Содержание
- История
- Строение нейронов
- Тело клетки
- Дендриты и аксон
- Синапс
- Сложная обёртка
- Свойства нервной ткани
- Как формируется аксон
- Нейроны: строение, виды и типы
- Классификация
- Структурная классификация
- Функциональная классификация
- Морфологическая классификация
- Классификация
- Последствия разрушения дендритов
- Узелки Ранвье.
- Особенности строения и функции аксонов и дендритов.
- Строение нейрона[править | править код]
История
Немецкому анатому Отто Фридриху Карлу Дейтерсу обычно приписывают открытие аксона, отличив его от дендритов. Швейцарский Рюдольф Альберт фон Келликер и немец Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Келликер назвал аксон в 1896 году. Луи-Антуан Ранвье был первым, кто описал промежутки или узлы, обнаруженные на аксонах, и за этот вклад эти аксональные особенности теперь обычно называют узлами Ранвье . Сантьяго Рамон-и-Кахаль , испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции. Джозеф Эрлангер и Герберт Гассер ранее разработали систему классификации периферических нервных волокон, основанную на скорости проводимости аксонов, миелинизации , размере волокна и т. Д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), и к 1952 году они получили полную количественную оценку. описание ионной основы потенциала действия , приведшее к формулировке модели Ходжкина – Хаксли . В 1963 году Ходжкин и Хаксли были совместно удостоены Нобелевской премии за эту работу. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхейзера-Хаксли. Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает много деталей об отдельных ионных каналах .
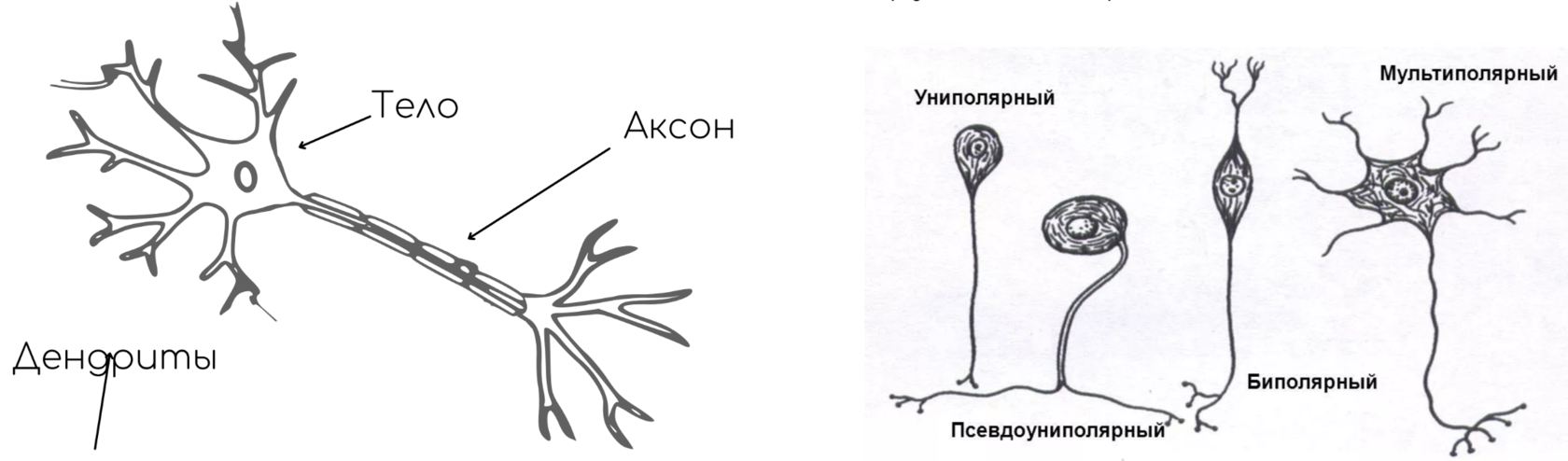
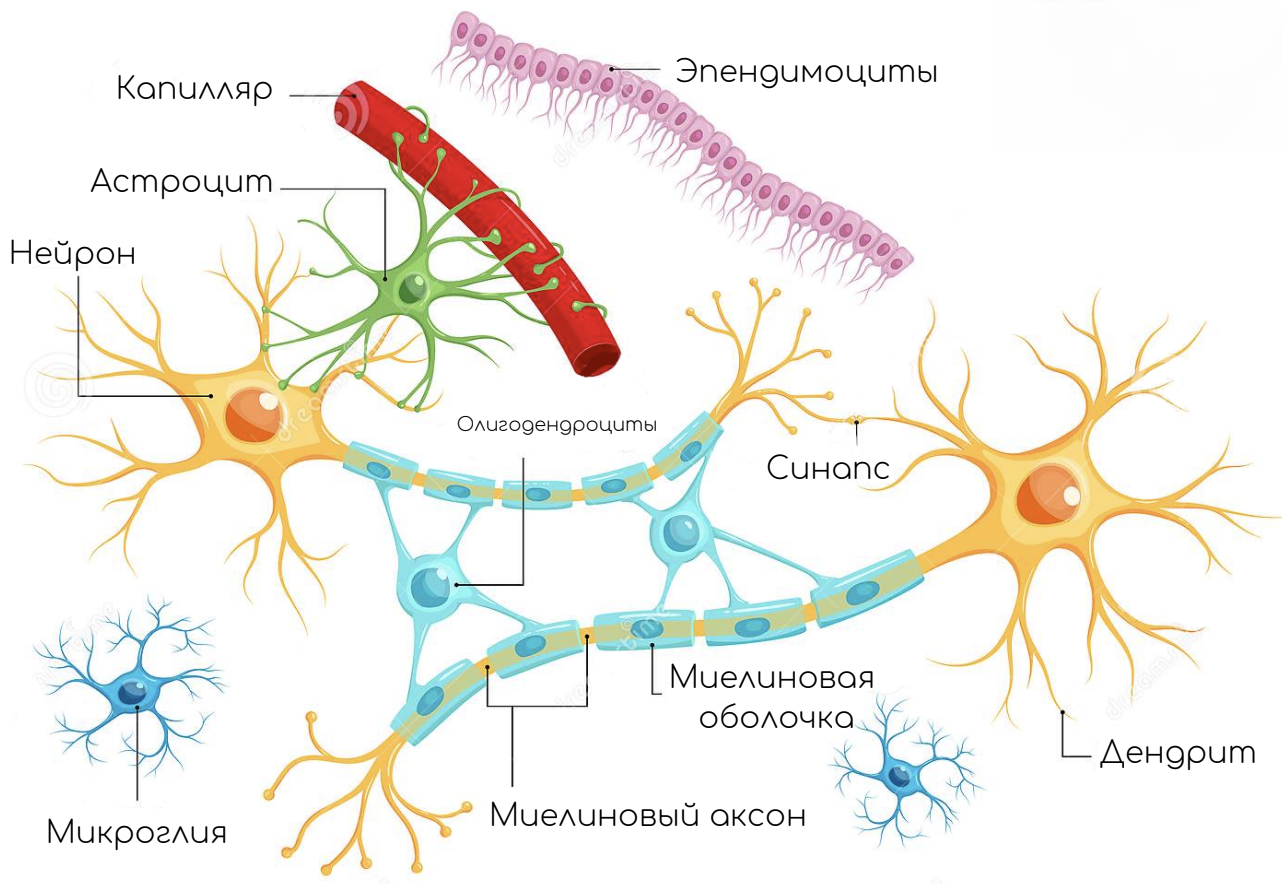
Строение нейронов
Схема нейрона
Тело клетки
