in the presynaptic cell to induce voltage changes in the postsynaptic cell. The main advantage of an electrical synapse is the rapid transfer of signals from one cell to the next.[11]
Synaptic communication is distinct from an ephaptic coupling, in which communication between neurons occurs via indirect electric fields.
An autapse is a chemical or electrical synapse that forms when the axon of one neuron synapses onto dendrites of the same neuron.
Types of interfaces[edit]
Synapses can be classified by the type of cellular structures serving as the pre- and post-synaptic components. The vast majority of synapses in the mammalian nervous system are classical axo-dendritic synapses (axon synapsing upon a dendrite), however, a variety of other arrangements exist. These include but are not limited to[clarification needed] axo-axonic, dendro-dendritic, axo-secretory, axo-ciliary,[12] somato-dendritic, dendro-somatic, and somato-somatic synapses.[citation needed]
The axon can synapse onto a dendrite, onto a cell body, or onto another axon or axon terminal, as well as into the bloodstream or diffusely into the adjacent nervous tissue.
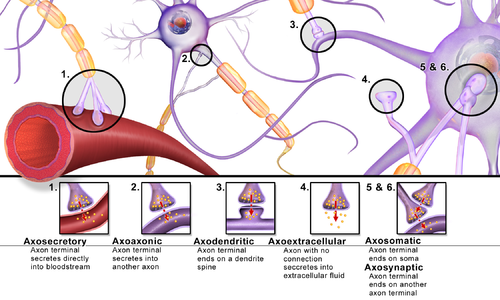
Different types of synapses
Role in memory[edit]
It is widely accepted that the synapse plays a role in the formation of memory. As neurotransmitters activate receptors across the synaptic cleft, the connection between the two neurons is strengthened when both neurons are active at the same time, as a result of the receptor’s signaling mechanisms. The strength of two connected neural pathways is thought to result in the storage of information, resulting in memory. This process of synaptic strengthening is known as long-term potentiation.[13]
By altering the release of neurotransmitters, the plasticity of synapses can be controlled in the presynaptic cell. The postsynaptic cell can be regulated by altering the function and number of its receptors. Changes in postsynaptic signaling are most commonly associated with a N-methyl-d-aspartic acid receptor (NMDAR)-dependent long-term potentiation (LTP) and long-term depression (LTD) due to the influx of calcium into the post-synaptic cell, which are the most analyzed forms of plasticity at excitatory synapses.[14]
Study models[edit]
For technical reasons, synaptic structure and function have been historically studied at unusually large model synapses, for example:
- Squid giant synapse
- Neuromuscular junction (NMJ), a cholinergic synapse in vertebrates, glutamatergic in insects
- Ciliary calyx in the ciliary ganglion of chicks[15]
- Calyx of Held in the brainstem
- Ribbon synapse in the retina
- Schaffer collateral synapses in the hippocampus. These synapses are small, but their pre- and postsynaptic neurons are well separated (CA3 and CA1, respectively).
Synaptic polarization[edit]
The function of neurons depends upon cell polarity. The distinctive structure of nerve cells allows action potentials to travel directionally (from dendrites to cell body down the axon), and for these signals to then be received and carried on by post-synaptic neurons or received by effector cells. Nerve cells have long been used as models for cellular polarization, and of particular interest are the mechanisms underlying the polarized localization of synaptic molecules. PIP2 signaling regulated by IMPase plays an integral role in synaptic polarity.
