Нейрон (от
греч. neuron — нерв) — это структурно-функциональная
единица нервной системы. Эта клетка
имеет сложное строение, высоко
специализирована и по структуре содержит
ядро, тело клетки и отростки. В организме
человека насчитывается более 100 миллиардов
нейронов.
Функции
нейронов Как
и другие клетки, нейроны должны
обеспечивать поддержание собственной
структуры и функций, приспосабливаться
к изменяющимся условиям и оказывать
регулирующее влияние на соседние клетки.
Однако основная функция нейронов — это
переработка информации: получение,
проведение и передача другим клеткам.
Получение информации происходит через
синапсы с рецепторами сенсорных органов
или другими нейронами, или непосредственно
из внешней среды с помощью специализированных
дендритов. Проведение информации
происходит по аксонам, передача — через
синапсы.
Строение
нейрона
Тело
клетки Тело
нервной клетки состоит из протоплазмы
(цитоплазмы и ядра), снаружи ограничена
мембраной из двойного слоя липидов
(билипидный слой). Липиды состоят из
гидрофильных головок и гидрофобных
хвостов, расположены гидрофобными
хвостами друг к другу, образуя гидрофобный
слой, который пропускает только
жирорастворимые вещества (напр. кислород
и углекислый газ). На мембране находятся
белки: на поверхности (в форме глобул),
на которых можно наблюдать наросты
полисахаридов (гликокаликс), благодаря
которым клетка воспринимает внешнее
раздражение, и интегральные белки,
пронизывающие мембрану насквозь, в них
находятся ионные каналы.
Нейрон
состоит из тела диаметром от 3 до 100 мкм,
содержащего ядро (с большим количеством
ядерных пор) и органеллы (в том числе
сильно развитый шероховатый ЭПР с
активными рибосомами, аппарат Гольджи),
а также из отростков. Выделяют два вида
отростков: дендриты и аксон. Нейрон
имеет развитый цитоскелет, проникающий
в его отростки. Цитоскелет поддерживает
форму клетки, его нити служат «рельсами»
для транспорта органелл и упакованных
в мембранные пузырьки веществ (например,
нейромедиаторов). В теле нейрона
выявляется развитый синтетический
аппарат, гранулярная ЭПС нейрона
окрашивается базофильно и известна под
названием «тигроид». Тигроид проникает
в начальные отделы дендритов, но
располагается на заметном расстоянии
от начала аксона, что служит гистологическим
признаком аксона. Различается антероградный
(от тела) и ретроградный (к телу) аксонный
транспорт.
Дендриты
и аксон
Аксон
— обычно длинный отросток, приспособленный
для проведения возбуждения от тела
нейрона. Дендриты — как правило, короткие
и сильно разветвлённые отростки, служащие
главным местом образования влияющих
на нейрон возбуждающих и тормозных
синапсов (разные нейроны имеют различное
соотношение длины аксона и дендритов).
Нейрон может иметь несколько дендритов
и обычно только один аксон. Один нейрон
может иметь связи со многими (до 20-и
тысяч) другими нейронами. Дендриты
делятся дихотомически, аксоны же дают
коллатерали. В узлах ветвления обычно
сосредоточены митохондрии. Дендриты
не имеют миелиновой оболочки, аксоны
же могут её иметь. Местом генерации
возбуждения у большинства нейронов
является аксонный холмик — образование
в месте отхождения аксона от тела. У
всех нейронов эта зона называется
триггерной.
Синапс Синапс
— место контакта между двумя нейронами
или между нейроном и получающей сигнал
эффекторной клеткой. Служит для передачи
нервного импульса между двумя клетками,
причём в ходе синаптической передачи
амплитуда и частота сигнала могут
регулироваться. Одни синапсы вызывают
деполяризацию нейрона, другие —
гиперполяризацию; первые являются
возбуждающими, вторые — тормозящими.
Обычно для возбуждения нейрона необходимо
раздражение от нескольких возбуждающих
синапсов.
Структурная
классификация нейронов
На
основании числа и расположения дендритов
и аксона нейроны делятся на безаксонные,
униполярные нейроны, псевдоуниполярные
нейроны, биполярные нейроны и мультиполярные
(много дендритных стволов, обычно
эфферентные) нейроны.
Безаксонные
нейроны —
небольшие клетки, сгруппированы вблизи
спинного мозга в межпозвоночных ганглиях,
не имеющие анатомических признаков
разделения отростков на дендриты и
аксоны. Все отростки у клетки очень
похожи. Функциональное назначение
безаксонных нейронов слабо изучено.
Униполярные
нейроны —
нейроны с одним отростком, присутствуют,
например в сенсорном ядре тройничного
нерва в среднем мозге.
Биполярные
нейроны —
нейроны, имеющие один аксон и один
дендрит, расположенные в специализированных
сенсорных органах — сетчатке глаза,
обонятельном эпителии и луковице,
слуховом и вестибулярном ганглиях;
Мультиполярные
нейроны —
Нейроны с одним аксоном и несколькими
дендритами. Данный вид нервных клеток
преобладает в центральной нервной
системе
Псевдоуниполярные
нейроны —
являются уникальными в своём роде. От
тела отходит один отросток, который
сразу же Т-образно делится. Весь этот
единый тракт покрыт миелиновой оболочкой
и структурно представляет собой аксон,
хотя по одной из ветвей возбуждение
идёт не от, а к телу нейрона. Структурно
дендритами являются разветвления на
конце этого (периферического) отростка.
Триггерной зоной является начало этого
разветвления (т. е. находится вне тела
клетки). Такие нейроны встречаются в
спинальных ганглиях.
Функциональная
классификация нейронов По
положению в рефлекторной дуге различают
афферентные нейроны (чувствительные
нейроны), эфферентные нейроны (часть из
них называется двигательными нейронами,
иногда это не очень точное название
распространяется на всю группу эфферентов)
и интернейроны (вставочные нейроны).
Афферентные
нейроны (чувствительный,
сенсорный или рецепторный). К нейронам
данного типа относятся первичные клетки
органов чувств и псевдоуниполярные
клетки, у которых дендриты имеют свободные
окончания.
Эфферентные
нейроны (эффекторный,
двигательный или моторный). К нейронам
данного типа относятся конечные нейроны
— ультиматные и предпоследние –
неультиматные.
Ассоциативные
нейроны (вставочные
или интернейроны) — эта группа нейронов
осуществляет связь между эфферентными
и афферентными, их делят на комиссуральные
и проекционные (головной мозг).
Морфологическая
классификация нейронов Морфологическое
строение нейронов многообразно. В связи
с этим при классификации нейронов
применяют несколько принципов:
учитывают
размеры и форму тела нейрона,
количество
и характер ветвления отростков,
длину
нейрона и наличие специализированные
оболочки.
По
форме клетки, нейроны могут быть
сферическими, зернистыми, звездчатыми,
пирамидными, грушевидными, веретеновидными,
неправильными и т. д. Размер тела нейрона
варьирует от 5 мкм у малых зернистых
клеток до 120-150 мкм у гигантских пирамидных
нейронов. Длина нейрона у человека
составляет от 150 мкм до 120 см. По количеству
отростков выделяют следующие
морфологические типы нейронов: —
униполярные (с одним отростком) нейроциты,
присутствующие, например, в сенсорном
ядре тройничного нерва в среднем мозге;
— псевдоуниполярные клетки, сгруппированные
вблизи спинного мозга в межпозвоночных
ганглиях; — биполярные нейроны (имеют
один аксон и один дендрит), расположенные
в специализированных сенсорных органах
— сетчатке глаза, обонятельном эпителии
и луковице, слуховом и вестибулярном
ганглиях; — мультиполярные нейроны
(имеют один аксон и несколько дендритов),
преобладающие в ЦНС.
Развитие
и рост нейрона Нейрон
развивается из небольшой клетки —
предшественницы, которая перестаёт
делиться ещё до того, как выпустит свои
отростки. (Однако, вопрос о делении
нейронов в настоящее время остаётся
дискуссионным.) Как правило, первым
начинает расти аксон, а дендриты
образуются позже. На конце развивающегося
отростка нервной клетки появляется
утолщение неправильной формы, которое,
видимо, и прокладывает путь через
окружающую ткань. Это утолщение называется
конусом роста нервной клетки. Он состоит
из уплощенной части отростка нервной
клетки с множеством тонких шипиков.
Микрошипики имеют толщину от 0,1 до 0,2
мкм и могут достигать 50 мкм в длину,
широкая и плоская область конуса роста
имеет ширину и длину около 5 мкм, хотя
форма её может изменяться. Промежутки
между микрошипиками конуса роста покрыты
складчатой мембраной. Микрошипики
находятся в постоянном движении —
некоторые втягиваются в конус роста,
другие удлиняются, отклоняются в разные
стороны, прикасаются к субстрату и могут
прилипать к нему. Конус роста заполнен
мелкими, иногда соединёнными друг с
другом, мембранными пузырьками
неправильной формы. Непосредственно
под складчатыми участками мембраны и
в шипиках находится плотная масса
перепутанных актиновых филаментов.
Конус роста содержит также митохондрии,
микротрубочки и нейрофиламенты, имеющиеся
в теле нейрона. Вероятно, микротрубочки
и нейрофиламенты удлиняются главным
образом за счёт добавления вновь
синтезированных субъединиц у основания
отростка нейрона. Они продвигаются со
скоростью около миллиметра в сутки, что
соответствует скорости медленного
аксонного транспорта в зрелом нейроне.
Поскольку
примерно такова и средняя скорость
продвижения конуса роста, возможно, что
во время роста отростка нейрона в его
дальнем конце не происходит ни сборки,
ни разрушения микротрубочек и
нейрофиламентов. Новый мембранный
материал добавляется, видимо, у окончания.
Конус роста — это область быстрого
экзоцитоза и эндоцитоза, о чём
свидетельствует множество находящихся
здесь пузырьков. Мелкие мембранные
пузырьки переносятся по отростку нейрона
от тела клетки к конусу роста с потоком
быстрого аксонного транспорта. Мембранный
материал, видимо, синтезируется в теле
нейрона, переносится к конусу роста в
виде пузырьков и включается здесь в
плазматическую мембрану путём экзоцитоза,
удлиняя таким образом отросток нервной
клетки. Росту аксонов и дендритов обычно
предшествует фаза миграции нейронов,
когда незрелые нейроны расселяются и
находят себе постоянное место.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Not to be confused with Neutron.
| Neuron | |
|---|---|

Anatomy of a multipolar neuron |
|
| Identifiers | |
| MeSH | D009474 |
| NeuroLex ID | sao1417703748 |
| TA98 | A14.0.00.002 |
| TH | H2.00.06.1.00002 |
| FMA | 54527 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A neuron, neurone, or nerve cell is an electrically excitable cell that fires electric signals called action potentials. Neurons communicate with other cells via synapses — specialized connections that commonly use minute amounts of chemical neurotransmitters to pass the electric signal from the presynaptic neuron to the target cell through the synaptic gap. The neuron is the main component of nervous tissue in all animals except sponges and placozoa. Non-animals like plants and fungi do not have nerve cells.
Neurons are typically classified into three types based on their function. Sensory neurons respond to stimuli such as touch, sound, or light that affect the cells of the sensory organs, and they send signals to the spinal cord or brain. Motor neurons receive signals from the brain and spinal cord to control everything from muscle contractions to glandular output. Interneurons connect neurons to other neurons within the same region of the brain or spinal cord. When multiple neurons are functionally connected together, they form what is called a neural circuit.
A typical neuron consists of a cell body (soma), dendrites, and a single axon. The soma is a compact structure, and the axon and dendrites are filaments extruding from the soma. Dendrites typically branch profusely and extend a few hundred micrometers from the soma. The axon leaves the soma at a swelling called the axon hillock and travels for as far as 1 meter in humans or more in other species. It branches but usually maintains a constant diameter. At the farthest tip of the axon’s branches are axon terminals, where the neuron can transmit a signal across the synapse to another cell. Neurons may lack dendrites or have no axon. The term neurite is used to describe either a dendrite or an axon, particularly when the cell is undifferentiated.
Most neurons receive signals via the dendrites and soma and send out signals down the axon. At the majority of synapses, signals cross from the axon of one neuron to a dendrite of another. However, synapses can connect an axon to another axon or a dendrite to another dendrite.
The signaling process is partly electrical and partly chemical. Neurons are electrically excitable, due to maintenance of voltage gradients across their membranes. If the voltage changes by a large enough amount over a short interval, the neuron generates an all-or-nothing electrochemical pulse called an action potential. This potential travels rapidly along the axon and activates synaptic connections as it reaches them. Synaptic signals may be excitatory or inhibitory, increasing or reducing the net voltage that reaches the soma.
In most cases, neurons are generated by neural stem cells during brain development and childhood. Neurogenesis largely ceases during adulthood in most areas of the brain.
Nervous system[edit]
Schematic of an anatomically accurate single pyramidal neuron, the primaryious excitatory neuron of cerebral cortex, with a synaptic connection from an incoming axon onto a dendritic spine
Neurons are the primary components of the nervous system, along with the glial cells that give them structural and metabolic support.[1] The nervous system is made up of the central nervous system, which includes the brain and spinal cord, and the peripheral nervous system, which includes the autonomic and somatic nervous systems.[2] In vertebrates, the majority of neurons belong to the central nervous system, but some reside in peripheral ganglia, and many sensory neurons are situated in sensory organs such as the retina and cochlea.
Axons may bundle into fascicles that make up the nerves in the peripheral nervous system (like strands of wire make up cables). Bundles of axons in the central nervous system are called tracts.
Anatomy and histology[edit]

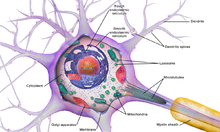
Diagram of the components of a neuron
Neurons are highly specialized for the processing and transmission of cellular signals. Given their diversity of functions performed in different parts of the nervous system, there is a wide variety in their shape, size, and electrochemical properties. For instance, the soma of a neuron can vary from 4 to 100 micrometers in diameter.[3]
- The soma is the body of the neuron. As it contains the nucleus, most protein synthesis occurs here. The nucleus can range from 3 to 18 micrometers in diameter.[4]
- The dendrites of a neuron are cellular extensions with many branches. This overall shape and structure are referred to metaphorically as a dendritic tree. This is where the majority of input to the neuron occurs via the dendritic spine.
- The axon is a finer, cable-like projection that can extend tens, hundreds, or even tens of thousands of times the diameter of the soma in length. The axon primarily carries nerve signals away from the soma and carries some types of information back to it. Many neurons have only one axon, but this axon may—and usually will—undergo extensive branching, enabling communication with many target cells. The part of the axon where it emerges from the soma is called the axon hillock. Besides being an anatomical structure, the axon hillock also has the greatest density of voltage-dependent sodium channels. This makes it the most easily excited part of the neuron and the spike initiation zone for the axon. In electrophysiological terms, it has the most negative threshold potential.
- While the axon and axon hillock are generally involved in information outflow, this region can also receive input from other neurons.
- The axon terminal is found at the end of the axon farthest from the soma and contains synapses. Synaptic boutons are specialized structures where neurotransmitter chemicals are released to communicate with target neurons. In addition to synaptic boutons at the axon terminal, a neuron may have en passant boutons, which are located along the length of the axon.
The accepted view of the neuron attributes dedicated functions to its various anatomical components; however, dendrites and axons often act in ways contrary to their so-called main function.[5]
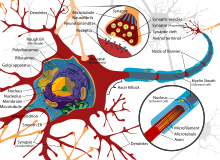
Diagram of a typical myelinated vertebrate motor neuron
Axons and dendrites in the central nervous system are typically only about one micrometer thick, while some in the peripheral nervous system are much thicker. The soma is usually about 10–25 micrometers in diameter and often is not much larger than the cell nucleus it contains. The longest axon of a human motor neuron can be over a meter long, reaching from the base of the spine to the toes.
Sensory neurons can have axons that run from the toes to the posterior column of the spinal cord, over 1.5 meters in adults. Giraffes have single axons several meters in length running along the entire length of their necks. Much of what is known about axonal function comes from studying the squid giant axon, an ideal experimental preparation because of its relatively immense size (0.5–1 millimeter thick, several centimeters long).
Fully differentiated neurons are permanently postmitotic[6] however, stem cells present in the adult brain may regenerate functional neurons throughout the life of an organism (see neurogenesis). Astrocytes are star-shaped glial cells. They have been observed to turn into neurons by virtue of their stem cell-like characteristic of pluripotency.
Membrane[edit]
Like all animal cells, the cell body of every neuron is enclosed by a plasma membrane, a bilayer of lipid molecules with many types of protein structures embedded in it.[7] A lipid bilayer is a powerful electrical insulator, but in neurons, many of the protein structures embedded in the membrane are electrically active. These include ion channels that permit electrically charged ions to flow across the membrane and ion pumps that chemically transport ions from one side of the membrane to the other. Most ion channels are permeable only to specific types of ions. Some ion channels are voltage gated, meaning that they can be switched between open and closed states by altering the voltage difference across the membrane. Others are chemically gated, meaning that they can be switched between open and closed states by interactions with chemicals that diffuse through the extracellular fluid. The ion materials include sodium, potassium, chloride, and calcium. The interactions between ion channels and ion pumps produce a voltage difference across the membrane, typically a bit less than 1/10 of a volt at baseline. This voltage has two functions: first, it provides a power source for an assortment of voltage-dependent protein machinery that is embedded in the membrane; second, it provides a basis for electrical signal transmission between different parts of the membrane.
Histology and internal structure[edit]

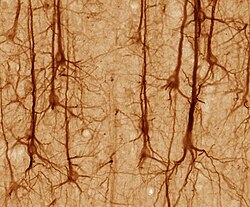
Golgi-stained neurons in human hippocampal tissue

Actin filaments in a mouse cortical neuron in culture
Numerous microscopic clumps called Nissl bodies (or Nissl substance) are seen when nerve cell bodies are stained with a basophilic («base-loving») dye. These structures consist of rough endoplasmic reticulum and associated ribosomal RNA. Named after German psychiatrist and neuropathologist Franz Nissl (1860–1919), they are involved in protein synthesis and their prominence can be explained by the fact that nerve cells are very metabolically active. Basophilic dyes such as aniline or (weakly) haematoxylin[8] highlight negatively charged components, and so bind to the phosphate backbone of the ribosomal RNA.
The cell body of a neuron is supported by a complex mesh of structural proteins called neurofilaments, which together with neurotubules (neuronal microtubules) are assembled into larger neurofibrils.[9] Some neurons also contain pigment granules, such as neuromelanin (a brownish-black pigment that is byproduct of synthesis of catecholamines), and lipofuscin (a yellowish-brown pigment), both of which accumulate with age.[10][11][12] Other structural proteins that are important for neuronal function are actin and the tubulin of microtubules. Class III β-tubulin is found almost exclusively in neurons. Actin is predominately found at the tips of axons and dendrites during neuronal development. There the actin dynamics can be modulated via an interplay with microtubule.[13]
There are different internal structural characteristics between axons and dendrites. Typical axons almost never contain ribosomes, except some in the initial segment. Dendrites contain granular endoplasmic reticulum or ribosomes, in diminishing amounts as the distance from the cell body increases.
Classification[edit]