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), ограниченной снаружи мембраной из липидного бислоя. Липиды состоят из гидрофильных головок и гидрофобных хвостов. Липиды располагаются гидрофобными хвостами друг к другу, образуя гидрофобный слой. Этот слой пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: в форме глобул на поверхности, на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20—30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии.(Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов).
В теле нейрона выявляется развитый синтетический аппарат, гранулярная эндоплазматическая сеть нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Основные статьи: Дендрит и Аксон
Схема строения нейрона
Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу.
Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом для образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Основная статья: Синапс
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г.
Сложная обёртка
Миелин окружает отростки нервных клеток, изолируя их от внешнего воздействия. Это необходимо для более надежной и быстрой передачи сигнала по нервной системе. Благодаря изоляции нервного волокна электрический сигнал не рассеивается и добирается до места назначения без помех. Скорость прохождения сигнала по миелиновым и безмиелиновым волокнам может отличаться на три порядка: от 70 до 140 м/с и от 0,3 до 0,5 м/с соответственно.
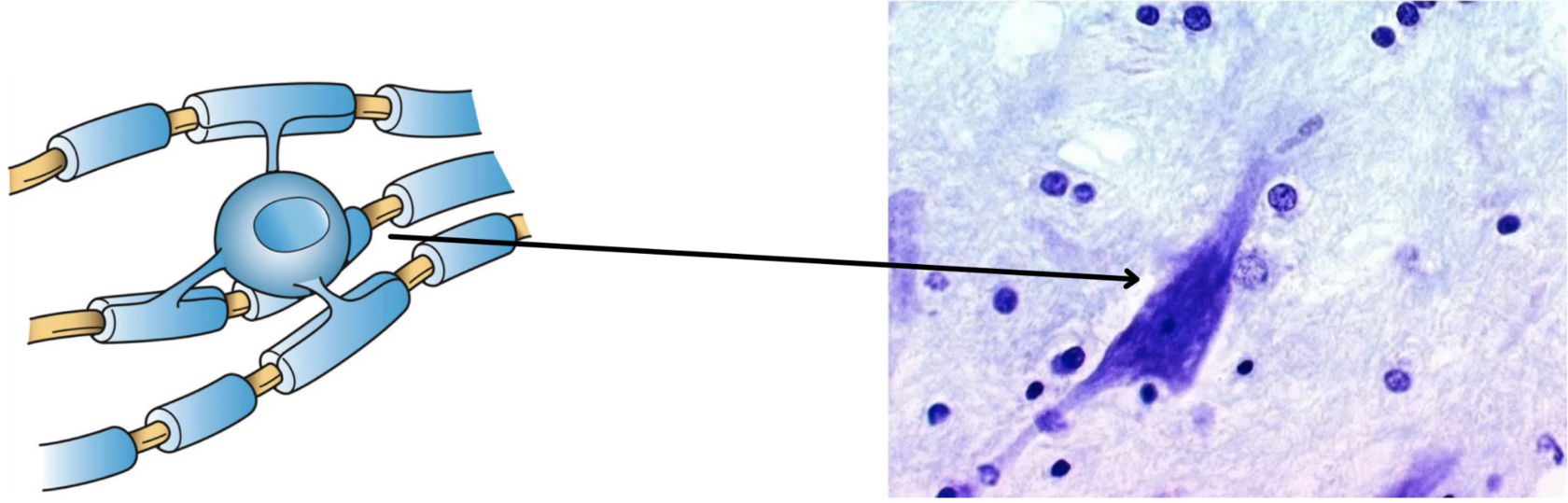
По сути миелин — это клеточная мембрана глиальных клеток, многократно обмотанная вокруг аксона. Сама мембрана на 70–75% состоит из липидов и на 25–30% — из белков. В периферической нервной системе донором мембран становятся шванновские клетки, а в центральной — олигодендроциты. Эти клетки бережно обматывают своими мембранами ценные каналы связи, чтобы обеспечить надежное взаимодействие нервной системы и периферических органов. Миелин покрывает нервное волокно не целиком: существуют промежутки между наслоениями миелина, называемые перехватами Ранвье (рис. 1). Есть прямая зависимость между расстоянием от одного промежутка до другого и скоростью распространения нервного импульса по волокну: чем больше расстояние между перехватами Ранвье, тем выше скорость передачи сигнала в нерве .