Phosphoinositides (PIP, PIP2, and PIP3) are molecules that have been shown to affect neuronal polarity.[16] A gene (ttx-7) was identified in Caenorhabditis elegans that encodes myo-inositol monophosphatase (IMPase), an enzyme that produces inositol by dephosphorylating inositol phosphate. Organisms with mutant ttx-7 genes demonstrated behavioral and localization defects, which were rescued by expression of IMPase. This led to the conclusion that IMPase is required for the correct localization of synaptic protein components.[17][18] The egl-8 gene encodes a homolog of phospholipase Cβ (PLCβ), an enzyme that cleaves PIP2. When ttx-7 mutants also had a mutant egl-8 gene, the defects caused by the faulty ttx-7 gene were largely reversed. These results suggest that PIP2 signaling establishes polarized localization of synaptic components in living neurons.[17]
Presynaptic modulation[edit]
Modulation of neurotransmitter release by G-protein-coupled receptors (GPCRs) is a prominent presynaptic mechanism for regulation of synaptic transmission. The activation of GPCRs located at the presynaptic terminal, can decrease the probability of neurotransmitter release. This presynaptic depression involves activation of Gi/o-type G-proteins that mediate different inhibitory mechanisms, including inhibition of voltage-gated calcium channels, activation of potassium channels, and direct inhibition of the vesicle fusion process. Endocannabinoids, synthesized in and released from postsynaptic neuronal elements, and their cognate receptors, including the (GPCR) CB1 receptor, located at the presynaptic terminal, are involved in this modulation by an retrograde signaling process, in which these compounds are synthesized in and released from postsynaptic neuronal elements, and travel back to the presynaptic terminal to act on the CB1 receptor for short-term (STD) or long-term synaptic depression (LTD), that cause a short or long lasting decrease in neurotransmitter release.[19]
Additional images[edit]
-

Diagram of the synapse. Please see learnbio.org for interactive version
-

A typical central nervous system synapse
-

The synapse and synaptic vesicle cycle
-

Major elements in chemical synaptic transmission




See also[edit]
- Active zone
- Autapse
- Exocytosis
- Immunological synapse
- Neurotransmitter vesicle
- Postsynaptic density
- Synaptopathy
References[edit]
- ^ a b Foster, M.; Sherrington, C.S. (1897). Textbook of Physiology, volume 3 (7th ed.). London: Macmillan. p. 929.
- ^ Perea, G.; Navarrete, M.; Araque, A. (August 2009). «Tripartite synapses: astrocytes process and control synaptic information». Trends in Neurosciences. Cambridge, MA: Cell Press. 32 (8): 421–431. doi:10.1016/j.tins.2009.05.001. PMID 19615761. S2CID 16355401.
- ^ Missler, M; Südhof, TC; Biederer, T (2012). «Synaptic cell adhesion». Cold Spring Harb Perspect Biol. 4 (4): a005694. doi:10.1101/cshperspect.a005694. PMC 3312681. PMID 22278667.
- ^ Elias, Lorin J.; Saucier, Deborah M. (2006). Neuropsychology: Clinical and Experimental Foundations. Boston: Pearson/Allyn & Bacon. ISBN 978-0-20534361-4. LCCN 2005051341. OCLC 61131869.
- ^ a b Harper, Douglas. «synapse». Online Etymology Dictionary.
- ^ Tansey, E.M. (1997). «Not committing barbarisms: Sherrington and the synapse, 1897». Brain Research Bulletin. Amsterdam: Elsevier. 44 (3): 211–212. doi:10.1016/S0361-9230(97)00312-2. PMID 9323432. S2CID 40333336.
The word synapse first appeared in 1897, in the seventh edition of Michael Foster’s Textbook of Physiology.
- ^ σύναψις, συνάπτειν, σύν, ἅπτειν. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project.
- ^ De Robertis, Eduardo D.P.; Bennett, H. Stanley (1955). «Some features of the submicroscopic morphology of synapses in frog and earthworm» (PDF). Journal of Biophysical and Biochemical Cytology. 1 (1): 47–58. doi:10.1083/jcb.1.1.47. PMC 2223594.
- ^ Palay, Sanford L.; Palade, George E. (1955). «The fine structure of neurons» (PDF). Journal of Biophysical and Biochemical Cytology. 1 (1): 69–88. doi:10.1083/jcb.1.1.69. PMC 2223597.
- ^ Palay, Sanford (1956). «Synapses in the central nervous system». J Biophys Biochem Cytol. 2 (4): 193–202. doi:10.1083/jcb.2.4.193. PMC 2229686. PMID 13357542.