Neurons vary in shape and size and can be classified by their morphology and function.[15] The anatomist Camillo Golgi grouped neurons into two types; type I with long axons used to move signals over long distances and type II with short axons, which can often be confused with dendrites. Type I cells can be further classified by the location of the soma. The basic morphology of type I neurons, represented by spinal motor neurons, consists of a cell body called the soma and a long thin axon covered by a myelin sheath. The dendritic tree wraps around the cell body and receives signals from other neurons. The end of the axon has branching axon terminals that release neurotransmitters into a gap called the synaptic cleft between the terminals and the dendrites of the next neuron.[citation needed]
Structural classification[edit]
Polarity[edit]
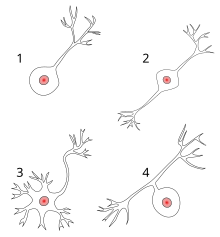
Most neurons can be anatomically characterized as:[citation needed]
- Unipolar: single process
- Bipolar: 1 axon and 1 dendrite
- Multipolar: 1 axon and 2 or more dendrites
- Golgi I: neurons with long-projecting axonal processes; examples are pyramidal cells, Purkinje cells, and anterior horn cells
- Golgi II: neurons whose axonal process projects locally; the best example is the granule cell
- Anaxonic: where the axon cannot be distinguished from the dendrite(s)
- Pseudounipolar: 1 process which then serves as both an axon and a dendrite
Other[edit]
Some unique neuronal types can be identified according to their location in the nervous system and distinct shape. Some examples are:[citation needed]
- Basket cells, interneurons that form a dense plexus of terminals around the soma of target cells, found in the cortex and cerebellum
- Betz cells, large motor neurons
- Lugaro cells, interneurons of the cerebellum
- Medium spiny neurons, most neurons in the corpus striatum
- Purkinje cells, huge neurons in the cerebellum, a type of Golgi I multipolar neuron
- Pyramidal cells, neurons with triangular soma, a type of Golgi I
- Renshaw cells, neurons with both ends linked to alpha motor neurons
- Unipolar brush cells, interneurons with unique dendrite ending in a brush-like tuft
- Granule cells, a type of Golgi II neuron
- Anterior horn cells, motoneurons located in the spinal cord
- Spindle cells, interneurons that connect widely separated areas of the brain
Functional classification[edit]
Direction[edit]
- Afferent neurons convey information from tissues and organs into the central nervous system and are also called sensory neurons.
- Efferent neurons (motor neurons) transmit signals from the central nervous system to the effector cells.
- Interneurons connect neurons within specific regions of the central nervous system.
Afferent and efferent also refer generally to neurons that, respectively, bring information to or send information from the brain.
Action on other neurons[edit]
A neuron affects other neurons by releasing a neurotransmitter that binds to chemical receptors. The effect upon the postsynaptic neuron is determined by the type of receptor that is activated, not by the presynaptic neuron or by the neurotransmitter. A neurotransmitter can be thought of as a key, and a receptor as a lock: the same neurotransmitter can activate multiple types of receptors. Receptors can be classified broadly as excitatory (causing an increase in firing rate), inhibitory (causing a decrease in firing rate), or modulatory (causing long-lasting effects not directly related to firing rate).[citation needed]
The two most common (90%+) neurotransmitters in the brain, glutamate and GABA, have largely consistent actions. Glutamate acts on several types of receptors, and has effects that are excitatory at ionotropic receptors and a modulatory effect at metabotropic receptors. Similarly, GABA acts on several types of receptors, but all of them have inhibitory effects (in adult animals, at least). Because of this consistency, it is common for neuroscientists to refer to cells that release glutamate as «excitatory neurons», and cells that release GABA as «inhibitory neurons». Some other types of neurons have consistent effects, for example, «excitatory» motor neurons in the spinal cord that release acetylcholine, and «inhibitory» spinal neurons that release glycine.[citation needed]
The distinction between excitatory and inhibitory neurotransmitters is not absolute. Rather, it depends on the class of chemical receptors present on the postsynaptic neuron. In principle, a single neuron, releasing a single neurotransmitter, can have excitatory effects on some targets, inhibitory effects on others, and modulatory effects on others still. For example, photoreceptor cells in the retina constantly release the neurotransmitter glutamate in the absence of light. So-called OFF bipolar cells are, like most neurons, excited by the released glutamate. However, neighboring target neurons called ON bipolar cells are instead inhibited by glutamate, because they lack typical ionotropic glutamate receptors and instead express a class of inhibitory metabotropic glutamate receptors.[16] When light is present, the photoreceptors cease releasing glutamate, which relieves the ON bipolar cells from inhibition, activating them; this simultaneously removes the excitation from the OFF bipolar cells, silencing them.[citation needed]
It is possible to identify the type of inhibitory effect a presynaptic neuron will have on a postsynaptic neuron, based on the proteins the presynaptic neuron expresses. Parvalbumin-expressing neurons typically dampen the output signal of the postsynaptic neuron in the visual cortex, whereas somatostatin-expressing neurons typically block dendritic inputs to the postsynaptic neuron.[17]
Discharge patterns[edit]
Neurons have intrinsic electroresponsive properties like intrinsic transmembrane voltage oscillatory patterns.[18] So neurons can be classified according to their electrophysiological characteristics:
- Tonic or regular spiking. Some neurons are typically constantly (tonically) active, typically firing at a constant frequency. Example: interneurons in neurostriatum.
- Phasic or bursting. Neurons that fire in bursts are called phasic.
- Fast spiking. Some neurons are notable for their high firing rates, for example some types of cortical inhibitory interneurons, cells in globus pallidus, retinal ganglion cells.[19][20]
Neurotransmitter[edit]
![]()
Synaptic vesicles containing neurotransmitters
Neurotransmitters are chemical messengers passed from one neuron to another neuron or to a muscle cell or gland cell.
- Cholinergic neurons – acetylcholine. Acetylcholine is released from presynaptic neurons into the synaptic cleft. It acts as a ligand for both ligand-gated ion channels and metabotropic (GPCRs) muscarinic receptors. Nicotinic receptors are pentameric ligand-gated ion channels composed of alpha and beta subunits that bind nicotine. Ligand binding opens the channel causing influx of Na+ depolarization and increases the probability of presynaptic neurotransmitter release. Acetylcholine is synthesized from choline and acetyl coenzyme A.
- Adrenergic neurons – noradrenaline. Noradrenaline (norepinephrine) is released from most postganglionic neurons in the sympathetic nervous system onto two sets of GPCRs: alpha adrenoceptors and beta adrenoceptors. Noradrenaline is one of the three common catecholamine neurotransmitter, and the most prevalent of them in the peripheral nervous system; as with other catecholamines, it is synthesised from tyrosine.
- GABAergic neurons – gamma aminobutyric acid. GABA is one of two neuroinhibitors in the central nervous system (CNS), along with glycine. GABA has a homologous function to ACh, gating anion channels that allow Cl− ions to enter the post synaptic neuron. Cl− causes hyperpolarization within the neuron, decreasing the probability of an action potential firing as the voltage becomes more negative (for an action potential to fire, a positive voltage threshold must be reached). GABA is synthesized from glutamate neurotransmitters by the enzyme glutamate decarboxylase.
- Glutamatergic neurons – glutamate. Glutamate is one of two primary excitatory amino acid neurotransmitters, along with aspartate. Glutamate receptors are one of four categories, three of which are ligand-gated ion channels and one of which is a G-protein coupled receptor (often referred to as GPCR).
-
- AMPA and Kainate receptors function as cation channels permeable to Na+ cation channels mediating fast excitatory synaptic transmission.
- NMDA receptors are another cation channel that is more permeable to Ca2+. The function of NMDA receptors depend on glycine receptor binding as a co-agonist within the channel pore. NMDA receptors do not function without both ligands present.
- Metabotropic receptors, GPCRs modulate synaptic transmission and postsynaptic excitability.
- Glutamate can cause excitotoxicity when blood flow to the brain is interrupted, resulting in brain damage. When blood flow is suppressed, glutamate is released from presynaptic neurons, causing greater NMDA and AMPA receptor activation than normal outside of stress conditions, leading to elevated Ca2+ and Na+ entering the post synaptic neuron and cell damage. Glutamate is synthesized from the amino acid glutamine by the enzyme glutamate synthase.
- Dopaminergic neurons—dopamine. Dopamine is a neurotransmitter that acts on D1 type (D1 and D5) Gs-coupled receptors, which increase cAMP and PKA, and D2 type (D2, D3, and D4) receptors, which activate Gi-coupled receptors that decrease cAMP and PKA. Dopamine is connected to mood and behavior and modulates both pre- and post-synaptic neurotransmission. Loss of dopamine neurons in the substantia nigra has been linked to Parkinson’s disease. Dopamine is synthesized from the amino acid tyrosine. Tyrosine is catalyzed into levodopa (or L-DOPA) by tyrosine hydroxlase, and levodopa is then converted into dopamine by the aromatic amino acid decarboxylase.
- Serotonergic neurons—serotonin. Serotonin (5-Hydroxytryptamine, 5-HT) can act as excitatory or inhibitory. Of its four 5-HT receptor classes, 3 are GPCR and 1 is a ligand-gated cation channel. Serotonin is synthesized from tryptophan by tryptophan hydroxylase, and then further by decarboxylase. A lack of 5-HT at postsynaptic neurons has been linked to depression. Drugs that block the presynaptic serotonin transporter are used for treatment, such as Prozac and Zoloft.
- Purinergic neurons—ATP. ATP is a neurotransmitter acting at both ligand-gated ion channels (P2X receptors) and GPCRs (P2Y) receptors. ATP is, however, best known as a cotransmitter. Such purinergic signalling can also be mediated by other purines like adenosine, which particularly acts at P2Y receptors.
- Histaminergic neurons—histamine. Histamine is a monoamine neurotransmitter and neuromodulator. Histamine-producing neurons are found in the tuberomammillary nucleus of the hypothalamus.[21] Histamine is involved in arousal and regulating sleep/wake behaviors.
Multimodel classification[edit]
Since 2012 there has been a push from the cellular and computational neuroscience community to come up with a universal classification of neurons that will apply to all neurons in the brain as well as across species. This is done by considering the three essential qualities of all neurons: electrophysiology, morphology, and the individual transcriptome of the cells. Besides being universal this classification has the advantage of being able to classify astrocytes as well. A method called Patch-Seq in which all three qualities can be measured at once is used extensively by the Allen Institute for Brain Science.[22]
Connectivity[edit]

A signal propagating down an axon to the cell body and dendrites of the next cell

Neurons communicate with each other via synapses, where either the axon terminal of one cell contacts another neuron’s dendrite, soma or, less commonly, axon. Neurons such as Purkinje cells in the cerebellum can have over 1000 dendritic branches, making connections with tens of thousands of other cells; other neurons, such as the magnocellular neurons of the supraoptic nucleus, have only one or two dendrites, each of which receives thousands of synapses.
Synapses can be excitatory or inhibitory, either increasing or decreasing activity in the target neuron, respectively. Some neurons also communicate via electrical synapses, which are direct, electrically conductive junctions between cells.[23]
When an action potential reaches the axon terminal, it opens voltage-gated calcium channels, allowing calcium ions to enter the terminal. Calcium causes synaptic vesicles filled with neurotransmitter molecules to fuse with the membrane, releasing their contents into the synaptic cleft. The neurotransmitters diffuse across the synaptic cleft and activate receptors on the postsynaptic neuron. High cytosolic calcium in the axon terminal triggers mitochondrial calcium uptake, which, in turn, activates mitochondrial energy metabolism to produce ATP to support continuous neurotransmission.[24]
An autapse is a synapse in which a neuron’s axon connects to its own dendrites.
The human brain has some 8.6 x 1010 (eighty six billion) neurons.[25] Each neuron has on average 7,000 synaptic connections to other neurons. It has been estimated that the brain of a three-year-old child has about 1015 synapses (1 quadrillion). This number declines with age, stabilizing by adulthood. Estimates vary for an adult, ranging from 1014 to 5 x 1014 synapses (100 to 500 trillion).[26]
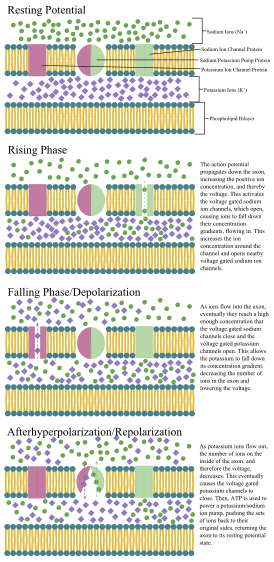
An annotated diagram of the stages of an action potential propagating down an axon including the role of ion concentration and pump and channel proteins
Nonelectrochemical signaling[edit]
Beyond electrical and chemical signaling, studies suggest neurons in healthy human brains can also communicate through:
- force generated by the enlargement of dendritic spines[27]
- the transfer of proteins – transneuronally transported proteins (TNTPs)[28][29]
They can also get modulated by input from the environment and hormones released from other parts of the organism,[30] which could be influenced more or less directly by neurons. This also applies to neurotrophins such as BDNF. The gut microbiome is also connected with the brain.[31]
Neurons also communicate with microglia, the brain’s main immune cells via specialised contact sites, called «somatic junctions». These connections enable microglia to constantly monitor and regulate neuronal functions, and exert neuroprotection, when needed.[32]
Mechanisms for propagating action potentials[edit]
In 1937 John Zachary Young suggested that the squid giant axon could be used to study neuronal electrical properties.[33] It is larger than but similar to human neurons, making it easier to study. By inserting electrodes into the squid giant axons, accurate measurements were made of the membrane potential.
The cell membrane of the axon and soma contain voltage-gated ion channels that allow the neuron to generate and propagate an electrical signal (an action potential). Some neurons also generate subthreshold membrane potential oscillations. These signals are generated and propagated by charge-carrying ions including sodium (Na+), potassium (K+), chloride (Cl−), and calcium (Ca2+).
Several stimuli can activate a neuron leading to electrical activity, including pressure, stretch, chemical transmitters, and changes of the electric potential across the cell membrane.[34] Stimuli cause specific ion-channels within the cell membrane to open, leading to a flow of ions through the cell membrane, changing the membrane potential. Neurons must maintain the specific electrical properties that define their neuron type.[35]
Thin neurons and axons require less metabolic expense to produce and carry action potentials, but thicker axons convey impulses more rapidly. To minimize metabolic expense while maintaining rapid conduction, many neurons have insulating sheaths of myelin around their axons. The sheaths are formed by glial cells: oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system. The sheath enables action potentials to travel faster than in unmyelinated axons of the same diameter, whilst using less energy. The myelin sheath in peripheral nerves normally runs along the axon in sections about 1 mm long, punctuated by unsheathed nodes of Ranvier, which contain a high density of voltage-gated ion channels. Multiple sclerosis is a neurological disorder that results from demyelination of axons in the central nervous system.
Some neurons do not generate action potentials, but instead generate a graded electrical signal, which in turn causes graded neurotransmitter release. Such non-spiking neurons tend to be sensory neurons or interneurons, because they cannot carry signals long distances.
Neural coding[edit]
Neural coding is concerned with how sensory and other information is represented in the brain by neurons. The main goal of studying neural coding is to characterize the relationship between the stimulus and the individual or ensemble neuronal responses, and the relationships among the electrical activities of the neurons within the ensemble.[36] It is thought that neurons can encode both digital and analog information.[37]
All-or-none principle[edit]
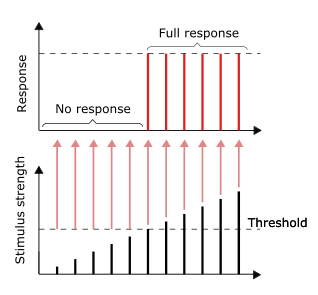
As long as the stimulus reaches the threshold, the full response would be given. Larger stimulus does not result in a larger response, vice versa.[38]: 31
The conduction of nerve impulses is an example of an all-or-none response. In other words, if a neuron responds at all, then it must respond completely. Greater intensity of stimulation, like brighter image/louder sound, does not produce a stronger signal, but can increase firing frequency.[38]: 31 Receptors respond in different ways to stimuli. Slowly adapting or tonic receptors respond to steady stimulus and produce a steady rate of firing. Tonic receptors most often respond to increased intensity of stimulus by increasing their firing frequency, usually as a power function of stimulus plotted against impulses per second. This can be likened to an intrinsic property of light where greater intensity of a specific frequency (color) requires more photons, as the photons can’t become «stronger» for a specific frequency.
Other receptor types include quickly adapting or phasic receptors, where firing decreases or stops with steady stimulus; examples include skin which, when touched causes neurons to fire, but if the object maintains even pressure, the neurons stop firing. The neurons of the skin and muscles that are responsive to pressure and vibration have filtering accessory structures that aid their function.
The pacinian corpuscle is one such structure. It has concentric layers like an onion, which form around the axon terminal. When pressure is applied and the corpuscle is deformed, mechanical stimulus is transferred to the axon, which fires. If the pressure is steady, stimulus ends; thus, typically these neurons respond with a transient depolarization during the initial deformation and again when the pressure is removed, which causes the corpuscle to change shape again. Other types of adaptation are important in extending the function of a number of other neurons.[39]
Etymology and spelling[edit]
The German anatomist Heinrich Wilhelm Waldeyer introduced the term neuron in 1891,[40] based on the ancient Greek νεῦρον neuron ‘sinew, cord, nerve’.[41]
The word was adopted in French with the spelling neurone. That spelling was also used by many writers in English,[42] but has now become rare in American usage and uncommon in British usage.[43][41]
History[edit]

The neuron’s place as the primary functional unit of the nervous system was first recognized in the late 19th century through the work of the Spanish anatomist Santiago Ramón y Cajal.[44]
To make the structure of individual neurons visible, Ramón y Cajal improved a silver staining process that had been developed by Camillo Golgi.[44] The improved process involves a technique called «double impregnation» and is still in use.
In 1888 Ramón y Cajal published a paper about the bird cerebellum. In this paper, he stated that he could not find evidence for anastomosis between axons and dendrites and called each nervous element «an absolutely autonomous canton.»[44][40] This became known as the neuron doctrine, one of the central tenets of modern neuroscience.[44]
In 1891, the German anatomist Heinrich Wilhelm Waldeyer wrote a highly influential review of the neuron doctrine in which he introduced the term neuron to describe the anatomical and physiological unit of the nervous system.[45][46]
The silver impregnation stains are a useful method for neuroanatomical investigations because, for reasons unknown, it stains only a small percentage of cells in a tissue, exposing the complete micro structure of individual neurons without much overlap from other cells.[47]
Neuron doctrine[edit]