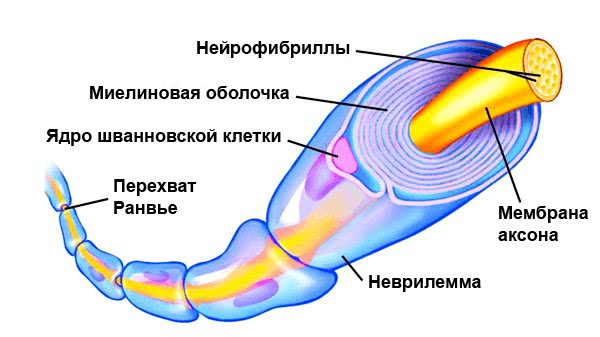
Рисунок 1. Нервное волокно, обернутое миелином. Видны ядра шванновских клеток (nucleus of Schwann cell) и перехваты Ранвье (nodes of Ranvier) — участки аксона, которые не покрыты миелиновой оболочкой.
сайт theclickercenterblog.com
Если говорить о белках, входящих в состав миелина, то надо уточнить, что это не только простые белки. В миелине встречаются гликопротеины — белки, к которым присоединены короткие углеводные последовательности
Важной составляющей миелина является главный структурный белок миелина (myelin basic protein, MBP), впервые выделенный около 50 лет назад. MBP — это трансмембранный белок, который может многократно «прошивать» липидный слой клетки
Его различные изоформы (рис. 2) кодируются геном под названием Golli (gene in the oligodendrocyte lineage). Структурной основой миелина служит изоформа массой 18,5 килодальтон .
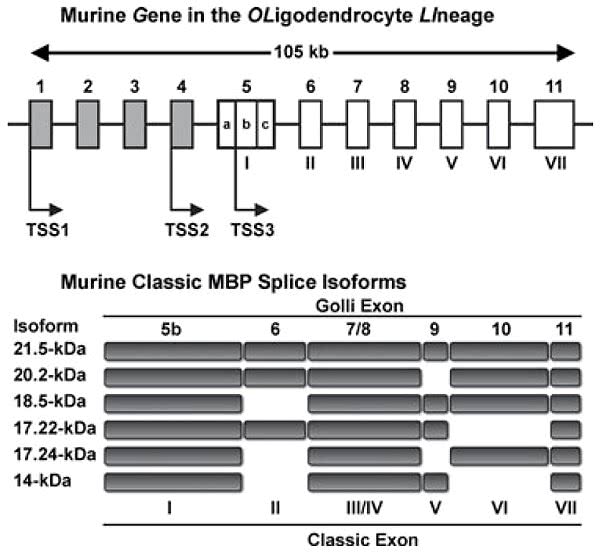
Рисунок 2. Различные изоформы основного белка миелина (MBP) создаются на основе одного и того же гена. Например, для синтеза изоформы массой 18,5 кДа используются все экзоны, кроме экзона II.
В состав миелина входят сложные липиды цереброзиды. Они представляют собой аминоспирт сфингозин, соединенный с жирной кислотой и остатком углевода. В синтезе липидов миелина принимают участие пероксисомы олигодендроцитов. Пероксисомы — это липидные пузырьки с различными ферментами (в общей сложности известно около 50 видов пероксисомных энзимов). Эти органеллы занимаются, в частности, β-окислением жирных кислот: жирных кислот с очень длинной цепью (very long chain fatty acids, VLCFA), некоторых эйкозаноидов и полиненасыщенных жирных кислот (ПНЖК, polyunsaturated fatty acids, PUFAs). Поскольку миелин может содержать до 70% липидов, пероксисомы крайне важны для нормального метаболизма этого вещества. Они используют N-ацетиласпартат, вырабатываемый нервной клеткой, для постоянного синтеза новых липидов миелина и поддержания его существования. Кроме этого, пероксисомы принимают участие в поддержании энергетического метаболизма аксонов .
Свойства нервной ткани
Возбудимость и проводимость — характерные особенности нейронов. Информация передается по отросткам в виде электрических импульсов возбуждения (рис. 4). Это быстрые и кратковременные изменения электрического заряда наружной клеточной мембраны.