- ^ Silverthorn, Dee Unglaub (2007). Human Physiology: An Integrated Approach. Illustration coordinator William C. Ober; illustrations by Claire W. Garrison; clinical consultant Andrew C. Silverthorn; contributions by Bruce R. Johnson (4th ed.). San Francisco: Pearson/Benjamin Cummings. p. 271. ISBN 978-0-8053-6851-2. LCCN 2005056517. OCLC 62742632.
- ^ Sheu, Shu-Hsien; Upadhyayula, Srigokul; Dupuy, Vincent; Pang, Song; Deng, Fei; Wan, Jinxia; Walpita, Deepika; Pasolli, H. Amalia; Houser, Justin; Sanchez-Martinez, Silvia; Brauchi, Sebastian E.; Banala, Sambashiva; Freeman, Melanie; Xu, C. Shan; Kirchhausen, Tom; Hess, Harald F.; Lavis, Luke; Li, Yulong; Chaumont-Dubel, Séverine; Clapham, David E. (1 September 2022). «A serotonergic axon-cilium synapse drives nuclear signaling to alter chromatin accessibility». Cell. 185 (18): 3390–3407.e18. doi:10.1016/j.cell.2022.07.026. ISSN 0092-8674. PMID 36055200. S2CID 251958800.
- University press release: «Scientists discover new kind of synapse in neurons’ tiny hairs». Howard Hughes Medical Institute via phys.org. Retrieved 19 October 2022.
- ^ Lynch, M. A. (January 1, 2004). «Long-Term Potentiation and Memory». Physiological Reviews. 84 (1): 87–136. doi:10.1152/physrev.00014.2003. PMID 14715912.
- ^ Krugers, Harm J.; Zhou, Ming; Joëls, Marian; Kindt, Merel (October 11, 2011). «Regulation of Excitatory Synapses and Fearful Memories by Stress Hormones». Frontiers in Behavioral Neuroscience. Switzerland: Frontiers Media SA. 5: 62. doi:10.3389/fnbeh.2011.00062. PMC 3190121. PMID 22013419.
- ^ Stanley, EF (1992). «The calyx-type synapse of the chick ciliary ganglion as a model of fast cholinergic transmission». Canadian Journal of Physiology and Pharmacology. 70 Suppl: S73-7. doi:10.1139/y92-246. PMID 1338300.
- ^ Arimura, Nariko; Kaibuchi, Kozo (December 22, 2005). «Key regulators in neuronal polarity». Neuron. Cambridge, MA: Cell Press. 48 (6): 881–884. doi:10.1016/j.neuron.2005.11.007. PMID 16364893.
- ^ a b Kimata, Tsubasa; Tanizawa, Yoshinori; Can, Yoko; et al. (June 1, 2012). «Synaptic Polarity Depends on Phosphatidylinositol Signaling Regulated by myo-Inositol Monophosphatase in Caenorhabditis elegans». Genetics. Bethesda, MD: Genetics Society of America. 191 (2): 509–521. doi:10.1534/genetics.111.137844. PMC 3374314. PMID 22446320.
- ^ Tanizawa, Yoshinori; Kuhara, Atsushi; Inada, Hitoshi; et al. (December 1, 2006). «Inositol monophosphatase regulates localization of synaptic components and behavior in the mature nervous system of C. elegans». Genes & Development. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. 20 (23): 3296–3310. doi:10.1101/gad.1497806. PMC 1686606. PMID 17158747.
- ^ Lovinger, David M. (2008), «Presynaptic Modulation by Endocannabinoids», in Südhof, Thomas C.; Starke, Klaus (eds.), Pharmacology of Neurotransmitter Release, Handbook of Experimental Pharmacology, vol. 184, Springer Berlin Heidelberg, pp. 435–477, doi:10.1007/978-3-540-74805-2_14, ISBN 9783540748052, PMID 18064422
Синапсыявляются специализированными
межклеточными контактами, которые
характерны только для нервной системы.