The neuron doctrine is the now fundamental idea that neurons are the basic structural and functional units of the nervous system. The theory was put forward by Santiago Ramón y Cajal in the late 19th century. It held that neurons are discrete cells (not connected in a meshwork), acting as metabolically distinct units.
Later discoveries yielded refinements to the doctrine. For example, glial cells, which are non-neuronal, play an essential role in information processing.[48] Also, electrical synapses are more common than previously thought,[49] comprising direct, cytoplasmic connections between neurons. In fact, neurons can form even tighter couplings: the squid giant axon arises from the fusion of multiple axons.[50]
Ramón y Cajal also postulated the Law of Dynamic Polarization, which states that a neuron receives signals at its dendrites and cell body and transmits them, as action potentials, along the axon in one direction: away from the cell body.[51] The Law of Dynamic Polarization has important exceptions; dendrites can serve as synaptic output sites of neurons[52] and axons can receive synaptic inputs.[53]
Compartmental modelling of neurons[edit]
Although neurons are often described of as «fundamental units» of the brain, they perform internal computations. Neurons integrate input within dendrites, and this complexity is lost in models that assume neurons to be a fundamental unit. Dendritic branches can be modeled as spatial compartments, whose activity is related due to passive membrane properties, but may also be different depending on input from synapses. Compartmental modelling of dendrites is especially helpful for understanding the behavior of neurons that are too small to record with electrodes, as is the case for Drosophila melanogaster.[54]
Neurons in the brain[edit]
The number of neurons in the brain varies dramatically from species to species.[55] In a human, there are an estimated 10–20 billion neurons in the cerebral cortex and 55–70 billion neurons in the cerebellum.[56] By contrast, the nematode worm Caenorhabditis elegans has just 302 neurons, making it an ideal model organism as scientists have been able to map all of its neurons. The fruit fly Drosophila melanogaster, a common subject in biological experiments, has around 100,000 neurons and exhibits many complex behaviors. Many properties of neurons, from the type of neurotransmitters used to ion channel composition, are maintained across species, allowing scientists to study processes occurring in more complex organisms in much simpler experimental systems.
Neurological disorders[edit]
Charcot–Marie–Tooth disease (CMT) is a heterogeneous inherited disorder of nerves (neuropathy) that is characterized by loss of muscle tissue and touch sensation, predominantly in the feet and legs extending to the hands and arms in advanced stages. Presently incurable, this disease is one of the most common inherited neurological disorders, affecting 36 in 100,000 people.[57]
Alzheimer’s disease (AD), also known simply as Alzheimer’s, is a neurodegenerative disease characterized by progressive cognitive deterioration, together with declining activities of daily living and neuropsychiatric symptoms or behavioral changes.[58] The most striking early symptom is loss of short-term memory (amnesia), which usually manifests as minor forgetfulness that becomes steadily more pronounced with illness progression, with relative preservation of older memories. As the disorder progresses, cognitive (intellectual) impairment extends to the domains of language (aphasia), skilled movements (apraxia), and recognition (agnosia), and functions such as decision-making and planning become impaired.[59][60]
Parkinson’s disease (PD), also known as Parkinsons, is a degenerative disorder of the central nervous system that often impairs motor skills and speech.[61] Parkinson’s disease belongs to a group of conditions called movement disorders.[62] It is characterized by muscle rigidity, tremor, a slowing of physical movement (bradykinesia), and in extreme cases, a loss of physical movement (akinesia). The primary symptoms are the results of decreased stimulation of the motor cortex by the basal ganglia, normally caused by the insufficient formation and action of dopamine, which is produced in the dopaminergic neurons of the brain. Secondary symptoms may include high level cognitive dysfunction and subtle language problems. PD is both chronic and progressive.
Myasthenia gravis is a neuromuscular disease leading to fluctuating muscle weakness and fatigability during simple activities. Weakness is typically caused by circulating antibodies that block acetylcholine receptors at the post-synaptic neuromuscular junction, inhibiting the stimulative effect of the neurotransmitter acetylcholine. Myasthenia is treated with immunosuppressants, cholinesterase inhibitors and, in selected cases, thymectomy.
Demyelination[edit]

Demyelination is the act of demyelinating, or the loss of the myelin sheath insulating the nerves. When myelin degrades, conduction of signals along the nerve can be impaired or lost, and the nerve eventually withers. This leads to certain neurodegenerative disorders like multiple sclerosis and chronic inflammatory demyelinating polyneuropathy.
Axonal degeneration[edit]
Although most injury responses include a calcium influx signaling to promote resealing of severed parts, axonal injuries initially lead to acute axonal degeneration, which is the rapid separation of the proximal and distal ends, occurring within 30 minutes of injury.[63] Degeneration follows with swelling of the axolemma, and eventually leads to bead-like formation. Granular disintegration of the axonal cytoskeleton and inner organelles occurs after axolemma degradation. Early changes include accumulation of mitochondria in the paranodal regions at the site of injury. Endoplasmic reticulum degrades and mitochondria swell up and eventually disintegrate. The disintegration is dependent on ubiquitin and calpain proteases (caused by the influx of calcium ion), suggesting that axonal degeneration is an active process that produces complete fragmentation. The process takes about roughly 24 hours in the PNS and longer in the CNS. The signaling pathways leading to axolemma degeneration are unknown.
Neurogenesis[edit]
Neurons are born through the process of neurogenesis, in which neural stem cells divide to produce differentiated neurons. Once fully differentiated neurons are formed, they are no longer capable of undergoing mitosis. Neurogenesis primarily occurs in the embryo of most organisms.
Adult neurogenesis can occur and studies of the age of human neurons suggest that this process occurs only for a minority of cells, and that the vast majority of neurons in the neocortex forms before birth and persists without replacement. The extent to which adult neurogenesis exists in humans, and its contribution to cognition are controversial, with conflicting reports published in 2018.[64]
The body contains a variety of stem cell types that have the capacity to differentiate into neurons. Researchers found a way to transform human skin cells into nerve cells using transdifferentiation, in which «cells are forced to adopt new identities».[65]
During neurogenesis in the mammalian brain, progenitor and stem cells progress from proliferative divisions to differentiative divisions. This progression leads to the neurons and glia that populate cortical layers. Epigenetic modifications play a key role in regulating gene expression in differentiating neural stem cells, and are critical for cell fate determination in the developing and adult mammalian brain. Epigenetic modifications include DNA cytosine methylation to form 5-methylcytosine and 5-methylcytosine demethylation.[66] These modifications are critical for cell fate determination in the developing and adult mammalian brain. DNA cytosine methylation is catalyzed by DNA methyltransferases (DNMTs). Methylcytosine demethylation is catalyzed in several stages by TET enzymes that carry out oxidative reactions (e.g. 5-methylcytosine to 5-hydroxymethylcytosine) and enzymes of the DNA base excision repair (BER) pathway.[66]
At different stages of mammalian nervous system development two DNA repair processes are employed in the repair of DNA double-strand breaks. These pathways are homologous recombinational repair used in proliferating neural precursor cells, and non-homologous end joining used mainly at later developmental stages[67]
Intercellular communication between developing neurons and microglia is also indispensable for proper neurogenesis and brain development.[68]
Nerve regeneration[edit]
Peripheral axons can regrow if they are severed,[69] but one neuron cannot be functionally replaced by one of another type (Llinás’ law).[18]
See also[edit]
- Artificial neuron
- Bidirectional cell
- Biological neuron model
- Compartmental neuron models
- Connectome
- Dogiel cell
- List of animals by number of neurons
- List of neuroscience databases
- Neuronal galvanotropism
- Neuroplasticity
- Growth cone
- Sholl analysis
References[edit]
- ^ Moore, Keith; Dalley, Arthur (2005). Clinically Oriented Anatomy (5th ed.). LWW. pp. 47. ISBN 0-7817-3639-0.
A bundle of nerve fibers (axons) connecting neighboring or distant nuclei of the CNS is a tract.
- ^ «What are the parts of the nervous system?». Retrieved 2022-07-08.
- ^ Davies, Melissa (2002-04-09). «The Neuron: size comparison». Neuroscience: A journey through the brain. Retrieved 2009-06-20.
- ^ Chudler EH. «Brain Facts and Figures». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ «16.7: Nervous System». Biology LibreTexts. 2021-01-14. Retrieved 2022-02-28.
- ^ Herrup K, Yang Y (May 2007). «Cell cycle regulation in the postmitotic neuron: oxymoron or new biology?». Nature Reviews. Neuroscience. 8 (5): 368–78. doi:10.1038/nrn2124. PMID 17453017. S2CID 12908713.
- ^ Giménez, C. (February 1998). «[Composition and structure of the neuronal membrane: molecular basis of its physiology and pathology]». Revista de Neurologia. 26 (150): 232–239. ISSN 0210-0010. PMID 9563093.
- ^ State Hospitals Bulletin. State Commission in Lunacy. 1897. p. 378.
- ^ «Medical Definition of Neurotubules». www.merriam-webster.com.
- ^ Zecca L, Gallorini M, Schünemann V, Trautwein AX, Gerlach M, Riederer P, Vezzoni P, Tampellini D (March 2001). «Iron, neuromelanin and ferritin content in the substantia nigra of normal subjects at different ages: consequences for iron storage and neurodegenerative processes». Journal of Neurochemistry. 76 (6): 1766–73. doi:10.1046/j.1471-4159.2001.00186.x. PMID 11259494. S2CID 31301135.
- ^ Herrero MT, Hirsch EC, Kastner A, Luquin MR, Javoy-Agid F, Gonzalo LM, Obeso JA, Agid Y (1993). «Neuromelanin accumulation with age in catecholaminergic neurons from Macaca fascicularis brainstem». Developmental Neuroscience. 15 (1): 37–48. doi:10.1159/000111315. PMID 7505739.
- ^ Brunk UT, Terman A (September 2002). «Lipofuscin: mechanisms of age-related accumulation and influence on cell function». Free Radical Biology & Medicine. 33 (5): 611–9. doi:10.1016/s0891-5849(02)00959-0. PMID 12208347.
- ^ Zhao B, Meka DP, Scharrenberg R, König T, Schwanke B, Kobler O, Windhorst S, Kreutz MR, Mikhaylova M, Calderon de Anda F (August 2017). «Microtubules Modulate F-actin Dynamics during Neuronal Polarization». Scientific Reports. 7 (1): 9583. Bibcode:2017NatSR…7.9583Z. doi:10.1038/s41598-017-09832-8. PMC 5575062. PMID 28851982.
- ^ Lee WC, Huang H, Feng G, Sanes JR, Brown EN, So PT, Nedivi E (February 2006). «Dynamic remodeling of dendritic arbors in GABAergic interneurons of adult visual cortex». PLOS Biology. 4 (2): e29. doi:10.1371/journal.pbio.0040029. PMC 1318477. PMID 16366735.
- ^ Al, Martini, Frederic Et (2005). Anatomy and Physiology’ 2007 Ed.2007 Edition. Rex Bookstore, Inc. p. 288. ISBN 978-971-23-4807-5.
- ^ Gerber U (January 2003). «Metabotropic glutamate receptors in vertebrate retina». Documenta Ophthalmologica. Advances in Ophthalmology. 106 (1): 83–7. doi:10.1023/A:1022477203420. PMID 12675489. S2CID 22296630.
- ^ Wilson NR, Runyan CA, Wang FL, Sur M (August 2012). «Division and subtraction by distinct cortical inhibitory networks in vivo». Nature. 488 (7411): 343–8. Bibcode:2012Natur.488..343W. doi:10.1038/nature11347. hdl:1721.1/92709. PMC 3653570. PMID 22878717.
- ^ a b Llinás RR (2014-01-01). «Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective». Frontiers in Cellular Neuroscience. 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ Kolodin YO, Veselovskaia NN, Veselovsky NS, Fedulova SA. Ion conductances related to shaping the repetitive firing in rat retinal ganglion cells. Acta Physiologica Congress. Archived from the original on 2012-10-07. Retrieved 2009-06-20.
- ^ «Ionic conductances underlying excitability in tonically firing retinal ganglion cells of adult rat». Ykolodin.50webs.com. 2008-04-27. Retrieved 2013-02-16.
- ^ Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y (January 2019). «Histamine: neural circuits and new medications». Sleep. 42 (1). doi:10.1093/sleep/zsy183. PMC 6335869. PMID 30239935.
- ^ «Patch-seq technique helps depict the variation of neural cells in the brain». News-medical.net. 3 December 2020. Retrieved 26 August 2021.
{{cite web}}: CS1 maint: url-status (link) - ^ Macpherson, Gordon (2002). Black’s Medical Dictionary (40 ed.). Lanham, MD: Scarecrow Press. pp. 431–434. ISBN 0810849844.
- ^ Ivannikov MV, Macleod GT (June 2013). «Mitochondrial free Ca²⁺ levels and their effects on energy metabolism in Drosophila motor nerve terminals». Biophysical Journal. 104 (11): 2353–61. Bibcode:2013BpJ…104.2353I. doi:10.1016/j.bpj.2013.03.064. PMC 3672877. PMID 23746507.
- ^ Herculano-Houzel S (November 2009). «The human brain in numbers: a linearly scaled-up primate brain». Frontiers in Human Neuroscience. 3: 31. doi:10.3389/neuro.09.031.2009. PMC 2776484. PMID 19915731.
- ^ Drachman DA (June 2005). «Do we have brain to spare?». Neurology. 64 (12): 2004–5. doi:10.1212/01.WNL.0000166914.38327.BB. PMID 15985565. S2CID 38482114.
- ^ Ucar, Hasan; Watanabe, Satoshi; Noguchi, Jun; Morimoto, Yuichi; Iino, Yusuke; Yagishita, Sho; Takahashi, Noriko; Kasai, Haruo (December 2021). «Mechanical actions of dendritic-spine enlargement on presynaptic exocytosis». Nature. 600 (7890): 686–689. Bibcode:2021Natur.600..686U. doi:10.1038/s41586-021-04125-7. ISSN 1476-4687. PMID 34819666. S2CID 244648506.
Lay summary:
«Forceful synapses reveal mechanical interactions in the brain». Nature. 24 November 2021. doi:10.1038/d41586-021-03516-0. Retrieved 21 February 2022. - ^ «Researchers discover new type of cellular communication in the brain». The Scripps Research Institute. Retrieved 12 February 2022.
- ^ Schiapparelli, Lucio M.; Sharma, Pranav; He, Hai-Yan; Li, Jianli; Shah, Sahil H.; McClatchy, Daniel B.; Ma, Yuanhui; Liu, Han-Hsuan; Goldberg, Jeffrey L.; Yates, John R.; Cline, Hollis T. (25 January 2022). «Proteomic screen reveals diverse protein transport between connected neurons in the visual system». Cell Reports. 38 (4): 110287. doi:10.1016/j.celrep.2021.110287. ISSN 2211-1247. PMC 8906846. PMID 35081342.
- ^ Levitan, Irwin B.; Kaczmarek, Leonard K. (2015). «Electrical Signaling in Neurons». The Neuron. Oxford University Press. pp. 41–62. doi:10.1093/med/9780199773893.003.0003. ISBN 978-0-19-977389-3.
- ^ O’Leary, Olivia F.; Ogbonnaya, Ebere S.; Felice, Daniela; Levone, Brunno R.; C. Conroy, Lorraine; Fitzgerald, Patrick; Bravo, Javier A.; Forsythe, Paul; Bienenstock, John; Dinan, Timothy G.; Cryan, John F. (1 February 2018). «The vagus nerve modulates BDNF expression and neurogenesis in the hippocampus». European Neuropsychopharmacology. 28 (2): 307–316. doi:10.1016/j.euroneuro.2017.12.004. ISSN 0924-977X. PMID 29426666. S2CID 46819013.
- ^ Cserép C, Pósfai B, Lénárt N, Fekete R, László ZI, Lele Z (January 2020). «Microglia monitor and protect neuronal function through specialized somatic purinergic junctions». Science. 367 (6477): 528–537. Bibcode:2020Sci…367..528C. doi:10.1126/science.aax6752. PMID 31831638. S2CID 209343260.
- ^ Chudler EH. «Milestones in Neuroscience Research». Neuroscience for Kids. Retrieved 2009-06-20.
- ^ Patlak J, Gibbons R (2000-11-01). «Electrical Activity of Nerves». Action Potentials in Nerve Cells. Archived from the original on August 27, 2009. Retrieved 2009-06-20.
- ^ Harris-Warrick, RM (October 2011). «Neuromodulation and flexibility in Central Pattern Generator networks». Current Opinion in Neurobiology. 21 (5): 685–92. doi:10.1016/j.conb.2011.05.011. PMC 3171584. PMID 21646013.
- ^ Brown EN, Kass RE, Mitra PP (May 2004). «Multiple neural spike train data analysis: state-of-the-art and future challenges». Nature Neuroscience. 7 (5): 456–61. doi:10.1038/nn1228. PMID 15114358. S2CID 562815.
- ^ Thorpe SJ (1990). «Spike arrival times: A highly efficient coding scheme for neural networks» (PDF). In Eckmiller R, Hartmann G, Hauske G (eds.). Parallel processing in neural systems and computers. North-Holland. pp. 91–94. ISBN 9780444883902. Archived from the original (PDF) on 2012-02-15.
- ^ a b Kalat, James W (2016). Biological psychology (12 ed.). Australia. ISBN 9781305105409. OCLC 898154491.
- ^ Eckert R, Randall D (1983). Animal physiology: mechanisms and adaptations. San Francisco: W.H. Freeman. p. 239. ISBN 978-0-7167-1423-1.
- ^ a b Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
Ramon y Cajal’s first paper on the Golgi stain was on the bird cerebellum, and it appeared in the Revista in 1888. He acknowledged that he found the nerve fibers to be very intricate, but stated that he could find no evidence for either axons or dendrites undergoing anastomosis and forming nets. He called each nervous element ‘an absolutely autonomous canton.’
- ^ a b Oxford English Dictionary, 3rd edition, 2003, s.v.
- ^ Mehta AR, Mehta PR, Anderson SP, MacKinnon BL, Compston A (January 2020). «Grey Matter Etymology and the neuron(e)». Brain. 143 (1): 374–379. doi:10.1093/brain/awz367. PMC 6935745. PMID 31844876.
- ^ «Google Books Ngram Viewer». books.google.com. Retrieved 19 December 2020.
- ^ a b c d López-Muñoz F, Boya J, Alamo C (October 2006). «Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal». Brain Research Bulletin. 70 (4–6): 391–405. doi:10.1016/j.brainresbull.2006.07.010. PMID 17027775. S2CID 11273256.
- ^ Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. p. 47. ISBN 9780195146943. OCLC 27151391.
… a man who would write a highly influential review of the evidence in favor of the neuron doctrine two years later. In his paper, Waldeyer (1891), … , wrote that nerve cells terminate freely with end arborizations and that the ‘neuron’ is the anatomical and physiological unit of the nervous system. The word ‘neuron’ was born this way.
- ^ «Whonamedit — dictionary of medical eponyms». www.whonamedit.com.
Today, Wilhelm von Waldeyer-Hartz is remembered as the founder of the neurone theory, coining the term «neurone» to describe the cellular function unit of the nervous system and enunciating and clarifying that concept in 1891.
- ^ Grant G (October 2007). «How the 1906 Nobel Prize in Physiology or Medicine was shared between Golgi and Cajal». Brain Research Reviews. 55 (2): 490–8. doi:10.1016/j.brainresrev.2006.11.004. PMID 17306375. S2CID 24331507.
- ^ Witcher MR, Kirov SA, Harris KM (January 2007). «Plasticity of perisynaptic astroglia during synaptogenesis in the mature rat hippocampus». Glia. 55 (1): 13–23. CiteSeerX 10.1.1.598.7002. doi:10.1002/glia.20415. PMID 17001633. S2CID 10664003.
- ^ Connors BW, Long MA (2004). «Electrical synapses in the mammalian brain». Annual Review of Neuroscience. 27 (1): 393–418. doi:10.1146/annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Guillery RW (June 2005). «Observations of synaptic structures: origins of the neuron doctrine and its current status». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1458): 1281–307. doi:10.1098/rstb.2003.1459. PMC 1569502. PMID 16147523.
- ^ Sabbatini RM (April–July 2003). «Neurons and Synapses: The History of Its Discovery». Brain & Mind Magazine: 17.
- ^ Djurisic M, Antic S, Chen WR, Zecevic D (July 2004). «Voltage imaging from dendrites of mitral cells: EPSP attenuation and spike trigger zones». The Journal of Neuroscience. 24 (30): 6703–14. doi:10.1523/JNEUROSCI.0307-04.2004. hdl:1912/2958. PMC 6729725. PMID 15282273.
- ^ Cochilla AJ, Alford S (March 1997). «Glutamate receptor-mediated synaptic excitation in axons of the lamprey». The Journal of Physiology. 499 (Pt 2): 443–57. doi:10.1113/jphysiol.1997.sp021940. PMC 1159318. PMID 9080373.
- ^ Gouwens NW, Wilson RI (2009). «Signal propagation in Drosophila central neurons». Journal of Neuroscience. 29 (19): 6239–6249. doi:10.1523/jneurosci.0764-09.2009. PMC 2709801. PMID 19439602.
- ^ Williams RW, Herrup K (1988). «The control of neuron number». Annual Review of Neuroscience. 11 (1): 423–53. doi:10.1146/annurev.ne.11.030188.002231. PMID 3284447.
- ^ von Bartheld CS, Bahney J, Herculano-Houzel S (December 2016). «The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting». The Journal of Comparative Neurology. 524 (18): 3865–3895. doi:10.1002/cne.24040. PMC 5063692. PMID 27187682.
- ^ Krajewski KM, Lewis RA, Fuerst DR, Turansky C, Hinderer SR, Garbern J, Kamholz J, Shy ME (July 2000). «Neurological dysfunction and axonal degeneration in Charcot-Marie-Tooth disease type 1A». Brain. 123 (7): 1516–27. doi:10.1093/brain/123.7.1516. PMID 10869062.
- ^ «About Alzheimer’s Disease: Symptoms». National Institute on Aging. Archived from the original on 15 January 2012. Retrieved 28 December 2011.
- ^ Burns A, Iliffe S (February 2009). «Alzheimer’s disease». BMJ. 338: b158. doi:10.1136/bmj.b158. PMID 19196745. S2CID 8570146.
- ^ Querfurth HW, LaFerla FM (January 2010). «Alzheimer’s disease». The New England Journal of Medicine. 362 (4): 329–44. doi:10.1056/NEJMra0909142. PMID 20107219. S2CID 205115756.
- ^ «Parkinson’s Disease Information Page». NINDS. 30 June 2016. Archived from the original on 4 January 2017. Retrieved 18 July 2016.
- ^ «Movement Disorders». The International Neuromodulation Society.
- ^ Kerschensteiner M, Schwab ME, Lichtman JW, Misgeld T (May 2005). «In vivo imaging of axonal degeneration and regeneration in the injured spinal cord». Nature Medicine. 11 (5): 572–7. doi:10.1038/nm1229. PMID 15821747. S2CID 25287010.
- ^ Kempermann G, Gage FH, Aigner L, Song H, Curtis MA, Thuret S, Kuhn HG, Jessberger S, Frankland PW, Cameron HA, Gould E, Hen R, Abrous DN, Toni N, Schinder AF, Zhao X, Lucassen PJ, Frisén J (July 2018). «Human Adult Neurogenesis: Evidence and Remaining Questions». Cell Stem Cell. 23 (1): 25–30. doi:10.1016/j.stem.2018.04.004. PMC 6035081. PMID 29681514.
- ^ Callaway, Ewen (26 May 2011). «How to make a human neuron». Nature. doi:10.1038/news.2011.328.
By transforming cells from human skin into working nerve cells, researchers may have come up with a model for nervous-system diseases and perhaps even regenerative therapies based on cell transplants. The achievement, reported online today in Nature, is the latest in a fast-moving field called transdifferentiation, in which cells are forced to adopt new identities. In the past year, researchers have converted connective tissue cells found in skin into heart cells, blood cells, and liver cells.
- ^ a b Wang Z, Tang B, He Y, Jin P (March 2016). «DNA methylation dynamics in neurogenesis». Epigenomics. 8 (3): 401–14. doi:10.2217/epi.15.119. PMC 4864063. PMID 26950681.
- ^ Orii KE, Lee Y, Kondo N, McKinnon PJ (June 2006). «Selective utilization of nonhomologous end-joining and homologous recombination DNA repair pathways during nervous system development». Proceedings of the National Academy of Sciences of the United States of America. 103 (26): 10017–22. Bibcode:2006PNAS..10310017O. doi:10.1073/pnas.0602436103. PMC 1502498. PMID 16777961.
- ^ Cserép, Csaba; Schwarcz, Anett D.; Pósfai, Balázs; László, Zsófia I.; Kellermayer, Anna; Környei, Zsuzsanna; Kisfali, Máté; Nyerges, Miklós; Lele, Zsolt; Katona, István (September 2022). «Microglial control of neuronal development via somatic purinergic junctions». Cell Reports. 40 (12): 111369. doi:10.1016/j.celrep.2022.111369. PMC 9513806. PMID 36130488. S2CID 252416407.
- ^ Yiu G, He Z (August 2006). «Glial inhibition of CNS axon regeneration». Nature Reviews. Neuroscience. 7 (8): 617–27. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
Further reading[edit]
- Bullock TH, Bennett MV, Johnston D, Josephson R, Marder E, Fields RD (November 2005). «Neuroscience. The neuron doctrine, redux». Science. 310 (5749): 791–3. doi:10.1126/science.1114394. PMID 16272104. S2CID 170670241.
- Kandel ER, Schwartz JH, Jessell TM (2000). Principles of Neural Science (4th ed.). New York: McGraw-Hill. ISBN 0-8385-7701-6.
- Peters A, Palay SL, Webster HS (1991). The Fine Structure of the Nervous System (3rd ed.). New York: Oxford University Press. ISBN 0-19-506571-9.
- Ramón y Cajal S (1933). Histology (10th ed.). Baltimore: Wood.
- Roberts A, Bush BM (1981). Neurones without Impulses. Cambridge: Cambridge University Press. ISBN 0-521-29935-7.
- Snell RS (2010). Clinical Neuroanatomy. Lippincott Williams & Wilkins. ISBN 978-0-7817-9427-5.
External links[edit]
- Neurobiology at Curlie
- IBRO (International Brain Research Organization). Fostering neuroscience research especially in less well-funded countries.
- NeuronBank an online neuromics tool for cataloging neuronal types and synaptic connectivity.
- High Resolution Neuroanatomical Images of Primate and Non-Primate Brains.
- The Department of Neuroscience at Wikiversity, which presently offers two courses: Fundamentals of Neuroscience and Comparative Neuroscience.
- NIF Search – Neuron via the Neuroscience Information Framework
- Cell Centered Database – Neuron
- Complete list of neuron types according to the Petilla convention, at NeuroLex.
- NeuroMorpho.Org an online database of digital reconstructions of neuronal morphology.
- Immunohistochemistry Image Gallery: Neuron
- Khan Academy: Anatomy of a neuron
- Neuron images
Нервная система выполняет ряд важных функций:
- обеспечивает связь организма с окружающим миром;
- управляет работой всех органов;
- координирует функционирование всех систем органов, обеспечивая их согласованную работу.
Нервная ткань
Нервная ткань отличается от других тканей нашего организма тем, что обладает особыми свойствами — возбудимостью и проводимостью. Эти свойства нервной ткани обусловлены особенностями её строения.
В состав нервной ткани входят клетки двух видов. Основные функции выполняют нейроны, а клетки-спутники (клетки нейроглии) служат опорой и обеспечивают обмен веществ.