Рис. 4. Передача возбуждения в нейронах
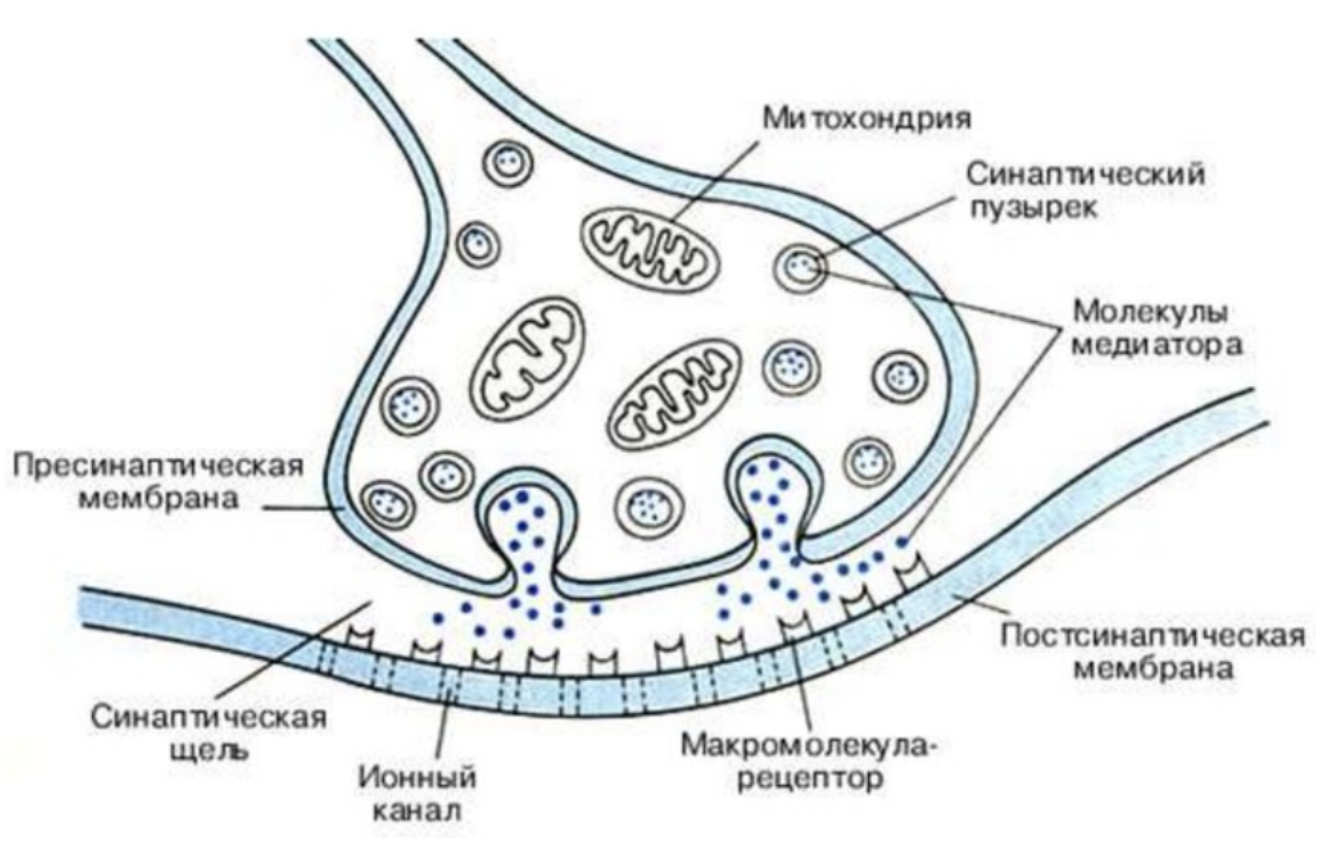
Передача информации от нейрона к нейрону происходит в синапсах — местах сближения клеток (нейронов друг с другом или с клетками других тканей). Процесс осуществляется с помощью физиологически активных веществ. Они получили названия «медиаторы» или «нейротрансмиттеры». Медиатор (гистамин, ацетилхолин, дофамин) содержится в специальных пузырьках в окончании аксона.
Части синапса:
- Аксон.
- Пресинаптическая мембрана.
- Синаптический пузырек.
- Синаптическая щель.
- Постсинаптическая мембрана.
- Рецепторы для медиатора.
При возбуждении нейрона импульс достигает окончания аксона. Медиатор выходит из пузырьков и передается через синаптическую щель аксону (дендриту, телу другой нервной клетки или другим клеткам организма). В этих соседних клетках возникает возбуждение или торможение.
Пучки аксонов в изолирующей оболочке образуют нервы. По этим волокнам распространяются нервные импульсы. Передача сигналов происходит только в одном направлении благодаря асимметричной конструкции синапса.
Нервная ткань способна выполнять сложные функции благодаря особому строению нервных клеток и наличию вспомогательных элементов, образующих нейроглию. Основные свойства ткани — раздражимость и возбудимость.
Как формируется аксон
Удлинение и развитие данных отростков нейрона обеспечивается локацией их расположения. Удлинение аксонов становится возможным по причине присутствия на их верхнем завершении филоподий, между которыми размещаются, подобие гофры, мембранные образования — ламелоподии. Филоподий деятельно взаимодействуют с близлежащими структурами, пробиваясь в ткань все глубже, по итогам чего и осуществляется направленное удлинение аксонов. Собственно филоподий задает направление увеличению аксона в длину, устанавливая, определенность организации волокон. Участие филоподий в направленном удлинении нейтритов было подтверждено в практическом эксперименте посредством введения в эмбрионы, цитохалазина В, разрушающего филоподий. При этом аксоны нейронов не дорастали до мозговых центров.
Продуцирование иммуноглобулина, который часто встречается в месте соединения участков роста аксонов с глиальными клетками и, согласно гипотезам ряда ученых, данный факт и предопределяет направление удлинения аксонов в зоне перекреста. Если данный фактор способствует удлинению аксонов, то хондроитинсульфат, напротив, замедляет рост нейтритов.