Различают химические и электрические
синапсы.Химический синапссостоит
их пресинаптической мембраны,
синаптической щели и постсинаптической
мембраны. Пресинаптическая мембранапредставляет собой участок плазмолеммы
аксона на его конце, который контактирует
с отростком или перикарионом другого
нейрона. Концевое расширение аксона
содержит митохондрии, микротрубочки
и промежуточные филаменты, а также
большое количество синаптических
пузырьков диаметром 4090
нм. Эти пузырьки заполненынейромедиатором– низкомолекулярным органическим
веществом, которое синтезируется в
перикарионе или в концевом расширении
аксона.Постсинаптическая мембранаобразована плазмолеммой второго
нейрона. Она содержит встроенные в
мембрану молекулы белкарецептора нейромедиатора.Синаптическая
щельпредставляет собой замкнутое
пространство между пресинаптической
и постсинаптической мембранами.
Приходящий
по аксону нейрона-передатчика к синапсу
нервный импульс вызывает слияние
синаптических пузырьков с пресинаптической
мембраной и выделение нейромедиатора
в синаптическую щель. Далее молекулы
нейромедиатора связываются рецепторами
постсинаптической мембраны, что
инициирует поступление в клетку ионов
натрия, деполяризацию постсинаптической
мембраны и возбуждение нейрона-приемника.
Если при связывании нейромедиатора
усиливается поступление в клетку ионов
хлора, наблюдается гиперполяризация
постсинаптической мембраны и торможение
нейрона-приемника. Для восстановления
способности синапса к повторной передаче
содержащийся в них медиатор подвергается
ферментативному разрушению. Способность
нейронов управлять передачей импульса
через синапс путем задержки его с
помощью других синапсов является
основополагающим принципом обработки
информации в нервной системе.
Каждый
нейрон вырабатывает свой специфический
нейромедиатор. Поэтому нейроны (и
соответствующие им синапсы) классифицируют
в зависимости от химической природы
секретируемого медиатора. В нервной
системе наиболее распространены
холинэргическиеиадренэргическиенейроны с ацетилхолином и норадреналином
в качестве медиаторов. Довольно часто
встречаются такжепептидэргическиенейроны, в которых медиаторами служат
различные пептиды,пуринэргическиенейроны с АТФ и ее производными иГАМК—эргическиенейроны, в
которых медиатором является-аминомасляная
кислота. В отличие от других ГАМК-эргические
нейроны и синапсы обычно вызывают
торможение.
Наиболее
полно изучены холинэргические нейроны,
к которым относятся среди прочих
мотонейроны спинного мозга. Ацетилхолин
в этих нейронах сконцентрирован в
синаптических пузырьках диаметром 40
нм. При возбуждении мотонейрона
ацетилхолин секретируется в синаптическую
щель, где связывается рецепторами
постсинаптической мембраны, принадлежащей
другим нейронам, мышечным волокнам или
гладкомышечным клеткам.
Межнейрональные
синапсы классифицируются также на
основе морфологических критериев.
Согласно этой классификации выделяют:
-
аксо-соматические
синапсы, которые связывают аксон одного
нейрона с перикарионом другого; -
аксо-дендритические
синапсы, связывающие аксон и дендрит; -
аксо-аксональные
синапсы, соединяющие аксоны двух
нейронов; -
сомато-соматические
синапсы, которые связывают перикарины
двух нейронов; -
дендро-дендритические
синапсы, связывающие дендриты двух
нейронов; -
дендро-соматические
синапсы, соединяющие дендрит и
перикарион.
Электрические
синапсы встречаются значительно
реже, чем химические. Они отличаются
почти полным слиянием мембран
контактирующих клеток. Передача нервного
импульса в электрических нейронах
происходит путем перехода волны
деполяризации с одной мембраны на
другую без участия нейромедиатора.
Этот тип синапсов обнаружен в спинном
мозге лягушки, в электрических органах
рыб и у ракообразных. Электрические
синапсы не способны обрабатывать
информацию так, как химические синапсы.