Рис. (1). Нервная ткань
Функции нейронов: генерирование и передача нервных импульсов; обработка и хранение поступающей информации.
Нервный импульс — это волна возбуждения (биоэлектрическая волна), распространяющаяся по нервным клеткам.
Нейрон — основная клетка нервной ткани. Он имеет тело и отростки двух типов. В теле нейрона располагается ядро и органоиды, а по отросткам передаются нервные импульсы.
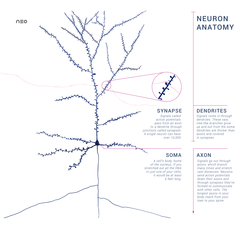
Дендриты — это отростки, по которым нервные импульсы передаются к телу нейрона. Эти отростки сильно ветвятся. У нейрона может быть несколько дендритов.
Аксон — это отросток, по которому импульсы передаются от тела клетки. Аксон обычно ветвится только на конце. У каждого нейрона всего один аксон.

Рис. (2). Строение нейрона
Аксоны часто окружены оболочкой из жироподобного вещества миелина. Это вещество имеет белый цвет. Скопления миелинизированных аксонов образуют белое вещество головного и спинного мозга. Тела нервных клеток и дендриты не покрыты миелином. Они серого цвета, а их группы составляют серое вещество центральной нервной системы.
Передача нервных импульсов с одной клетки на другую происходит в синапсах.
Синапс — это место контакта между двумя нейронами или между нейроном и клеткой рабочего органа.
Главными элементами синапса являются мембраны двух клеток (пресинаптическая и постсинаптическая мембраны) и пространство между ними (синаптическая щель).

Рис. (3). Строение синапса
В аксоне пресинаптического нейрона вырабатывается медиатор — особое вещество, с помощью которого происходит передача нервного импульса.
Под действием нервного импульса медиатор выделяется в синаптическую щель. Рецепторы постсинаптической мембраны реагируют на его появление и генерируют возникновение нервного импульса в следующем нейроне. Так в синапсе происходит химическая передача возбуждения с одной клетки на другую.
Нейроны различаются по своему строению и выполняемым функциям.

Рис. (4). Виды нейронов
По выполняемым функциям выделяют три типа нейронов.
Чувствительные (сенсорные) нейроны проводят информацию от органов в мозг. Тела таких нейронов находятся в нервных узлах вне центральной нервной системы.
Другая группа нейронов передаёт информацию от головного и спинного мозга к органам. Это двигательные (моторные) нейроны. Их тела находятся в сером веществе центральной нервной системы, а аксоны находятся за пределами ЦНС.
Третий вид нейронов осуществляет связь между чувствительными и двигательными нейронами. Это вставочные нейроны, они находятся в головном и спинном мозге.
Скопление нейронов в головном или спинном мозге называют ядром.

Рис. (5). Типы нейронов и синапсы
Связь между органами и центральной нервной системой осуществляется через нервы.
Нерв — это орган, в состав которого входят пучки нервных волокон, покрытые соединительнотканной оболочкой.

Рис. (6). Нерв
Нервы выполняют проводниковую функцию. Они связывают головной и спинной мозг с кожей, органами чувств и с внутренними органами.
Нервы бывают чувствительные, двигательные и смешанные.
Чувствительные нервы проводят нервные импульсы от рецепторов в мозг. В их состав входят дендриты чувствительных нейронов.
Двигательные нервы состоят из аксонов двигательных нейронов. Их функция — проведение импульсов от мозга к рабочим органам.
Смешанные нервы образованы чувствительными и двигательными волокнами и способные проводить импульсы как к ЦНС, так и от ЦНС.
Нервные сплетения представлены сетчатыми скоплениями нервных волокон разных нервов, связывающих ЦНС с внутренними органами, скелетными мышцами и кожей.
Наиболее известное солнечное сплетение находится в брюшной полости.
Источники:
Рис. 1. Нервная ткань https://image.shutterstock.com/image-photo/mammalian-nervous-tissue-under-microscope-600w-74170234.jpg
Рис. 2. Строение нейрона https://image.shutterstock.com/image-vector/education-chart-biology-nerve-cell-600w-661087429.jpg
Рис. 3. Строение синапса https://image.shutterstock.com/image-illustration/gap-between-two-nerve-cells-600w-1284912691.jpg
Рис. 4. Виды нейронов https://image.shutterstock.com/image-illustration/different-kinds-neurons-scheme-structure-600w-138356969.jpg
Рис. 5. Типы нейронов и синапсы © ЯКласс
Рис. 6. Нерв https://image.shutterstock.com/image-illustration/nerve-structure-anatomy-600w-1041115012.jpg
В теле человека бессчетное количество клеток, каждая из которых имеет собственную функцию. Среди них самые загадочные – нейроны, отвечающие за любое совершаемое нами действие. Попробуем разобраться как работают нейроны и в чем их предназначение.
Что такое нейрон (нейронные связи)
В переводе с греческого нейрон, или как его еще называют неврон, означает «волокно», «нерв». Нейрон – это специфическая структура в нашем организме, которая отвечает за передачу внутри него любой информации, в быту называемая нервной клеткой.
Нейроны работают при помощи электрических сигналов и способствуют обработке мозгом поступающей информации для дальнейшей координации производимых телом действий.
Эти клетки являются составляющей частью нервной системы человека, предназначение которой состоит в том, чтобы собрать все сигналы, поступающие из вне или от собственного организма и принять решение о необходимости того или иного действия. Именно нейроны помогают справиться с такой задачей.
Каждый из нейронов имеет связь с огромным количеством таких же клеток, создаётся своеобразная «паутина», которая называется нейронной сетью. Посредством данной связи в организме передаются электрические и химические импульсы, приводящие всю нервную систему в состояние покоя либо, наоборот, возбуждения.
К примеру, человек столкнулся с неким значимым событием. Возникает электрохимический толчок (импульс) нейронов, приводящий к возбуждению неровной системы. У человека начинает чаще биться сердце, потеют руки или возникают другие физиологические реакции.
Мы рождаемся с заданным количеством нейронов, но связи между ними еще не сформированы. Нейронная сеть строится постепенно в результате поступающих из вне импульсов. Новые толчки формируют новые нейронные пути, именно по ним в течение жизни побежит аналогичная информация. Мозг воспринимает индивидуальный опыт каждого человека и реагирует на него. К примеру, ребенок, схватился за горячий утюг и отдернул руку. Так у него появилась новая нейронная связь.
Стабильная нейронная сеть выстраивается у ребенка уже к двум годам. Удивительно, но уже с этого возраста те клетки, которые не используются, начинают ослабевать. Но это никак не мешает развитию интеллекта. Наоборот, ребенок познает мир через уже устоявшиеся нейронные связи, а не анализирует бесцельно все вокруг.
Даже у такого малыша есть практический опыт, позволяющий отсекать ненужные действия и стремиться к полезным. Поэтому, например, так сложно отучить ребенка от груди — у него сформировалась крепкая нейронная связь между приложением к материнскому молоку и удовольствию, безопасности, спокойствию.
Познание нового опыта на протяжении всей жизни приводит к отмиранию ненужных нейронных связей и формированию новых и полезных. Этот процесс оптимизирует головной мозг наиболее эффективным для нас образом. Например, люди, проживающие в жарких странах, учатся жить в определенном климате, а северянам нужен совсем другой опыт для выживания.
Сколько нейронов в мозге
Нервные клетки в составе головного мозга занимают порядка 10 процентов, остальные 90 процентов это астроциты и глиальные клетки, но их задача заключается лишь в обслуживании нейронов.
Подсчитать «вручную» численность клеток в головном мозге также сложно, как узнать количество звезд на небе.
Тем не менее ученые придумали сразу несколько способов для определения количества нейронов у человека:
- Рассчитывается число нервных клеток на небольшой части мозга, а затем, количество умножается пропорционально полному объему. Исследователи исходят из постулата о том, что нейроны равномерно распределены в нашем мозге.
- Происходит растворение всех мозговых клеток. В результате получается жидкость, в составе которой можно увидеть клеточные ядра. Их можно посчитать. При этом служебные клетки, о которых мы сказали выше, не учитываются.
В результате описанных экспериментов установлено, что число нейронов в головном мозге человека — 85 миллиардов единиц. Ранее, на протяжении многих веков считалось, что нервных клеток больше, порядка 100 миллиардов.
Строение нейрона
На рисунке приведено строение нейрона. Он состоит из основного тела и ядра. От клеточного тела идет ответвление многочисленных волокон, которые именуются дендритами.