Нейроны: строение, виды и типы
Нервная ткань содержит клетки нервные клетки и нейроглию (рис. 1). Ткань образует головной и спинной мозг, нервные волокна и узлы. Нервная система отвечает за согласованную работу органов и систем органов, обеспечивает связь организма с окружающей средой.
Рис. 1. Нервные клетки сетчатки лошади
Нейрон — основная, высокоспециализированная клетка нервной ткани. Она осуществляет прием, обработку и передачу информации. Состоит из тела или сомы, в котором заключены ядро с основной массой цитоплазмы, и отростков. Диаметр тела нервной клетки составляет 15–150 мк или 0,001 мм.
Виды нейронов по количеству отростков (рис. 2):
- биполярные;
- униполярные;
- мультиполярные;
- псевдоуниполярные.
Рис. 2. Виды нейронов
Тела нейронов сконцентрированы, главным образом, в сером веществе головного и спинного мозга. Длинные отростки тянутся на большие расстояния от места, где находятся нервные клетки с ядром. Длина аксона может достигать 1 м и более.
Составные части двигательного (мультиполярного) нейрона (рис. 3):
- Тело нервной клетки с расположенным в центре ядром.
- Короткие ветвящиеся отростки дендриты, несущие информацию к телу клетки.
- Длинный клеточный отросток аксон, несущий информацию от тела нейрона.
- Изолирующая миелиновая оболочка аксона из шванновских клеток (входят в состав нейроглии).
- Перехваты Ранвье — узкие промежутки, разделяющие шванновские клетки.
- Чувствительные окончания — рецепторы.
Рис. 3. Строение нейрона
Типы нейронов в зависимости от выполняемой функции
|
Основное название |
Дополнительные названия |
Функции |
|
Чувствительные |
Афферентные, сенсорные |
Проводят информацию об ощущении (импульс) от поверхности тела и внутренних органов в мозг. |
|
Вставочные |
Ассоциативные, связывающие, переключающие |
Составляют около 99% всех нервных клеток, обрабатывают, анализируют информацию, вырабатывают решения. |
|
Двигательные |
Эффекторные, эфферентные |
Проводят импульс от головного и спинного мозга к исполнительным органам. |
Классификация
Структурная классификация
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны
— небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны
— нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Многие морфологи считают, что униполярные нейроны в теле человека и высших позвоночных не встречаются.
Биполярные нейроны
— нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны
— нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны
— являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны
(чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны
(эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны
(вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными.
Секреторные нейроны
— нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация
Морфологическое строение нейронов многообразно. При классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона;
- количество и характер ветвления отростков;
- длину аксона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов.
По количеству отростков выделяют следующие морфологические типы нейронов:
- униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
- псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
- биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
- мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Классификация
Нейроны разделяют на виды в зависимости от типа медиатора (посредника проводящего импульса) выделяемого на окончаниях аксона. Это может быть холин, адреналин и пр. От места расположения в отделах ЦНС они могут относиться к соматическим нейронам или к вегетативным. Различают воспринимающие клетки (афферентные) и передающие обратные сигналы (эфферентные) в ответ на раздражение. Между ними могут находиться итернейроны, отвечающие за обмен информацией внутри ЦНС. По типу ответной реакции клетки могут тормозить возбуждение или, наоборот, повышать его.
По состоянию их готовности различают: «молчащие», которые начинают действовать (передают импульс) только при наличии определенного вида раздражения, и фоновые, что постоянно осуществляют мониторинг (непрерывная генерация сигналов). В зависимости от типа воспринимаемой от сенсоров информации меняется и строение нейрона. В этой связи их классифицируют на бимодальные, с относительно простым ответом на раздражение (два взаимосвязанных вида ощущения: укол и — как результат — боль, и полимодальные. Это более сложная структура – полимодальные нейроны (специфическая и неоднозначная реакция).
Последствия разрушения дендритов
Они хоть и после устранения условий, вызвавших нарушения в их построении, способны восстанавливаться, полностью нормализуя обмен веществ, но только если эти факторы недолго, незначительно воздействовали на нейрон, в противном же случае, части дендритов погибают, и так как не имеют возможности покинуть организм, накапливаются в их цитоплазме, провоцируя отрицательные последствия.
У животных это приводит к нарушению форм поведения, за исключением простейших условных рефлексов, а у человека может вызвать нарушения нервной системы.
Кроме того, рядом ученных доказано, что при слабоумии в пожилом возрасте и заболевание Альцгеймера у нейронов не отслеживаются отростки. Стволы дендритов внешне становятся похожи на обгоревшие (обугленные).
Не менее важным является и изменения количественного эквивалента шипиков вследствие патогенных условий. Так как они признаны структурными компонентами межнейрональных контактов, то нарушения, возникающие в них, могут спровоцировать достаточно серьезные нарушениям функций мозговой деятельности.
Узелки Ранвье.
Миелиновая оболочка нейронов не непрерывна по всему аксону. Фактически миелин образует «пакеты», которые немного отделены друг от друга. И это разделение, длина которого меньше микрометра, и есть то, что называется узелком Ранвье.
Следовательно, узлы Ранвье — это небольшие области аксона, которые не окружены миелином и открывают его во внеклеточное пространство. Они необходимы для правильной передачи электрического импульса, поскольку через них проходят электролиты натрия и калия, которые жизненно важны для правильного (и с более высокой скоростью) прохождения электрического сигнала через аксон.
Особенности строения и функции аксонов и дендритов.
Аксон (греч. ἀξον — ось) — нейрит, осевой цилиндр, отросток нервной клетки, по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам. В месте соединения аксона с телом нейрона находится аксонный холмик. Регистрация электрических потенциалов выявила, что нервный импульс генерируется в самом аксоне, а именно в начальном сегменте. Питание и рост аксона зависят от тела нейрона: при перерезке аксона его периферическая часть отмирает, а центральная сохраняет жизнеспособность. При диаметре в несколько микронов длина аксона может достигать у крупных животных 1 метра и более (например, аксоны, идущие от нейронов спинного мозга в конечности). У многих животных встречаются гигантские аксоны толщиной в сотни мкм (у кальмаров — до 2—3 мм). Обычно такие аксоны отвечают за проведение сигналов к мышцам.обеспечивающим «реакцию бегства» (втягивание в норку, быстрое плавание и др.). При прочих равных условиях с увеличением диаметра аксона увеличивается скорость проведения по нему нервных импульсов.
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна. Миелиновая оболочка аксонов имеется только у позвоночных. Её образуют «накручивающиеся» на аксон специальные шванновские клетки, между которыми остаются свободные от миелиновой оболочки участки — перехваты Ранвье. Только на перехватах присутствуют потенциал-зависимые натриевые каналы и заново возникает потенциал действия. При этом нервный импульс распространяется по миелинизированным волокнам ступенчато, что в несколько раз повышает скорость его распространения. Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью.
Дендрит.
Основные характерные черты дендрита, которые выделяют его на электронно-микроскопических срезах:
§ отсутствие миелиновой оболочки
§ наличие правильной системы микротрубочек
§ наличие на них активных зон синапсов с ясно выраженной электронной плотностью цитоплазмы дендрита
§ отхождение от общего ствола дендрита шипиков,
§ специально организованные зоны узлов ветвлений,
§ вкрапление рибосом
§ наличие в проксимальных участках гранулированного и не гранулированного ЭПР.
Наиболее примечательной особенностью цитоплазмы дендритов является наличие многочисленных микротрубочек. Они хорошо выявляются как на поперечных срезах, так и на продольных. Микротрубочки следуют в дендрите параллельно друг другу, не соединяясь и не пересекаясь между собой. Шипик является производным дендрита, в нем отсутствуют нейрофиламенты и дендритические трубочки, его цитоплазма содержит грубо или тонко гранулированный матрикс. Еще одной характерной чертой шипика в коре мозга является обязательное присутствие на них синаптических контактов с аксонными окончаниями. Можно отметить своеобразную триаду в цитоплазме шипика: субсинаптическая специализация активных зон — шипиковый аппарат — митохондрии. Можно говорить о том, что цитоплазма дендритического шипика и шипиковый аппарат могут иметь непосредственное отношение к синаптической функции.
Дендритические шипики и концы дендритов также очень чувствительны к действию экстремальных факторов. При любом виде отравления (например, алкогольном, гипоксическом, тяжелыми металлами — свинцом, ртутью и т.д.) меняется количество выявленных шипиков на дендритах клеток коры больших полушарий. По всей вероятности, шипики при этом не исчезают, но у них нарушаются цитоплазматические компоненты, и они хуже импрегнируются солями тяжелых металлов. Так как шипики — один из структурных компонентов обеспечения межнейрональных контактов, то неполадки в них приводят к серьезным нарушениям функции мозга.
Для всех дендритов ЦНС характерно увеличение поверхности за счет многократного дихотомического деления. При этом образуются в зонах деления особые расширительные площадки или узлы ветвления.
Функции дендритов.
· трофическая функция
· передача нервного импульса от одной клетки к другой, также отвечает за частоту и амплитуду сигнала.
Строение нейрона[править | править код]
|
Аксоны
(Свет снизу) |
|---|
|
Рис. A.
Стрелка и пунткирна линия — внешняя пограничная мембрана H — горизонтальные клетки
|
Нейрон состоит из одного аксона (см. Ax рис.A), тела и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные, мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС, участвует в обеспечении тормозных рефлексов).
В месте соединения аксона с телом нейрона находится аксонный холмик — именно здесь происходит преобразование постсинаптического потенциала нейрона в нервные импульсы, для чего требуется совместная работа натриевых, кальциевых и как минимум трех типов калиевых каналов.
Питание и рост аксона зависят от тела нейрона: при перерезке аксона его периферическая часть отмирает, а центральная сохраняет жизнеспособность. При диаметре в несколько микрон длина аксона может достигать у крупных животных 1 метра и более (например, аксоны, идущие от нейронов спинного мозга в конечности). У многих животных (кальмаров, рыб, кольчатых червей, форонид, ракообразных) встречаются гигантские аксоны толщиной в сотни мкм (у кальмаров — до 2-3 мм). Обычно такие аксоны отвечают за проведение сигналов к мышцам. обеспечивающим «реакцию бегства» (втягивание в норку, быстрое плавание и др.). При прочих равных условиях с увеличением диаметра аксона увеличивается скорость проведения по нему нервных импульсов.
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная (гладкая) эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна.
Миелиновая оболочка аксонов имеется только у позвоночных. Её образуют «накручивающиеся» на аксон специальные шванновские клетки, между которыми остаются свободные от миелиновой оболочки участки — перехваты Ранвье. Только на перехватах присутствуют потенциал-зависимые натриевые каналы и заново возникает потенциал действия. При этом нервный импульс распространяется по миелинизированным волокнам ступенчато, что в несколько раз повышает скорость его распространения.