Мощные и длинные дендриты называются аксонами, которые в действительности намного длиннее, чем на картинке. Их протяженность варьируется от нескольких миллиметров до более метра.
Аксоны играют ведущую роль в передаче информации между нейронами и обеспечивают работу всей нервной системы.
Место соединения дендрита (аксона) с другим нейроном называется синапсом. Дендриты при наличии раздражителей могут разрастись настолько сильно, что станут улавливать импульсы от других клеток, что приводит к образованию новых синаптических связей.
Синаптические связи играют существенную роль в формировании личности человека. Так, личность с устоявшимся позитивным опытом будет смотреть на жизнь с любовью и надеждой, человек, у которого нейронные связи с негативным зарядом, станет со временем пессимистом.
Виды нейронов и нейронных связей
Нейроны можно обнаружить в различных органах человека, а не исключительно в головном мозге. Большое их количество расположено в рецепторах (глаза, уши, язык, пальцы рук – органы чувств). Совокупность нервных клеток, которые пронизывают наш организм составляет основу периферической нервной системы. Выделим основные виды нейронов.
| Вид нейронной клетки | За что отвечает |
|---|---|
| Аффекторные | Являются переносчиками информации от органов чувств в головной мозг. У этого вида нейронов самые длинные аксоны. Импульс из вне поступает по аксонам строго в определенный участок головного мозга, звук — в слуховой «отсек», запах – в «обонятельный» и т.д. |
| Промежуточные | Промежуточные нервные клетки обрабатывают сведения, поступившие от аффекторных нейронов и передают ее периферическим органам и мышцам. |
| Эффекторные | На заключительном этапе в дело вступают эфференты, которые доводят команду промежуточных нейронов до мышц и других органов тела. |
Слаженная работа нейронов трех типов выглядит так: человек «слышит» запах шашлыка, нейрон передает информацию в соответствующий раздел мозга, мозг передает сигнал желудку, который выделяет желудочный сок, человек принимает решение «хочу есть» и бежит покупать шашлык. Упрощенно так это действует.
Самыми загадочными являются промежуточные нейроны. С одной стороны, их работа обуславливает наличие рефлекса: дотронулся до электричества – отдернул руку, полетела пыль –зажмурился. Однако, пока не объяснимо как обмен между волокнами рождает идеи, образы, мысли?
Единственное, что установили ученые, это тот факт, что любой вид мыслительной деятельности (чтение книг, рисование, решение математических задач) сопровождается особой активностью (вспышкой) нервных клеток определенного участка головного мозга.
Есть особая разновидность нейронов, которые именуются зеркальными. Их особенность заключается в том, что они не только приходят в возбуждение от внешних сигналов, но и начинают «шевелиться», наблюдая за действиями своих собратьев – других нейронов.
Функции нейронов
Без нейронов невозможна работа организма человека. Мы увидели, что эти наноклетки отвечают буквально за каждое наше движение, любой поступок. Выполняемые ими функции до настоящего времени в полной мере не изучены и не определены.
Существует несколько классификаций функций нейронов. Мы остановимся на общепринятой в научном мире.
Функция распространения информации
Данная функция:
- является основной;
- изучена лучше остальных.
Суть ее в том, что нейронами обрабатываются и переносятся в головной мозг все импульсы, которые поступают из окружающего мира или собственного тела. Далее происходит их обработка, подобно тому, как работает поисковик в браузере.
По результатам сканирования сведений из вне, головной мозг в форме обратной связи передает обработанную информацию к органам чувств или мышцам.
Мы не подозреваем, что в нашем теле происходит ежесекундная доставка и переработка информации, не только в голове и на уровне периферической нервной системы.
До настоящего времени создать искусственный интеллект, который бы приблизился к работе нейронных сетей человека, не удалось. У каждого из 85 миллиардов нейронов имеется, как минимум, 10 тысяч обусловленных опытом связей, и все они работают на передачу и обработку информации.
Функция аккумуляции знаний (сохранения опыта)
Человек обладает памятью, возможностью понимать суть вещей, явлений и действий, которые он единожды или многократно повторял. За формирование памяти отвечают именно нейронные клетки, точнее нейротрансмиттеры, связующие звенья между соседними нейронами.
Таким образом, за память отвечает не какая-то отдельная часть мозга, а маленькие белковые мостики между клетками. Человек может потерять память, когда произошло крушение этих нервных связей.
Функция интеграции
Данная функция позволяет взаимодействовать между собой отдельным долям головного мозга. Как мы уже сказали, сигналы от разных органов чувств поступают в разные отделы мозга.
Нейроны посредством «вспышек» активности передают и принимают импульсы в разных частях мозга. Так происходит процесс появления мыслей, эмоций и чувств. Чем больше таких разноплановых связей, тем эффективнее человек мыслит. Если человек способен к размышлениям и аналитике в определенном направлении, то он будет хорошо соображать и в другом вопросе.
Функция производства белков
Нейроны – настолько полезные клетки, что не ограничиваются только передаточными функциями. Нервные клетки вырабатывают необходимые для жизни человека белки. Опять же ключевую роль в производстве белков имеют нейротрансмиттеры, которые отвечают за память.
Всего в невронах индуцируется порядка 80 белков, вот основные из них, влияющие на самочувствие человека:
- Серотонин – вещество, вызывающее радость и удовольствие.
- Допамин – ведущий источник бодрости и счастья для человека. Активизирует физическую активность, помогает проснуться, переизбыток может привести к состоянию эйфории.
- Норадреналин – это «плохой» гормон, вызывающий приступы ярости и гнева. Наряду с кортизолом его называют гормоном стресса.
- Глутамат – вещество, отвечающие за хранение памяти.
Прекращение выработки белков или их выпуск в недостаточном количестве способны привести к тяжелым заболеваниям.
Восстанавливаются ли нервные клетки
При нормальном состоянии организма нейроны могут жить и функционировать очень долго. К сожалению, случается так, что они начинают массово погибать. Причин разрушения нервных волокон может быть много, но до конца механизм их деструкции не изучен.
Установлено, что нервные клетки погибают из-за гипоксии (кислородное голодание). Нейронные сети рушатся при отдельных травмах головного мозга, человек теряет память или утрачивает способность к хранению информации. В этом случае сами нейроны сохранены, но теряется их передаточная функция.
Отсутствие допамина ведет к развитию болезни Паркинсона, а его переизбыток является причиной шизофрении. Почему прекращается выработка белка не известно, спусковой механизм не выявлен.
Гибель нервных клеток происходит при алкоголизации личности. Алкоголик со временем может совершенно деградировать и утратить вкус к жизни.
Формирование нервных клеток происходит при рождении. Долгое время ученые полагали, что со временем нейроны отмирают. Поэтому с возрастом человек утрачивает способность накапливать информацию, хуже соображает. Нарушение функции по выработке допамина и серотонина связывается с наличием практически у всех пожилых людей депрессивных состояний.
Гибель нейронов, действительно неизбежна, в год исчезает примерно 1 процент от их количества. Но есть и хорошие новости. Последние исследования показали, что в коре головного мозга есть особенный участок, именуемый гипокаммом. Именно в нем генерируются новые чистые нейроны. Подсчитано примерное количество генерируемых ежедневно нервных клеток – 1400.
В науке обозначилось новое понятие «нейропластичность», обозначающее возможность мозга регенерироваться и перестраиваться. Но есть одна тонкость: новые нейроны еще не имеют никакого опыта и наработанных связей. Поэтому с возрастом или после заболевания мозг нужно тренировать, как и все иные мышцы тела: получать новые знания, анализировать происходящие события и явления.
Подобно тому, как мы усиливаем бицепс при помощи гантели, активизировать процесс включения новых нервных клеток можно следующими способами:
- изучение новых сфер знаний, которые ранее были не нужны или не интересны. К примеру, математику можно начать изучать живопись, а юристу – основы физики.
- через постановку сложных задач и поиск их решения;
- составлением планов деятельности, которые включают в себя множество исходных данных.
Механизм возрождения прост. У нас имеются совершенно не задействованные новые клетки, которые нужно заставить работать, а сделать это можно лишь путем постановки новых задач и изучения неизвестных предметных сфер.
Вся информация взята из открытых источников.
Если вы считаете, что ваши авторские права нарушены, пожалуйста,
напишите в чате на этом сайте, приложив скан документа подтверждающего ваше право.
Мы убедимся в этом и сразу снимем публикацию.
Как работает наш мозг или как смоделировать душу?
Время на прочтение
11 мин
Количество просмотров 50K

Здравствуй, Geektimes! В ранее опубликованной статье, была представлена модель нервной системы, опишу теорию и принципы, которые легли в её основу.
Теория основана на анализе имеющейся информации о биологическом нейроне и нервной системе из современной нейробиологии и физиологии мозга.
Сначала приведу краткую информацию об объекте моделирования, вся информация изложена далее, учтена и использована в модели.
НЕЙРОН

Нейрон является основным функциональным элементом нервной системы, он состоит из тела нервной клетки и её отростков. Существуют два вида отростков: аксоны и дендриты. Аксон – длинный покрытый миелиновой оболочкой отросток, предназначенный для передачи нервного импульса на далекие расстояния. Дендрит – короткий, ветвящийся отросток, благодаря которым происходит взаимосвязь с множеством соседних клеток.
ТРИ ТИПА НЕЙРОНОВ
Нейроны могут сильно отличаться по форме, размерам и конфигурации, не смотря на это, отмечается принципиальное сходство нервной ткани в различных участках нервной системе, отсутствуют и серьезные эволюционные различия. Нервная клетка моллюска Аплизии может выделять такие же нейромедиаторы и белки, что и клетка человека.
В зависимости от конфигурации выделяют три типа нейронов:

а) рецепторные, центростремительные, или афферентные нейроны, данные нейроны имеют центростремительный аксон, на конце которого имеются рецепторы, рецепторные или афферентные окончания. Эти нейроны можно определить, как элементы, передающие внешние сигналы в систему.
б) интернейроны (вставочные, контактные, или промежуточные) нейроны, не имеющие длинных отростков, но имеющие только дендриты. Таких нейронов в человеческом мозгу больше чем остальных. Данный вид нейронов является основным элементом рефлекторной дуги.
в) моторные, центробежные, или эфферентные, они имеют центростремительный аксон, который имеет эфферентные окончания передающий возбуждение мышечным или железистым клеткам. Эфферентные нейроны служат для передачи сигналов из нервной среды во внешнюю среду.
Обычно в статьях по искусственным нейронным сетям оговаривается наличие только моторных нейронов (с центробежным аксоном), которые связаны в слои иерархической структуры. Подобное описание применимо к биологической нервной системе, но является своего рода частным случаем, речь идет о структурах, базовых условных рефлексов. Чем выше в эволюционном значении нервная система, тем меньше в ней превалируют структуры типа «слои» или строгая иерархия.
ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ
Передача возбуждения происходит от нейрона к нейрону, через специальные утолщения на концах дендритов, называемых синапсами. По типу передачи синапсы разделяют на два вида: химические и электрические. Электрические синапсы передают нервный импульс непосредственно через место контакта. Таких синапсов в нервных системах очень мало, в моделях не будут учитываться. Химические синапсы передают нервный импульс посредством специального вещества медиатора (нейромедиатора, нейротрансмиттера), данный вид синапса широко распространен и подразумевает вариативность в работе.
Важно отметить, что в биологическом нейроне постоянно происходят изменения, отращиваются новые дендриты и синапсы, возможны миграции нейронов. В местах контактов с другими нейронами образуются новообразования, для передающего нейрона — это синапс, для принимающего — это постсинаптическая мембрана, снабжаемая специальными рецепторами, реагирующими на медиатор, то есть можно говорить, что мембрана нейрона — это приемник, а синапсы на дендритах — это передатчики сигнала.
СИНАПС

При активации синапса он выбрасывает порции медиатора, эти порции могут варьироваться, чем больше выделится медиатора, тем вероятнее, что принимаемая сигнал нервная клетка будет активирована. Медиатор, преодолевая синоптическую щель, попадает на постсинаптическую мембрану, на которой расположены рецепторы, реагирующие на медиатор. Далее медиатор может быть разрушен специальным разрушающим ферментом, либо поглощен обратно синапсом, это происходит для сокращения времени действия медиатора на рецепторы.
Так же помимо побудительного воздействия существуют синапсы, оказывающие тормозящее воздействие на нейрон. Обычно такие синапсы принадлежат определенным нейронам, которые обозначаются, как тормозящие нейроны.
Синапсов связывающих нейрон с одной и той же целевой клеткой, может быть множество. Для упрощения примем, всю совокупность, оказываемого воздействия одним нейроном, на другой целевой нейрон за синапс с определённой силой воздействия. Главной характеристикой синапса будет, является его сила.
СОСТОЯНИЕ ВОЗБУЖДЕНИЯ НЕЙРОНА
В состоянии покоя мембрана нейрона поляризована. Это означает, что по обе стороны мембраны располагаются частицы, несущие противоположные заряды. В состоянии покоя наружная поверхность мембраны заряжена положительно, внутренняя – отрицательно. Основными переносчиками зарядов в организме являются ионы натрия (Na+), калия (K+) и хлора (Cl-).
Разница между зарядами поверхности мембраны и внутри тела клетки составляет мембранный потенциал. Медиатор вызывает нарушения поляризации – деполяризацию. Положительные ионы снаружи мембраны устремляются через открытые каналы в тело клетки, меняя соотношение зарядов между поверхностью мембраны и телом клетки.

Изменение мембранного потенциала при возбуждении нейрона
Характер изменений мембранного потенциала при активации нервной ткани неизменен. Независимо от того кокой силы воздействия оказывается на нейрон, если сила превышает некоторое пороговое значение, ответ будет одинаков.
Забегая вперед, хочу отметить, что в работе нервной системы имеет значение даже следовые потенциалы (см. график выше). Они не появляются, вследствие каких-то гармонических колебаний уравновешивающих заряды, являются строгим проявлением определённой фазы состояния нервной ткани при возбуждении.
ТЕОРИЯ ЭЛЕКТРОМАГНИТНОГО ВЗАИМОДЕЙСТВИЯ
Итак, далее приведу теоретические предположения, которые позволят нам создавать математические модели. Главная идея заключается во взаимодействии между зарядами формирующихся внутри тела клетки, во время её активности, и зарядами с поверхностей мембран других активных клеток. Данные заряды являются разноименными, в связи этим можно предположить, как будут располагаться заряды в теле клетки под воздействием зарядов других активных клеток.

Можно сказать, что нейрон чувствует активность других нейронов на расстоянии, стремится направить распространения возбуждения в направлении других активных участков.
В момент активности нейрона можно рассчитать определённую точку в пространстве, которая определялась бы, как сумма масс зарядов, расположенных на поверхностях других нейронов. Указанную точку назовем точкой паттерна, её месторождение зависит от комбинации фаз активности всех нейронов нервной системы. Паттерном в физиологии нервной системы называется уникальная комбинация активных клеток, то есть можно говорить о влиянии возбуждённых участков мозга на работу отдельного нейрона.
Нужно представлять работу нейрона не просто как вычислителя, а своего рода ретранслятор возбуждения, который выбирает направления распространения возбуждения, таким образом, формируются сложные электрические схемы. Первоначально предполагалось, что нейрон просто избирательно отключает/включает для передачи свои синапсы, в зависимости от предпочитаемого направления возбуждения. Но более детальное изучение природы нейрона, привело к выводам, что нейрон может изменять степень воздействия на целевую клетку через силу своих синапсов, что делает нейрон более гибким и вариативным вычислительным элементом нервной системы.

Какое же направление для передачи возбуждения является предпочтительным? В различных экспериментах связанных с образованием безусловных рефлексов, можно определить, что в нервной системе образуются пути или рефлекторные дуги, которые связывают активируемые участки мозга при формировании безусловных рефлексов, создаются ассоциативные связи. Значит, нейрон должен передавать возбуждения к другим активным участкам мозга, запоминать направление и использовать его в дальнейшем.
Представим вектор начало, которого находится в центре активной клети, а конец направлен в точку паттерна определённую для данного нейрона. Обозначим, как вектор предпочитаемого направления распространения возбуждения (T, trend). В биологическом нейроне вектор Т может проявляться в структуре самой нейроплазмы, возможно, это каналы для движения ионов в теле клетки, или другие изменения в структуре нейрона.
Нейрон обладает свойством памяти, он может запоминать вектор Т, направление этого вектора, может меняться и перезаписываться в зависимости от внешних факторов. Степень с которой вектор Т может подвергается изменениям, называется нейропластичность.
Этот вектор в свою очередь оказывает влияние на работу синапсов нейрона. Для каждого синапса определим вектор S начало, которого находится в центре клетки, а конец направлен в центр целевого нейрона, с которым связан синапс. Теперь степень влияния для каждого синапса можно определить следующим образом: чем меньше угол между вектором T и S, тем больше синапс будет, усиливается; чем меньше угол, тем сильнее синапс будет ослабевать и возможно может прекратить передачу возбуждения. Каждый синапс имеет независимое свойство памяти, он помнит значение своей силы. Указанные значения изменяются при каждой активизации нейрона, под влиянием вектора Т, они либо увеличиваются, либо уменьшаются на определённое значение.
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ

Входные сигналы (x1, x2,…xn) нейрона представляют собой вещественные числа, которые характеризуют силу синапсов нейронов, оказывающих воздействие на нейрон.
Положительное значение входа означает побудительное воздействие, оказываемое на нейрон, а отрицательное значение – тормозящее воздействие.
Для биологического нейрона не имеет значение, откуда поступил возбуждающий его сигнал, результат его активности будет идентичен. Нейрон будет активизирован, когда сумма воздействий на него будет превышать определённое пороговое значение. Поэтому, все сигналы проходят через сумматор (а), а поскольку нейроны и нервная система работают в реальном времени, следовательно, воздействие входов должно оцениваться в короткий промежуток времени, то есть воздействие синапса имеет временный характер.
Результат сумматора проходит пороговую функцию (б), если сумма превосходит пороговое значение, то это приводит к активности нейрона.
При активации нейрон сигнализирует о своей активности системе, передовая информацию о своём положении в пространстве нервной системы и заряде, изменяемом во времени (в).
Через определённое время, после активации нейрон передает возбуждение по всем имеющимся синапсам, предварительно производя пересчет их силы. Весь период активации нейрон перестает реагировать на внешние раздражители, то есть все воздействия синапсов других нейронов игнорируются. В период активации входит так же период восстановления нейрона.
Происходит корректировка вектора Т (г) с учётом значения точки паттерна Pp и уровнем нейропластичности. Далее происходит переоценка значений всех сил синапсов в нейроне(д).
Обратите внимание, что блоки (г) и (д) выполняются параллельно с блоком (в).
ЭФФЕКТ ВОЛНЫ
Если внимательно проанализировать предложенную модель, то можно увидеть, что источник возбуждения должен оказывать большее влияние на нейрон, чем другой удалённый, активный участок мозга. Следовательно возникает вопрос: почему же все равно происходит передача в направлении другого активного участка?
Данную проблему я смог определить, только создав компьютерную модель. Решение подсказал график изменения мембранного потенциала при активности нейрона.

Усиленная реполяризация нейрона, как говорилось ранее, имеет важное значение для нервной системы, благодаря ей создается эффект волны, стремление нервного возбуждения распространятся от источника возбуждения.
При работе с моделью я наблюдал два эффекта, ели пренебречь следовым потенциалом или сделать его недостаточно большим, то возбуждение не распространяется от источников, а в большей степени стремится к локализации. Если сделать следовой потенциал сильно большим, то возбуждение стремится «разбежаться» в разные стороны, не только от своего источника, но и от других.
КОГНИТИВНАЯ КАРТА
Используя теорию электромагнитного взаимодействия, можно объяснить многие явления и сложные процессы, протекающие в нервной системе. К примеру, одним из последних открытий, которое широко обсуждается в науках о мозге, является открытие когнитивных карт в гиппокампе.
Гиппокамп – это отдел мозга, которому отвечает за кратковременную память. Эксперименты на крысах выявили, что определённому месту в лабиринте соответствует своя локализованная группа клеток в гиппокампе, причем, не имеет значение, как животное попадает в это место, все равно будет активирован соответствующий этому месту участок нервной ткани. Естественно, животное должно помнить данный лабиринт, не стоит рассчитывать на топологическое соответствие пространства лабиринта и когнитивной карты.

Каждое место в лабиринте представляется в мозге, как совокупность раздражителей различного характера: запахи, цвет стен, возможные примечательные объекты, характерные звуки и т. д. Указанные раздражители отражаются на коре, различных представительствах органов чувств, в виде всплесков активности в определённых комбинациях. Мозг одновременно обрабатывает информацию в нескольких отделах, зачастую информационные каналы разделяются, одна и та же информация поступает в различные участки мозга.

Активация нейронов места в зависимости от положения в лабиринте (активность разных нейронов показана разным цветом). источник
Гиппокамп расположен в центре мозга, вся кара и её области удалены от него, на одинаковые расстояния. Если определить для каждой уникальной комбинации раздражителей точку масс зарядов поверхностей нейронов, то можно увидеть, что указанные точки будут различны, и будут находиться примерно в центре мозга. К этим точкам будет стремиться и распространятся возбуждение в гиппокампе, формируя устойчивые участки возбуждения. Более того, поочередная смена комбинаций раздражителей, будет приводить к смещению точки паттерна. Участки когнитивной карты будут ассоциативно связываться друг с другом последовательно, что приведет к тому, что животное, помещенное в начало знакомого ей лабиринта, может вспомнить весь последующий путь.
Заключение
У многих возникнет вопрос, где в данной работе предпосылки к элементу разумности или проявления высшей интеллектуальной деятельности?
Важно отметить, что феномен человеческого поведения, есть следствие функционирования биологической структуры. Следовательно, чтобы имитировать разумное поведение, необходимо хорошо понимать принципы и особенности функционирования биологических структур. К сожалению, в науке биологии пока не представлен четкий алгоритм: как работает нейрон, как понимает, куда необходимо отращивать свои дендриты, как настроить свои синапсы, что бы в нервной системе смог сформироваться простой условный рефлекс, на подобие тех, которые демонстрировал и описывал в своих работах академик И.П. Павлов.
С другой стороны в науке об искусственном интеллекте, в восходящем (биологическом) подходе, сложилось парадоксальная ситуация, а именно: когда используемые в исследованиях модели основаны на устаревших представлениях о биологическом нейроне, консерватизм, в основе которого берётся персептрон без переосмысления его основных принципов, без обращения к биологическому первоисточнику, придумывается все более хитроумные алгоритмы и структуры, не имеющих биологических корней.
Конечно, никто не уменьшает достоинств классических нейронных сетей, которые дали множество полезных программных продуктов, но игра с ними не является путем к созданию интеллектуально действующей системы.
Более того, не редки заявления, о том, что нейрон подобен мощной вычислительной машине, приписывают свойство квантовых компьютеров. Из-за этой сверхсложности, нервной системе приписывается невозможность её повторения, ведь это соизмеримо с желанием смоделировать человеческую душу. Однако, в реальности природа идет по пути простоты и элегантности своих решений, перемещение зарядов на мембране клетки может служить, как для передачи нервного возбуждения, так и для трансляции информации о том, где происходит данная передача.
Несмотря на то, что указанная работа демонстрирует, как образуются элементарные условные рефлексы в нервной системе, она приближает к пониманию того, что такое интеллект и разумная деятельность.
Существуют еще множество аспектов работы нервной системы: механизмы торможения, принципы построения эмоций, организация безусловных рефлексов и обучение, без которых невозможно построить качественную модель нервной системы. Есть понимание, на интуитивном уровне, как работает нервная система, принципы которой возможно воплотить в моделях.
Создание первой модели помогли отработать и откорректировать представление об электромагнитном взаимодействии нейронов. Понять, как происходит формирование рефлекторных дуг, как каждый отдельный нейрон понимает, каким образом ему настроить свои синапсы для получения ассоциативных связей.
На данный момент я начал разрабатывать новую версию программы, которая позволит смоделировать многие другие аспекты работы нейрона и нервной системы.
Прошу принять активное участие в обсуждении выдвинутых здесь гипотез и предположений, так как я могу относиться к своим идеям предвзято. Ваше мнение очень важно для меня.
Модель(Windows PC) + туториал
Электрически возбудимая клетка, которая общается через синапсы
| Нейрон | |
|---|---|
| Анатомия мультиполярного нейрона |
|
| Идентификаторы | |
| MeSH | D009474 |
| NeuroLex ID | sao1417703748 |
| TA98 | A14.0.00.002 |
| TH | H2.00.06.1.00002 |
| FMA | 54527 |
| Анатомические термины нейроанатомии [редактировать в Викиданных ] |
Схема анатомически точного одиночного пирамидного нейрона, первичного возбуждающего нейрона коры головного мозга, с синаптическим соединением от входящего аксона к дендритному позвоночнику.
A нейрон или нерв ячейка — это электрически возбудимая ячейка, которая связывается с другими ячейками через специализированные соединения, называемые синапсами. Он является основным компонентом нервной ткани у всех животных, кроме губок и плакозоа. Растения и грибы не имеют нервных клеток. Написание нейрон стало необычным.
Нейроны обычно делятся на три типа в зависимости от их функции. Сенсорные нейроны реагируют на стимулы, такие как прикосновение, звук или свет, которые воздействуют на клетки органов чувств, и отправляют сигналы в спинной или головной мозг. Моторные нейроны получают сигналы от головного и спинного мозга, чтобы контролировать все, от мышечных сокращений до железистого выхода. Интернейроны соединяют нейроны с другими нейронами в той же области головного или спинного мозга. Группа связанных нейронов называется нейронной цепью.
Типичный нейрон состоит из тела клетки (сома ), дендритов и одного аксона. Сома обычно компактная. Аксон и дендриты — это нити, которые выходят из него. Дендриты обычно обильно разветвляются и выходят на несколько сотен микрометров от сомы. Аксон покидает сому в области вздутия, называемого бугорком аксона, и перемещается на расстояние до 1 метра у людей или более у других видов. Он разветвляется, но обычно имеет постоянный диаметр. На самом дальнем конце ветвей аксона находятся терминалы аксона, где нейрон может передавать сигнал через синапс другой клетке. Нейроны могут не иметь дендритов или аксонов. Термин нейрит используется для описания либо дендрита, либо аксона, особенно когда клетка недифференцированная.
Большинство нейронов получают сигналы через дендриты и сомы и отправляют их. сигналы вниз по аксону. В большинстве синапсов сигналы переходят от аксона одного нейрона к дендриту другого. Однако синапсы могут соединять аксон с другим аксоном или дендрит с другим дендритом.
Сигнальный процесс частично электрический, а частично химический. Нейроны электрически возбудимы из-за поддержания градиентов напряжения на их мембранах. Если напряжение изменяется на достаточно большую величину за короткий промежуток времени, нейрон генерирует электрохимический импульс все или ничего , называемый потенциалом действия. Этот потенциал быстро распространяется по аксону и активирует синаптические связи, когда достигает их. Синаптические сигналы могут быть возбуждающими или тормозными, увеличивая или уменьшая сетевое напряжение, которое достигает сомы.
В большинстве случаев нейроны генерируются нервными стволовыми клетками во время развития мозга и в детстве. Нейрогенез в большинстве областей мозга в большинстве случаев прекращается в зрелом возрасте. Однако убедительные доказательства подтверждают образование значительного числа новых нейронов в гиппокампе и обонятельной луковице.
Содержание
- 1 Нервная система
- 2 Анатомия и гистология
- 2.1 Мембрана
- 2.2 Гистология и внутреннее строение
- 3 Классификация
- 3.1 Структурная классификация
- 3.1.1 Полярность
- 3.1.2 Другое
- 3.2 Функциональная классификация
- 3.2.1 Направление
- 3.2. 2 Воздействие на другие нейроны
- 3.2.3 Паттерны разряда
- 3.2.4 Нейротрансмиттер
- 3.1 Структурная классификация
- 4 Связность
- 5 Механизмы распространения потенциалов действия
- 6 Нейронное кодирование
- 7 Принцип «все или ничего»
- 8 Этимология и орфография
- 9 История
- 9.1 Доктрина нейронов
- 9.2 Компартментная модель нейронов
- 10 Нейроны в головном мозге
- 11 Неврологические расстройства
- 11.1 Демиелинизация
- 11.2 Аксональная дегенерация
- 12 Нейрогенез
- 13 Регенерация нервов
- 14 См. также
- 15 Ссылки
- 16 Дополнительная литература
- 17 Внешние ссылки
Нервная система
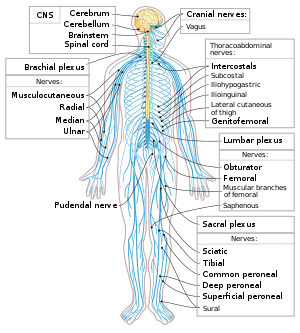 Схема нервной системы человека система. Продемонстрирована взаимосвязь между головным и спинным мозгом и остальными нервами в организме.
Схема нервной системы человека система. Продемонстрирована взаимосвязь между головным и спинным мозгом и остальными нервами в организме.
Нейроны являются основными компонентами нервной системы, наряду с глиальными клетками, которые обеспечивают им структурные и метаболические функции. поддержка. Нервная система состоит из центральной нервной системы, которая включает головной и спинной мозг, а также периферическую нервную систему, который включает вегетативную и соматическую нервную систему. У позвоночных большинство нейронов принадлежит центральной нервной системе, но некоторые находятся в периферических ганглиях, а многие сенсорные нейроны расположены в органах чувств, таких как сетчатка и улитка.
Аксоны могут объединяться в пучки, которые составляют нервы в периферической нервной системе (как нити проволоки составляют кабели). Связки аксонов в центральной нервной системе называются трактами.
Анатомия и гистология
| Нейрон (периферическая нервная система ) |
|---|
 Дендрит Сома Аксон Ядро Узел. Ранвье Аксонный терминал Шванновская клетка Миелиновая оболочка Дендрит Сома Аксон Ядро Узел. Ранвье Аксонный терминал Шванновская клетка Миелиновая оболочка |
Нейроны в высшей степени специализированы для обработки и передачи клеточных сигналов. Учитывая их разнообразие функций выполняемые в разных частях нервной системы, они могут быть самыми разными по форме, размеру и электрохимическим свойствам. Например, сома нейрона может варьироваться от 4 до 100 микрометров в диаметре.
- сома — это тело нейрона. Поскольку она содержит ядро , здесь происходит большая часть синтеза белка. Ядро может варьироваться от 3 до 18 микрометров в диаметре.
- дендриты нейрона представляют собой клеточные продолжения с множеством ветвей. Эта общая форма и структура обозначаются метафорически как дендритное дерево. Это то место, где большая часть входа в нейрон происходит через дендритный позвоночник.
- аксон — более тонкий, похожий на кабель выступ, который может простираться на десятки, сотни, или даже в десятки тысяч раз больше диаметра сомы по длине. Аксон в первую очередь несет нервные сигналы от сомы и возвращает к ней некоторые типы информации. Многие нейроны имеют только один аксон, но этот аксон может — и обычно будет — подвергаться обширному разветвлению, обеспечивая связь со многими клетками-мишенями. Часть аксона, в которой он выходит из сомы, называется бугорком аксона. Помимо анатомической структуры, холмик аксона также имеет наибольшую плотность потенциалзависимых натриевых каналов. Это делает его наиболее легко возбуждаемой частью нейрона и зоной инициации спайков для аксона. С точки зрения электрофизиологии, он имеет самый отрицательный пороговый потенциал.
- Хотя аксон и бугорок аксона обычно участвуют в оттоке информации, эта область также может получать входные данные от других нейронов.
- конец аксона находится на конце аксона, наиболее удаленном от сомы, и содержит синапсы. Синаптические бутоны — это специализированные структуры, в которых выделяются химические вещества нейромедиатора для связи с нейронами-мишенями. Помимо синаптических бутонов на конце аксона, нейрон может иметь проходные бутоны, которые расположены по длине аксона.
Тело клетки нейрона
Общепринятая точка зрения на то, что нейрон приписывает специальные функции его различным анатомическим особенностям компоненты; однако дендриты и аксоны часто действуют вопреки своей так называемой основной функции.
Схема типичного миелинизированного двигательного нейрона позвоночного  Воспроизвести медиа Видео о неврологии
Воспроизвести медиа Видео о неврологии
Аксоны и дендриты в центральной нервной системе обычно имеют толщину всего около одного микрометра, в то время как некоторые в периферической нервной системе намного толще. Сома обычно имеет диаметр около 10–25 микрометров и часто ненамного больше ядра клетки, которое она содержит. Самый длинный аксон человеческого мотонейрона может быть более метра в длину, простираясь от основания позвоночника до пальцев ног.
Сенсорные нейроны могут иметь аксоны, которые проходят от пальцев ног до заднего столба спинного мозга на высоте более 1,5 метров у взрослых. Жирафы имеют одиночные аксоны длиной в несколько метров, идущие по всей длине их шеи. Многое из того, что известно о функции аксонов, получено в результате изучения гигантского аксона кальмара, идеального экспериментального препарата из-за его относительно огромных размеров (0,5–1 миллиметр в толщину, несколько сантиметров в длину).
Полностью дифференцированные нейроны являются постоянно постмитотическими, однако стволовые клетки, присутствующие в мозге взрослого человека, могут регенерировать функциональные нейроны на протяжении всей жизни организма (см. нейрогенез ). Астроциты представляют собой звездообразные глиальные клетки. Было замечено, что они превращаются в нейроны в силу их сходных со стволовыми клетками характеристики плюрипотентности.
мембраны
Как и все животные клетки, тело каждого нейрона окружено плазматическая мембрана, бислой липидных молекул со встроенными в него многими типами белковых структур. Липидный бислой представляет собой мощный электрический изолятор, но в нейронах многие из белковых структур, встроенных в мембрану, электрически активны. К ним относятся ионные каналы, которые позволяют электрически заряженным ионам проходить через мембрану, и ионные насосы, которые химически переносят ионы с одной стороны мембраны на другую. Большинство ионных каналов проницаемы только для определенных типов ионов. Некоторые ионные каналы ограничены напряжением, что означает, что они могут переключаться между открытым и закрытым состояниями путем изменения разности напряжений на мембране. Другие являются химически закрытыми, что означает, что они могут переключаться между открытым и закрытым состояниями посредством взаимодействия с химическими веществами, которые диффундируют через внеклеточную жидкость. Материалы иона включают натрий, калий, хлорид и кальций. Взаимодействие между ионными каналами и ионными насосами создает разность напряжений на мембране, обычно немного меньше 1/10 вольта на исходном уровне. Это напряжение выполняет две функции: во-первых, оно обеспечивает источник питания для ряда зависимых от напряжения белковых механизмов, встроенных в мембрану; во-вторых, он обеспечивает основу для передачи электрического сигнала между различными частями мембраны.
Гистология и внутренняя структура
нейроны, окрашенные по Гольджи в ткани гиппокампа человека Актиновые нити в кортикальном нейроне мыши в культуре
Многочисленные микроскопические скопления, называемые тельцами Ниссля (или Субстанция Ниссля) наблюдаются при окрашивании тел нервных клеток базофильным («любящим основание») красителем. Эти структуры состоят из грубого эндоплазматического ретикулума и связанной с ним рибосомной РНК. Названные в честь немецкого психиатра и невропатолога Франца Ниссля (1860–1919), они участвуют в синтезе белка, и их значимость можно объяснить тем фактом, что нервные клетки очень метаболически активны. Базофильные красители, такие как анилин или (слабо) гематоксилин, выделяют отрицательно заряженные компоненты и, таким образом, связываются с фосфатным остовом рибосомной РНК.
Тело клетки нейрона поддерживается сложной сеткой структурных белков, называемых нейрофиламентами, которые вместе с нейротрубочками (нейрональными микротрубочками) собраны в более крупные нейрофибриллы. Некоторые нейроны также содержат гранулы пигмента, такие как нейромеланин (коричневато-черный пигмент, являющийся побочным продуктом синтеза катехоламинов ) и липофусцин (желтовато-коричневый пигмент), оба из которых накапливаются с возрастом. Другими структурными белками, которые важны для функции нейронов, являются актин и тубулин из микротрубочек. β-тубулин класса III обнаруживается почти исключительно в нейронах. Актин преимущественно находится на концах аксонов и дендритов во время развития нейронов. Там динамика актина может модулироваться посредством взаимодействия с микротрубочками.
Между аксонами и дендритами существуют разные внутренние структурные характеристики. Типичные аксоны почти никогда не содержат рибосомы, за исключением некоторых в начальном сегменте. Дендриты содержат гранулярную эндоплазматическую сеть или рибосомы, количество которых уменьшается по мере увеличения расстояния от тела клетки.
Классификация
Изображение пирамидных нейронов в мышиной коре головного мозга, экспрессирующих зеленый флуоресцентный белок. Красное окрашивание указывает на ГАМКергические интернейроны. Окрашенные SMI32 пирамидные нейроны в коре головного мозга
Нейроны различаются по форме и размеру и могут быть классифицированы по их морфологии и функция. Анатом Камилло Гольджи сгруппировал нейроны на два типа; тип I с длинными аксонами, используемый для перемещения сигналов на большие расстояния, и тип II с короткими аксонами, которые часто можно спутать с дендритами. Клетки типа I можно дополнительно классифицировать по расположению сомы. Основная морфология нейронов типа I, представленных спинномозговыми мотонейронами, состоит из тела клетки, называемого сомой, и длинного тонкого аксона, покрытого миелиновой оболочкой. Дендритное дерево обвивает тело клетки и получает сигналы от других нейронов. Конец аксона имеет терминалы ветвления (терминал аксона ), которые высвобождают нейротрансмиттеры в промежуток, называемый синаптической щелью, между терминалами и дендритами следующего нейрона.
Структурная классификация
Полярность
Различные типы нейронов:. 1 Униполярный нейрон. 2 Биполярный нейрон. 3 Мультиполярный нейрон. 4 Псевдоуниполярный нейрон
Большинство нейронов можно анатомически охарактеризовать как:
- униполярный : одиночный отросток
- биполярный : 1 аксон и 1 дендрит
- мультиполярный : 1 аксон и 2 или более дендритов
- Гольджи I : нейроны с выступающими аксональными отростками; примерами являются пирамидные клетки, клетки Пуркинье и клетки переднего рога