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
-
Особенности строения и функции аксонов, аксонный транспорт.
Аксон
(греч.
ἀξον —
ось) — нейрит, осевой цилиндр, отросток
нервной клетки, по которому нервные
импульсы идут от тела клетки
(сомы) к иннервируемым органам и другим
нервным клеткам.
Нейрон состоит из
одного аксона, тела и нескольких
дендритов,
в зависимости от числа которых нервные
клетки делятся на униполярные, биполярные,
мультиполярные. Передача нервного
импульса происходит от дендритов (или
от тела клетки) к аксону, а затем
сгенерированный потенциал действия от
начального сегмента аксона передается
назад к дендритам . Если аксон в нервной
ткани соединяется с телом следующей
нервной клетки, такой контакт называется
аксо-соматическим, с дендритами —
аксо-дендритический, с другим аксоном —
аксо-аксональный (редкий тип соединения,
встречается в ЦНС).
В месте соединения
аксона с телом нейрона у наиболее крупных
пирамидных клеток 5-ого слоя коры
находится аксонный
холмик.
Ранее предполагалось, что здесь происходит
преобразование постсинаптического
потенциала нейрона в нервные импульсы,
но экспериментальные данные это не
подтвердили. Регистрация электрических
потенциалов выявила, что нервный импульс
генерируется в самом аксоне, а именно
в начальном сегменте на расстоянии ~50
мкм от тела нейрона. Для генерации
потенциала действия в начальном сегменте
аксона требуется повышенная концентрация
натриевых каналов (до ста раз по сравнению
с телом нейрона[3]).
Питание и рост
аксона зависят от тела нейрона: при
перерезке аксона его периферическая
часть отмирает, а центральная сохраняет
жизнеспособность. При диаметре в
несколько микронов
длина аксона может достигать у крупных
животных 1 метра и более (например,
аксоны, идущие от нейронов спинного
мозга
в конечности).
У многих животных (кальмаров,
рыб,
кольчатых
червей,
форонид,
ракообразных)
встречаются гигантские аксоны толщиной
в сотни мкм (у кальмаров — до 2—3 мм).
Обычно такие аксоны отвечают за проведение
сигналов к мышцам. обеспечивающим
«реакцию бегства» (втягивание в норку,
быстрое плавание и др.). При прочих равных
условиях с увеличением диаметра аксона
увеличивается скорость проведения по
нему нервных импульсов.
В протоплазме
аксона — аксоплазме — имеются
тончайшие волоконца — нейрофибриллы,
а также микротрубочки,
митохондрии
и агранулярная (гладкая) эндоплазматическая
сеть.
В зависимости от того, покрыты ли аксоны
миелиновой
(мякотной) оболочкой или лишены её, они
образуют мякотные или безмякотные
нервные
волокна.
Миелиновая оболочка
аксонов имеется только у позвоночных.
Её образуют «накручивающиеся» на аксон
специальные шванновские
клетки,
между которыми остаются свободные от
миелиновой оболочки участки —
перехваты
Ранвье.
Только на перехватах присутствуют
потенциал-зависимые натриевые каналы
и заново возникает потенциал
действия.
При этом нервный импульс распространяется
по миелинизированным волокнам ступенчато,
что в несколько раз повышает скорость
его распространения.
Концевые участки
аксона — терминали — ветвятся и
контактируют с другими нервными,
мышечными или железистыми клетками. На
конце аксона находится синаптическое
окончание —
концевой участок терминали, контактирующий
с клеткой-мишенью. Вместе с постсинаптической
мембраной клетки-мишени синаптическое
окончание образует синапс.
Через синапсы
передаётся возбуждение.
Специфическая
функция аксона — проведение потенциала
действия
от тела клетки к другим клеткам или
периферическим органам. Другая его
функция — аксонный
транспорт
веществ.
Помимо своей
специфической функции в качестве
проводника потенциалов
действия
аксон
является каналом для транспорта веществ.
Аксонный транспорт
— это перемещение веществ по аксону
. Белки, синтезированные в теле клетки,
синаптические
медиаторные вещества
и низкомолекулярные соединения
перемещаются по аксону вместе с клеточными
органеллами, в частности, митохондриями
. Для большинства веществ и органелл
обнаружен также транспорт в обратном
направлении. Вирусы
и токсины
могут проникать в аксон на его периферии
и перемещаться по нему. Аксонный транспорт
— активный процесс.
Аксонный транспорт
зависит от достаточного снабжения
энергией, при снижении уровня АТФ вдвое
аксонный транспорт блокируется, при
возобновлении доступа энергии —
возобновляется.
Белки цитоскелета
доставляются из тела клетки, двигаясь
по аксону со скоростью от 1 до 5 мм в
сутки. Это медленный
аксонный транспорт
(похожий на него транспорт имеется и в
дендритах). Многие ферменты и другие
белки цитозоля также переносятся при
помощи этого типа транспорта.
Нецитозольные
материалы, которые необходимы в синапсе,
такие как секретируемые белки и
мембраносвязанные молекулы, двигаются
по аксону с гораздо большей скоростью.
Эти вещества переносятся из места их
синтеза, эндоплазматического ретикулума,
к аппарату Гольджи, который часто
располагается у основания аксона. Затем
эти молекулы, упакованные в мембранные
пузырьки, переносятся вдоль
рельсов-микротрубочек путем быстрого
аксонного транспорта
со скоростью до 400 мм в сутки. Таким
образом по аксону транспортируются
митохондрии, различные белки, включая
нейропептиды (нейромедиаторы пептидной
природы), непептидные нейромедиаторы.