- Golgi II : нейроны, аксональные отростки которых проецируются локально; лучший пример — гранулярная клетка
- анаксоническая : где аксон нельзя отличить от дендрита (ов)
- Псевдоуниполярный : 1 процесс, который затем служит и аксоном, и дендритом
Прочее
Некоторые уникальные типы нейронов можно идентифицировать по их расположению в нервной системе и отличительной форме. Некоторые примеры:
- клетки корзины, интернейроны, которые образуют плотное сплетение терминалей вокруг сомы клеток-мишеней, обнаруженные в коре и мозжечке
- клетки Беца, большие двигательные нейроны
- клетки Лугаро, интернейроны мозжечка
- средние шиповатые нейроны, большинство нейронов в полосатом теле
- клетки Пуркинье, огромные нейроны мозжечка, тип Гольджи I мультиполярный нейрон
- пирамидные клетки, нейроны с треугольной сомой, тип клеток Гольджи I
- клеток Реншоу, нейроны с обоими концами, связанными с альфа-моторными нейронами
- униполярными щеточными клетками, интернейроны с уникальным дендритом, заканчивающимся щеткообразным пучком
- Гранулярные клетки, тип нейрона Гольджи II
- клетки переднего рога, мотонейроны, расположенные в спинном мозге шнур
- Веретеновидные клетки, интернейроны, которые соединяют широко разделенные области мозга
Функциональная классификация
Направление
- Афферентные нейроны передают информацию от тканей и органов в центральную нервную систему и также называются сенсорными нейронами.
- Эфферентные нейроны (двигательные нейроны) передают сигналы от центральной нервной системы к эффекторным клеткам.
- Интернейроны соединяют нейроны внутри определенных области центральной нервной системы.
Афферентный и эфферентный также обычно относятся к нейронам, которые, соответственно, передают информацию или отправляют информацию из мозга.
Воздействие на другие нейроны
Нейрон влияет на другие нейроны, высвобождая нейротрансмиттер, который связывается с химическими рецепторами. Воздействие на постсинаптический нейрон определяется типом рецептора, который активируется, а не пресинаптическим нейроном или нейромедиатором. Нейромедиатор можно рассматривать как ключ, а рецептор — как замок: один и тот же нейромедиатор может активировать несколько типов рецепторов. Рецепторы можно в широком смысле классифицировать как возбуждающие (вызывающие увеличение скорости стрельбы), тормозящие (вызывающие снижение скорости стрельбы) или модулирующие (вызывающие длительные эффекты, напрямую не связанные со скоростью стрельбы).
Два наиболее распространенных (90% +) нейротрансмиттеров в головном мозге, глутамат и ГАМК, имеют в основном согласованные действия. Глутамат действует на несколько типов рецепторов и оказывает возбуждающее действие на ионотропные рецепторы и модулирующее действие на метаботропные рецепторы. Точно так же ГАМК действует на несколько типов рецепторов, но все они обладают ингибирующим действием (по крайней мере, у взрослых животных). Из-за такой последовательности нейробиологи обычно называют клетки, которые выделяют глутамат, «возбуждающими нейронами», а клетки, которые выделяют ГАМК, — «тормозящими нейронами». Некоторые другие типы нейронов обладают последовательными эффектами, например, «возбуждающие» мотонейроны в спинном мозге, которые высвобождают ацетилхолин, и «тормозящие» спинномозговые нейроны, которые выделяют глицин.
Различие между возбуждающими и тормозными нейротрансмиттерами не является абсолютным. Скорее, это зависит от класса химических рецепторов, присутствующих в постсинаптическом нейроне. В принципе, отдельный нейрон, высвобождая единственный нейротрансмиттер, может оказывать возбуждающее действие на одни мишени, подавляющее действие на другие и модулирующее воздействие на другие. Например, фоторецепторные клетки в сетчатке постоянно выделяют глутамат нейромедиатора в отсутствие света. Так называемые OFF биполярные клетки, как и большинство нейронов, возбуждаются высвобожденным глутаматом. Однако соседние нейроны-мишени, называемые биполярными клетками ON, вместо этого ингибируются глутаматом, потому что они не имеют типичных ионотропных рецепторов глутамата и вместо этого экспрессируют класс ингибирующих метаботропных рецепторов глутамата.. Когда присутствует свет, фоторецепторы перестают выделять глутамат, который освобождает ON биполярные клетки от ингибирования, активируя их; это одновременно снимает возбуждение с выключенных биполярных клеток, заставляя их замолчать.
На основе белков, экспрессируемых пресинаптическим нейроном, можно определить тип ингибирующего эффекта, который пресинаптический нейрон будет оказывать на постсинаптический нейрон. Парвальбумин -экспрессирующие нейроны обычно подавляют выходной сигнал постсинаптического нейрона в зрительной коре, тогда как соматостатин -экспрессирующие нейроны обычно блокируют дендритные входы в постсинаптический нейрон.
Разрядные паттерны
Нейроны обладают внутренними электроответственными свойствами, такими как внутреннее трансмембранное напряжение колебательные паттерны. Таким образом, нейроны можно классифицировать по их электрофизиологическим характеристикам:
- Тонические или регулярные пики. Некоторые нейроны обычно постоянно (тонически) активны, как правило, с постоянной частотой. Пример: интернейроны в нейростриатуме.
- фазовый или разрывной. Нейроны, которые стреляют очередями, называются фазическими.
- Быстрые пики. Некоторые нейроны отличаются высокой частотой возбуждения, например, некоторые типы корковых тормозных интернейронов, клетки в бледном шаре, ганглиозные клетки сетчатки.
Нейротрансмиттер Синаптические пузырьки, содержащие нейротрансмиттеры
Синаптические пузырьки, содержащие нейротрансмиттеры
- Холинергические нейроны — ацетилхолин. Ацетилхолин высвобождается из пресинаптических нейронов в синаптическую щель. Он действует как лиганд как для лиганд-управляемых ионных каналов, так и для метаботропных (GPCR) мускариновых рецепторов. Никотиновые рецепторы представляют собой ионные каналы, управляемые пентамерным лигандом, состоящие из альфа- и бета-субъединиц, которые связывают никотин. Связывание лиганда открывает канал, вызывая приток деполяризации Na и увеличивает вероятность высвобождения пресинаптического нейротрансмиттера. Ацетилхолин синтезируется из холина и ацетилкофермента A.
- ГАМКергических нейронов — гамма-аминомасляной кислоты. ГАМК является одним из двух нейроингибиторов в центральной нервной системе (ЦНС), наряду с глицином. ГАМК имеет функцию, гомологичную ACh, блокируя анионные каналы, которые позволяют ионам Cl проникать в постсинаптический нейрон. Cl вызывает гиперполяризацию внутри нейрона, уменьшая вероятность срабатывания потенциала действия по мере того, как напряжение становится более отрицательным (для срабатывания потенциала действия необходимо достичь положительного порога напряжения). ГАМК синтезируется из глутаматных нейротрансмиттеров с помощью фермента глутаматдекарбоксилазы.
- глутаматергических нейронов — глутамата. Глутамат является одним из двух основных нейротрансмиттеров возбуждающих аминокислот, наряду с аспартатом. Глутаматные рецепторы относятся к одной из четырех категорий, три из которых представляют собой ионные каналы, управляемые лигандами, а одна из которых — рецептор, связанный с G-белком (часто называемый GPCR).
-
- AMPA и Kainate рецепторы функционируют как каналы катионов, проницаемые для каналов катионов натрия, обеспечивающие быструю возбуждающую синаптическую передачу.
- рецепторы NMDA представляют собой еще один катионный канал, который более проницаем для Ca. Функция рецепторов NMDA зависит от связывания рецептора глицина в качестве со- агониста в поре канала. Рецепторы NMDA не функционируют без присутствия обоих лигандов.
- Метаботропные рецепторы, GPCR модулируют синаптическую передачу и постсинаптическую возбудимость.
- Глутамат может вызывать эксайтотоксичность, когда кровоток в мозг прерывается, что приводит к повреждению мозга. Когда кровоток подавляется, глутамат высвобождается из пресинаптических нейронов, вызывая большую активацию рецепторов NMDA и AMPA, чем обычно, вне стрессовых условий, что приводит к повышенному содержанию Ca и Na в постсинаптическом нейроне и повреждению клеток. Глутамат синтезируется из аминокислоты глутамина с помощью фермента глутаматсинтазы.
- допаминергических нейронов — допамина. Дофамин представляет собой нейротрансмиттер, который действует на Gs-сопряженные рецепторы типа D1 (D1 и D5), которые увеличивают цАМФ и PKA, и рецепторы типа D2 (D2, D3 и D4), которые активируют Gi-сопряженные рецепторы которые уменьшают цАМФ и ПКА. Дофамин связан с настроением и поведением и модулирует как пре-, так и постсинаптическую нейротрансмиссию. Утрата дофаминовых нейронов в черной субстанции была связана с болезнью Паркинсона. Дофамин синтезируется из аминокислоты тирозин. Тирозин катализируется в левадопа (или L-DOPA ) с помощью тирозингидрокслазы, а затем левадопа превращается в дофамин ароматической аминокислотой декарбоксилазой.
- Серотонинергические нейроны — серотонин. Серотонин (5-гидрокситриптамин, 5-HT) может действовать как возбуждающее или тормозящее действие. Из четырех классов рецепторов 5-HT 3 являются GPCR, а 1 — лиганд-зависимым катионным каналом. Серотонин синтезируется из триптофана с помощью гидроксилазы триптофана, а затем декарбоксилазы. Недостаток 5-HT в постсинаптических нейронах был связан с депрессией. Для лечения используются препараты, которые блокируют пресинаптический переносчик серотонина, такие как Прозак и Золофт.
- Гистаминергические нейроны — гистамин. Гистамин представляет собой нейромедиатор моноаминов и нейромодулятор. Нейроны, продуцирующие гистамин, обнаруживаются в туберомаммиллярном ядре гипоталамуса. Гистамин участвует в возбуждении и регулирует поведение во сне / бодрствовании.
- AMPA и Kainate рецепторы функционируют как каналы катионов, проницаемые для каналов катионов натрия, обеспечивающие быструю возбуждающую синаптическую передачу.
- рецепторы NMDA представляют собой еще один катионный канал, который более проницаем для Ca. Функция рецепторов NMDA зависит от связывания рецептора глицина в качестве со- агониста в поре канала. Рецепторы NMDA не функционируют без присутствия обоих лигандов.
- Метаботропные рецепторы, GPCR модулируют синаптическую передачу и постсинаптическую возбудимость.
- Глутамат может вызывать эксайтотоксичность, когда кровоток в мозг прерывается, что приводит к повреждению мозга. Когда кровоток подавляется, глутамат высвобождается из пресинаптических нейронов, вызывая большую активацию рецепторов NMDA и AMPA, чем обычно, вне стрессовых условий, что приводит к повышенному содержанию Ca и Na в постсинаптическом нейроне и повреждению клеток. Глутамат синтезируется из аминокислоты глутамина с помощью фермента глутаматсинтазы.
Связность
Сигнал, распространяющийся по аксону к телу клетки и дендритам следующей клетки Химический синапс
Нейроны связываются друг с другом через синапсы, где либо конец аксона одной клетки контактирует с дендритом другого нейрона, сомой или, что реже, аксоном. Нейроны, такие как клетки Пуркинье в мозжечке, могут иметь более 1000 дендритных ветвей, соединяющихся с десятками тысяч других клеток; другие нейроны, такие как магноклеточные нейроны супраоптического ядра, имеют только один или два дендрита, каждый из которых получает тысячи синапсов.
Синапсы могут быть возбуждающими или тормозными, увеличивая или уменьшая активность в целевом нейроне соответственно. Некоторые нейроны также взаимодействуют через электрические синапсы, которые представляют собой прямые, электрически проводящие соединения между клетками.
Когда потенциал действия достигает конца аксона, он открывает потенциалзависимые кальциевые каналы, позволяя ионам кальция проникать в терминал. Кальций заставляет синаптические везикулы, заполненные молекулами нейротрансмиттера, сливаться с мембраной, высвобождая их содержимое в синаптическую щель. Нейромедиаторы диффундируют через синаптическую щель и активируют рецепторы постсинаптического нейрона. Высокий уровень цитозольного кальция в конце аксона запускает поглощение кальция митохондриями, что, в свою очередь, активирует митохондриальный энергетический метаболизм с образованием АТФ для поддержки непрерывной нейротрансмиссии.
autapse — это синапс, в котором аксон нейрона соединяется с его собственными дендритами.
человеческий мозг имеет примерно 8,6 x 10 (восемьдесят шесть миллиардов) нейронов. Каждый нейрон имеет в среднем 7000 синаптических связей с другими нейронами. Было подсчитано, что мозг трехлетнего ребенка имеет около 10 синапсов (1 квадриллион). Это число снижается с возрастом и стабилизируется к зрелому возрасту. Для взрослого человека оценки варьируются от 10 до 5 x 10 синапсов (от 100 до 500 триллионов).
Аннотированная диаграмма стадий распространения потенциала действия по аксону, включая роль концентрации ионов и белков насоса и канала.
Механизмы распространения потенциалов действия
В 1937 году Джон Закари Янг предположил, что гигантский аксон кальмара может быть использован для изучения электрических свойств нейронов. Он больше, чем нейроны человека, но похож на него, что упрощает его изучение. Путем введения электродов в аксоны гигантских кальмаров были выполнены точные измерения мембранного потенциала .
. Клеточная мембрана аксона и сомы содержит управляемые по напряжению ионные каналы, которые позволяют нейрону генерировать и распространять электрический сигнал ( потенциал действия). Некоторые нейроны также генерируют подпороговые колебания мембранного потенциала. Эти сигналы генерируются и распространяются несущими заряд ионами, включая натрий (Na), калий (K), хлорид (Cl) и кальций (Ca).
Некоторые стимулы могут активировать нейрон приводящие к электрической активности, включая давление, растяжение, химические передатчики и изменения электрического потенциала на клеточной мембране. Стимулы заставляют определенные ионные каналы внутри клеточной мембраны открываться, что приводит к потоку ионов через клеточную мембрану, изменяющему мембранный потенциал. Нейроны должны поддерживать определенные электрические свойства, которые определяют их тип нейрона.
Тонкие нейроны и аксоны требуют меньше метаболических затрат на производство и перенос потенциалов действия, но более толстые аксоны передают импульсы быстрее. Чтобы минимизировать метаболические затраты при сохранении быстрой проводимости, многие нейроны имеют изолирующие оболочки из миелина вокруг своих аксонов. Оболочки образованы глиальными клетками: олигодендроцитами в центральной нервной системе и шванновскими клетками в периферической нервной системе. Оболочка позволяет потенциалам действия перемещаться быстрее,, чем в немиелинизированных аксонах того же диаметра, при этом потребляя меньше энергии. Миелиновая оболочка в периферических нервах обычно проходит вдоль аксона на участках длиной около 1 мм, перемежающихся незащищенными узлами Ранвье, которые содержат высокую плотность потенциалзависимых ионных каналов. Рассеянный склероз — неврологическое заболевание, возникающее в результате демиелинизации аксонов в центральной нервной системе.
Некоторые нейроны не генерируют потенциалы действия, а вместо этого генерируют ступенчатый электрический сигнал , который, в свою очередь, вызывает постепенное высвобождение нейромедиатора. Такие нейроны без пиков, как правило, являются сенсорными нейронами или интернейронами, поскольку они не могут переносить сигналы на большие расстояния.
Нейронное кодирование
Нейронное кодирование связано с тем, как сенсорная и другая информация представлена в мозге нейронами. Основная цель изучения нейронного кодирования — охарактеризовать взаимосвязь между стимулом и индивидуальными или ансамблевыми ответами нейронов, а также взаимосвязи между электрической активностью нейронов внутри ансамбля. Считается, что нейроны могут кодировать как цифровую, так и аналоговую информацию.
Принцип «все или ничего»
Пока стимул достигает порогового значения, будет дан полный ответ. Более сильный стимул не приводит к большему ответу, наоборот.
Проведение нервных импульсов является примером реакции все или ничего. Другими словами, если нейрон вообще отвечает, то он должен отвечать полностью. Более интенсивная стимуляция, например более яркое изображение / более громкий звук, не дает более сильного сигнала, но может увеличить частоту возбуждения. Рецепторы по-разному реагируют на раздражители. Медленно приспосабливающиеся или тонические рецепторы реагируют на устойчивый раздражитель и производят постоянную скорость возбуждения. Тонические рецепторы чаще всего реагируют на повышенную интенсивность стимула увеличением своей частоты срабатывания, обычно как силовая функция стимула в зависимости от количества импульсов в секунду. Это можно сравнить с внутренним свойством света, когда для большей интенсивности определенной частоты (цвета) требуется больше фотонов, поскольку фотоны не могут стать «сильнее» для определенной частоты.
Другие типы рецепторов включают быстро адаптирующиеся или фазовые рецепторы, у которых возбуждение уменьшается или прекращается при устойчивом раздражении; Примеры включают skin, при прикосновении к которой нейроны срабатывают, но если объект поддерживает равномерное давление, нейроны перестают активироваться. Нейроны кожи и мышц, которые реагируют на давление и вибрацию, имеют фильтрующие вспомогательные структуры, которые помогают им функционировать.
пачинское корпускула — одна из таких структур. Он имеет концентрические слои, как у луковицы, которые образуются вокруг окончания аксона. Когда прикладывается давление и тельце деформируется, механический стимул передается аксону, который срабатывает. Если давление устойчиво, стимул прекращается; таким образом, обычно эти нейроны отвечают временной деполяризацией во время начальной деформации и снова, когда давление снимается, что заставляет тельце снова изменять форму. Другие типы адаптации важны для расширения функций ряда других нейронов.
Этимология и орфография
Немецкий анатом Генрих Вильгельм Вальдейер ввел термин нейрон в 1891 году., основанный на древнегреческом νεῦρον нейроне «сухожилие, шнур, нерв».
Слово было принято во французском языке с орфографическим нейроном. Это написание также использовалось многими английскими писателями, но теперь оно стало редким в американском употреблении и необычным в использовании в Великобритании.
История
Рисунок Камилло Гольджи гиппокампа, окрашенный с использованием нитрат серебра метод Рисунок клетки Пуркинье в коре мозжечка, выполненный Сантьяго Рамоном-и-Кахалем, демонстрирующий способность метода окрашивания Гольджи выявлять мелкие детали
Место нейрона как первичной функциональной единицы нервной системы было впервые обнаружено в конце 19 века благодаря работам испанского анатома Сантьяго Рамона-и-Кахала.
Чтобы сделать структуру отдельных нейронов видимой, Рамон и Кахал усовершенствовал процесс окрашивания серебром, который был разработан Камилло Гольджи. Усовершенствованный процесс включает в себя технику, называемую «двойной пропиткой», и все еще используется.
В 1888 году Рамон-и-Кахаль опубликовал статью о мозжечке птиц. В этой статье он заявил, что не может найти доказательств анастомоза между аксонами и дендритами, и назвал каждый нервный элемент «абсолютно автономным кантоном». Это стало известно как нейронная доктрина, один из центральных принципов современной нейробиологии.
. В 1891 году немецкий анатом Генрих Вильгельм Вальдейер написал очень влиятельный обзор доктрина нейронов, в которой он ввел термин нейрон для описания анатомической и физиологической единицы нервной системы.
Пятна серебряной импрегнации — полезный метод для нейроанатомических исследований, потому что по неизвестным причинам он окрашивает лишь небольшой процент клеток в ткани, обнажая полную микроструктуру отдельных нейронов без значительного перекрытия с другими клетками.
Доктрина нейронов
Рисование нейронов в мозжечке голубя , испанского нейробиолога Сантьяго Рамона и Кахала в 1899 году. (A) обозначает клетки Пуркинье и (B) обозначает гранулярные клетки, обе из которых являются мультиполярными.
Доктрина нейронов — это теперь фундаментальная идея о том, что нейроны являются основными структурными и функциональными единицами. нервной системы. Теория была выдвинута Сантьяго Рамоном-и-Кахалем в конце 19 века. Он считал, что нейроны — это отдельные клетки (не связанные в сеть), действующие как метаболически отдельные единицы.
Более поздние открытия привели к уточнению доктрины. Например, глиальные клетки, которые не считаются нейронами, играют важную роль в обработке информации. Кроме того, электрические синапсы встречаются чаще, чем считалось ранее, они представляют собой прямые цитоплазматические связи между нейронами. Фактически, нейроны могут образовывать еще более тесные связи: гигантский аксон кальмара возникает в результате слияния нескольких аксонов.
Рамон-и-Кахаль также постулировал закон динамической поляризации, который гласит, что нейрон получает сигналы на своих дендритах и тело клетки и передает их в виде потенциалов действия по аксону в одном направлении: от тела клетки. Закон динамической поляризации имеет важные исключения; дендриты могут служить в качестве участков синаптического выхода нейронов, а аксоны могут получать синаптические входы.
Компартментная модель нейронов
Хотя нейроны часто называют «фундаментальные единицы» мозга, они выполняют внутренние вычисления. Нейроны интегрируют входные данные в дендриты, и эта сложность теряется в моделях, которые предполагают, что нейроны являются фундаментальной единицей. Дендритные ветви можно моделировать как пространственные компартменты, активность которых связана из-за пассивных свойств мембраны, но также может быть различной в зависимости от входных сигналов от синапсов. Компартментное моделирование дендритов особенно полезно для понимания поведения нейронов, которые слишком малы для регистрации с помощью электродов, как в случае с Drosophila melanogaster.
Нейроны в головном мозге
Число нейронов в головном мозге сильно варьируется от вида к виду. У человека примерно 10–20 миллиардов нейронов в коре головного мозга и 55–70 миллиардов нейронов в мозжечке. Напротив, нематода червь Caenorhabditis elegans имеет всего 302 нейрона, что делает его идеальным модельным организмом, поскольку ученые смогли отобразить все его нейроны. Плодовая мушка Drosophila melanogaster, обычный объект биологических экспериментов, имеет около 100 000 нейронов и демонстрирует множество сложных форм поведения. Многие свойства нейронов, от типа используемых нейромедиаторов до состава ионных каналов, сохраняются у разных видов, что позволяет ученым изучать процессы, происходящие в более сложных организмах, в гораздо более простых экспериментальных системах.
Неврологические расстройства
Болезнь Шарко – Мари – Тута (CMT) — это гетерогенное наследственное заболевание нервов (невропатия ), которое характеризуется потерей мышечной ткани и ощущение прикосновения, преимущественно в ступнях и ногах, распространяющееся на кисти и руки на поздних стадиях. В настоящее время неизлечимое, это заболевание является одним из наиболее распространенных наследственных неврологических расстройств, которым страдают 36 из 100 000.
Болезнь Альцгеймера (AD), также известная просто как болезнь Альцгеймера, является нейродегенеративным заболеванием характеризуется прогрессирующим когнитивным ухудшением, вместе со снижением активности повседневной жизни и психоневрологическими симптомами или изменениями поведения. Наиболее ярким ранним симптомом является потеря кратковременной памяти (амнезия ), которая обычно проявляется в виде незначительной забывчивости, которая становится все более выраженной по мере прогрессирования болезни, с относительным сохранением старых воспоминаний. По мере прогрессирования расстройства когнитивные (интеллектуальные) нарушения распространяются на области языка (афазия ), умелых движений (апраксия ) и распознавания (агнозия ), и такие функции, как принятие решений и планирование, становятся нарушенными.
Болезнь Паркинсона (БП), также известная как болезнь Паркинсона, является дегенеративным заболеванием центральной нервной системы, которое часто ухудшает моторику и речь. Болезнь Паркинсона относится к группе состояний, называемых двигательными расстройствами. Он характеризуется ригидностью мышц, тремором, замедлением физических движений (брадикинезия ) и, в крайних случаях, потерей физических движений (акинезия ). Первичные симптомы являются результатом пониженной стимуляции моторной коры базальными ганглиями, обычно вызванной недостаточным образованием и действием дофамина, который вырабатывается дофаминергическими нейронами головной мозг. Вторичные симптомы могут включать высокий уровень когнитивной дисфункции и тонкие языковые проблемы. БП бывает хроническим и прогрессирующим.
Myasthenia gravis — нервно-мышечное заболевание, приводящее к неустойчивой мышечной слабости и утомляемости во время простых действий. Слабость обычно вызывается циркулирующими антителами, которые блокируют рецепторы ацетилхолина в постсинаптическом нервно-мышечном соединении, подавляя стимулирующий эффект нейротрансмиттера ацетилхолина. Миастения лечится иммунодепрессантами, ингибиторами холинэстеразы и, в отдельных случаях, тимэктомией.
демиелинизацией
синдромом Гийена-Барре — демиелинизацией
демиелинизацией это акт демиелинизации, или потеря миелиновой оболочки, изолирующей нервы. Когда миелин разлагается, передача сигналов по нерву может быть нарушена или потеряна, и в конечном итоге нерв увядает. Это приводит к определенным нейродегенеративным расстройствам, таким как рассеянный склероз и хроническая воспалительная демиелинизирующая полинейропатия.
Дегенерация аксонов
Хотя большинство реакций на травмы включают передачу сигналов притока кальция, способствующую повторному закрытию отрубленных частей, травмы аксонов первоначально приводят к острой дегенерации аксонов, то есть быстрому разделению проксимального и дистального концов в течение 30 минут после травмы. Дегенерация следует за набуханием аксолеммы и в конечном итоге приводит к образованию бусинок. Гранулярный распад аксонального цитоскелета и внутренних органелл происходит после деградации аксолеммы. Ранние изменения включают накопление митохондрий в паранодальных областях в месте повреждения. Эндоплазматический ретикулум разрушается, митохондрии набухают и в конечном итоге распадаются. Распад зависит от протеаз убиквитина и кальпаина (вызывается притоком иона кальция), что позволяет предположить, что дегенерация аксонов является активным процессом, который вызывает полную фрагментацию. Этот процесс занимает около 24 часов в ПНС и дольше в ЦНС. Сигнальные пути, ведущие к дегенерации аксолеммы, неизвестны.
Нейрогенез
Нейроны рождаются в процессе нейрогенеза, в котором нервные стволовые клетки делятся с образованием дифференцированных нейронов. После того как сформированы полностью дифференцированные нейроны, они больше не могут подвергаться митозу. Нейрогенез в основном происходит в эмбрионе большинства организмов.
Нейрогенез может происходить в мозге взрослого позвоночного, открытие, которое вызвало споры в 1999 году. Более поздние исследования возраста человеческих нейронов показывают, что этот процесс происходит только для меньшинства клеток, и подавляющее большинство нейронов, составляющих неокортекс, формируется до рождения и сохраняется без замены. Степень, в которой нейрогенез взрослых людей существует у людей, и его вклад в познание являются спорными, и в 2018 году были опубликованы противоречивые отчеты.
Тело содержит различные типы стволовых клеток, которые способны дифференцироваться в нейроны. Исследователи нашли способ трансформировать клетки кожи человека в нервные клетки с помощью трансдифференцировки, при которой «клетки вынуждены принимать новые идентичности».
Во время нейрогенеза у млекопитающих мозг, клетки-предшественники и стволовые клетки прогрессируют от пролиферативных делений к дифференцированным. Эта прогрессия приводит к нейронам и глии, которые населяют корковые слои. Эпигенетические модификации играют ключевую роль в регуляции экспрессии гена в дифференцировке нервных стволовых клеток и имеют решающее значение для определения судьбы клеток в развивающемся и взрослом мозге млекопитающих. Эпигенетические модификации включают метилирование цитозина ДНК с образованием 5-метилцитозина и деметилирование 5-метилцитозина. Эти модификации имеют решающее значение для определения судьбы клеток в мозге развивающихся и взрослых млекопитающих. Метилирование цитозина ДНК катализируется ДНК-метилтрансферазами (DNMT). Деметилирование метилцитозина в несколько стадий катализируется ферментами TET, которые проводят окислительные реакции (например, 5-метилцитозин в 5-гидроксиметилцитозин ) и ферментами ДНК Путь эксцизионной репарации оснований (BER).
На разных стадиях развития нервной системы млекопитающих используются два процесса репарации ДНК при репарации двухцепочечных разрывов ДНК. Эти пути представляют собой гомологичную рекомбинационную репарацию, используемую в пролиферирующих клетках-предшественниках нервных клеток, и негомологичное соединение концов, используемые в основном на более поздних стадиях развития
регенерация нервов
Периферические аксоны могут вырасти заново, если они разорваны, но один нейрон не может быть функционально заменен другим типом (закон Ллинаса ).
См. Также
- Искусственный нейрон
- Двунаправленная клетка
- Модель биологического нейрона
- Клетка Догеля
- Список животных по количеству нейронов
- Список баз данных нейробиологии
- Нейронный гальванотропизм
- Нейропластичность
- Конус роста
- Анализ Шолля
Ссылки
Дополнительная литература
Внешние ссылки
- Нейробиология на Керли
- IBRO (Международная организация исследования мозга). Содействие исследованиям в области нейробиологии, особенно в странах с менее обеспеченным финансированием.
- NeuronBank онлайн-инструмент для нейромики для каталогизации типов нейронов и синаптических связей.
- Нейроанатомический анализ высокого разрешения I Маги мозга приматов и не-приматов.
- Департамент нейробиологии в Викиверситет, который в настоящее время предлагает два курса: Основы нейробиологии и Сравнительный Нейробиология.
- Поиск NIF — Нейрон через Информационную структуру нейробиологии
- Центрированная база данных — Нейрон
- Полный список типов нейронов в соответствии с соглашением Петиллы, в NeuroLex.
- NeuroMorpho.Org онлайн-база данных цифровых реконструкций морфологии нейронов.
- Иммуногистохимия Галерея изображений: Нейрон
- Академия Хана: Анатомия нейрона
- Изображения нейрона
Строение
Общее строение нейрона выглядит следующим образом: есть тело (сома), в котором содержатся ядро и другие органоиды, и отростки — аксон и дендриты:
- Аксон присутствует всего один — это отросток, по которому нервный импульс идет от данной клетки к другим. Другими словами, аксон — канал выхода сигнала.
- Дендриты, соответственно, — каналы входа сигналов, и их может быть как очень много, так и совсем мало. Количество дендритов зависит от типа нейрона, и об этом мы поговорим позже.
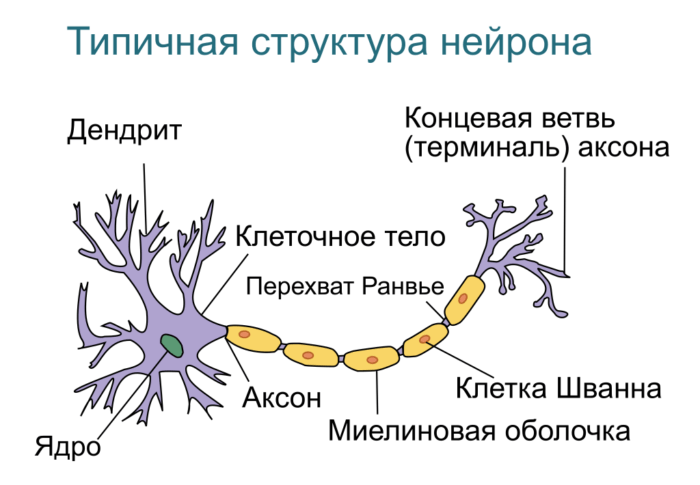
Аксоны и дендриты
Аксоны — отростки, которые могут достигать в длину более метра. Чтобы сигнал не “рассеивался” по пути от одной клетки к другой, большинство аксонов в теле покрыты миелиновой оболочкой, состоящей из клеток нейроглии (общее обозначение вспомогательных клеток нервной ткани). Оболочка обеспечивает изоляцию одного аксона от других и не позволяет электрическому импульсу рассеяться. Благодаря миелиновой оболочке, проведение импульса по аксону осуществляется быстрее. Дендриты более короткие и не покрыты миелином.
Эндоплазматическая сеть (ЭПС) и комплекс Гольджи
Наиболее важными органоидами, помимо ядра, являются шероховатая ЭПС, имеющая рибосомы и осуществляющая синтез белков, и аппарат Гольджи, синтезирующий различные органические вещества и “упаковывающий” их в мембранные пузырьки. Почему эти системы так важны для функциональной деятельности нейрона — будет понятно далее.
Типы нейронов
А сейчас разберем, какие бывают типы по количеству отростков, и в чем их особенности:
- Униполярные имеют только один аксон. Это — чувствительные клетки, в которых возникает возбуждение, и они проводят его далее по аксону.
- Псевдоуниполярные. От тела отходит один небольшой “хвостик”, но он ветвится на два — один из них представляет собой аксон, а другой — дендрит. Такие нейроны находятся, в частности, в нервных узлах спинного мозга.
- Биполярные. Как следует из названия, отростка у них два — аксон и дендрит. Их можно найти в органах чувств, например, на сетчатке глаза.
- Мультиполярные. Имеют один аксон и множество дендритов, расходящихся вокруг подобно щупальцам. Как правило, именно так изображают эти клетки в учебниках, и таких клеток большинство.
Важно! Мультиполярные — преобладают в центральной нервной системе (ЦНС). К ним поступают сигналы от множества соседних нейронов — каждый мультиполярный нейрон может быть связан с 1000 других!
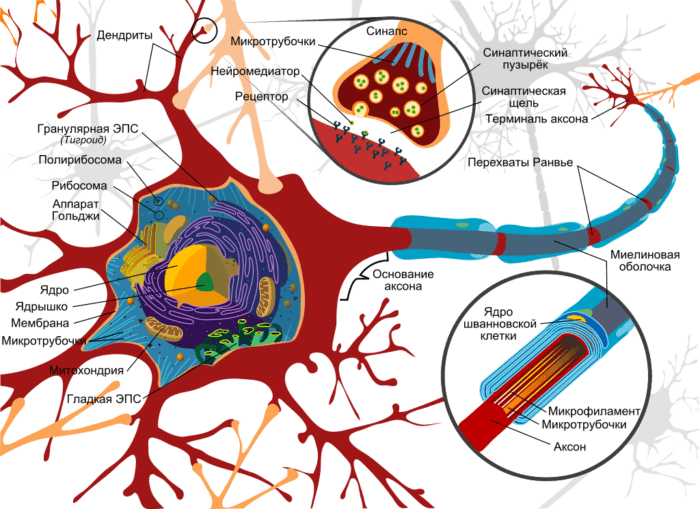
Функции нейронов
Какие функции есть у нервной ткани? Нервная система, наряду с эндокринной, осуществляет координацию деятельности всего организма. Каждый нейрон является частью цепи в координации того или иного физиологического (или же психического) процесса. Говоря вообще, основная функция нейрона заключается в получении и передаче информации. Это справедливо в отношении любой клетки рассматриваемой системы, ведь именно этим она и занимается — получает от одних клеток и передает другим информацию в форме нервных импульсов. Однако для различных нейронов выделяют и более специфические функции. Виды нейронов по функциям:
- Афферентные (чувствительные): получают информацию непосредственно от рецепторов, осуществляя взаимодействие между внешним миром и нашей нервной системой.
- Эфферентные (двигательные): отвечают за осуществление конкретных действий — сокращение мышцы, выделение секрета железой.
- Ассоциативные (вставочные): это все “средние” нейроны в цепочке, их может не быть вовсе или быть несколько в одной рефлекторной дуге. Они сосредоточены в ЦНС, отвечают за обработку информации и, говоря грубо, принятие нервной системой решений о действии организма.
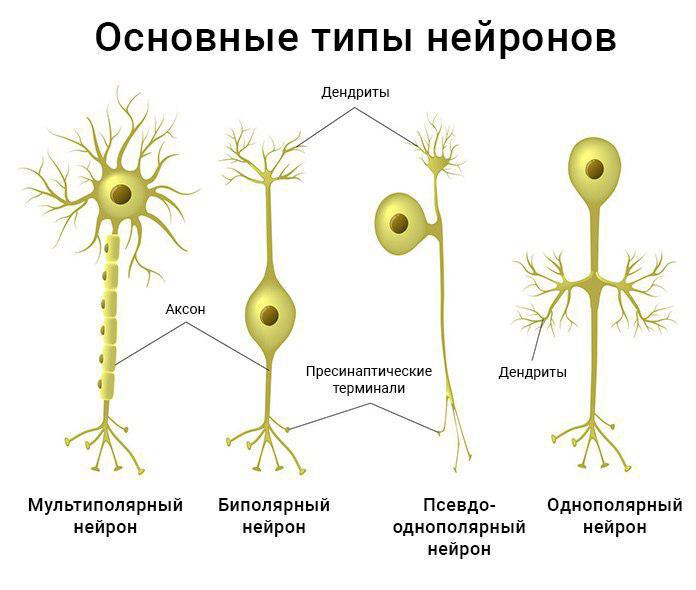
Передача нервного импульса
Каким образом импульс передается между клетками? Этот вопрос интереснее, чем кажется на первый взгляд. Сейчас мы поймем, зачем же нейронам сильно развитые ЭПС и аппарат Гольджи. Место контакта аксона с дендритом, телом другого нейрона или эффектором называется синапс. Помимо связи между клетками, синапс также осуществляет перекодировку сигнала, меняя различные его характеристики (частоту, амплитуду). Рассмотрим подробнее схему работы синаптической передачи нервного импульса:
- Электрический нервный импульс возбуждает мембрану нейрона. Наиболее возбудимая часть на теле таких клеток — аксонный холмик, с него и начинается общее возбуждение клетки. Оно передается далее по мембране аксона и достигает его конца — пресинаптической мембраны.
- Внутри аксона содержатся пузырьки с нейромедиаторами — биологически активными органическими веществами. Помните про ЭПС и аппарат Гольджи? Именно эти системы отвечают за синтез и транспорт веществ-медиаторов к аксонному окончанию.
- Синаптическая щель — пространство между аксоном этой клетки и дендритом следующей. Под действием электрического возбуждения пресинаптическая мембрана высвобождает медиаторы в пространство синаптической щели. Там они связываются с соответствующими рецепторами на постсинаптической мембране, запуская тем самым возбуждение следующего звена нервной цепочки.
Важно! На уровне нейро-физиологии эти процессы выглядят достаточно сложно, и для глубокого понимания требуются знания не только биологии, но и химии, и физики. Однако основное, что нужно понимать — есть две мембраны, связь между которыми осуществляют медиаторы (буквальный перевод — посредники). От того, какой медиатор выделяется, зависит эффект — возбуждение или торможение следующей клетки.
Подведем итоги: нейроны — клетки нервной системы. Их слаженная работа обеспечивает координацию всех функций организма, начиная от движения и работы внутренних органов и заканчивая высшими психическими процессами. Нейроны передают информацию посредством электрических импульсов, которые проходят по аксону одной клетки и при помощи нейромедиаторов в синапсе передаются другой клетке. Такой с виду простой механизм лежит в основе работы нервной системы. Лучше понять работу всей нервной системы вам также поможет предложенное ниже видео.