Транспорт материалов
от тела нейрона к синапсу называется
антероградным,
а в обратном направлении — ретроградным.
Транспорт по аксону
на большие расстояния происходит с
участием микротрубочек. Микротрубочки
в аксоне обладают присущей им полярностью
и ориентированны быстрорастущим
(плюс-)концом к синапсу, а медленнорастущим
(минус-) — к телу нейрона. Белки-моторы
аксонного транспорта принадлежат к
кинезиновому и динеиновому суперсемействам.
Кинезины являются,
в основном, плюс-концевыми моторными
белками, транспортирующими такие грузы,
как предшественники синаптических
везикул и мембранные органеллы. Этот
транспорт идет в направлению к синапсу
(антероградно). Цитоплазматические
динеины — это минус-концевые моторные
белки, транспортирующие нейротрофные
сигналы, эндосомы и другие грузы
ретроградно к телу нейрона. Ретроградный
транспорт осуществляется динеинами не
эксклюзивно: обнаружены несколько
кинезинов, перемещающихся в ретроградном
направлении.
-
Миенилизированные
и немиенилизированные волокна. Процесс
миелинизации.
Миелин
(в некоторых изданиях употребляется
некорректная теперь форма миэлин) —
вещество, образующее миелиновую
оболочку
нервных
волокон.
Миелиновая
оболочка —
электроизолирующая оболочка, покрывающая
аксоны
многих нейронов. Миелиновую оболочку
образуют глиальные клетки: в периферической
нервной системе — Шванновские
клетки,
в центральной нервной системе —
олигодендроциты.
Миелиновая оболочка формируется из
плоского выроста тела глиальной клетки,
многократно оборачивающего аксон
подобно изоляционной ленте. Цитоплазма
в выросте практически отсутствует, в
результате чего миелиновая оболочка
представляет собой, по сути, множество
слоёв клеточной мембраны.
Миелин прерывается
только в области перехватов Ранвье,
которые встречаются через правильные
промежутки длиной примерно 1 мм. В
связи с тем, что ионные токи не могут
проходить сквозь миелин, вход и выход
ионов осуществляется лишь в области
перехватов. Это ведёт к увеличению
скорости проведения нервного импульса.
Таким образом, по миелинизированным
волокнам импульс проводится приблизительно
в 5—10 раз быстрее, чем по немиелинизированным.
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Безмиелиновые
нервные волокна находятся преимущественно
в составе вегетативной нервной системы.
Клетки олигодендроглии оболочек
безмиелиновых нервных волокон,
располагаясь плотно, образуют тяжи, в
которых на определенном расстоянии
друг от друга видны овальные ядра. В
нервных волокнах внутренних органов,
как правило, в таком тяже располагается
не один, а несколько (10-20) осевых цилиндров,
принадлежащих различным нейронам. Они
могут, покидая одно волокно, переходить
в смежное, такие волокна, содержащие
несколько осевых цилиндров, называются
волокнами кабельного типа. При электронной
микроскопии безмиелиновых нервных
волокон видно, что по мере погружения
осевых цилиндров в тяж леммоцитов
последние одевают их как муфтой.
Оболочки леммоцитов
при этом прогибаются, плотно охватывают
осевые цилиндры и, смыкаясь над ними,
образуют глубокие складки, на дне которых
и располагаются отдельные осевые
цилиндры. Сближенные в области складки
участки оболочки нейролеммоцита образуют
сдвоенную мембрану — мезаксон, на которой
как бы подвешен осевой цилиндр. Оболочки
нейролеммоцитов очень тонкие, поэтому
ни мезаксона, ни границ этих клеток под
световым микроскопом нельзя рассмотреть,
и оболочка безмиелиновых нервных волокон
в этих условиях выявляется как однородный
тяж цитоплазмы, «одевающий» осевые
цилиндры. С поверхности каждое нервное
волокно покрыто базальной мембраной.
-
Миелиновые
– более толстые, т.к. содержат миелин-
находятся внутри под оболочкой; входят
в состав соматической нервной системы.
Миелин через несколько участков резко
истончается, образуя таким образом
перехват. Он называется перехват Ранвье.
Отросток между перехватами называется
межузловой сигмент. Миелин на подобии
жира. -
Безмиелиновые-
входят в состав вегетативной нервной
системы, содержат несколько осевых
цилиндров, при этом осевые цилиндры
могут покидать основное волокно и
переходить в смежное.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
ОБЩАЯ ГИТОЛОГИЯ — НЕРВНАЯ ТКАНЬ
Общая информация
Нервная ткань – это система взаимосвязанных нервных клеток и нейроглии,
обеспечивающих специфические функции восприятия раздражений,
возбуждения, выработки импульса и его передачи. Она является основой
строения органов нервной системы, обеспечивающих регуляцию всех тканей
и органов, их интеграцию в организме и связь с окружающей средой.
Типы клеток
- Нервные клетки
Основные структурные компоненты нервной ткани, выполняющие специфическую функцию
- Глиальные клетки
Обеспечивают существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции
Глиоциты
Количество: в 5-10 раз больше, чем нервных клеток.
Функции: опорная, стромальная, трофическая, защитная, всасывательная имвыделительная
Форма: призматическая.
Что выстилают? желудочки головного мозга и центральный канал спинного мозга.
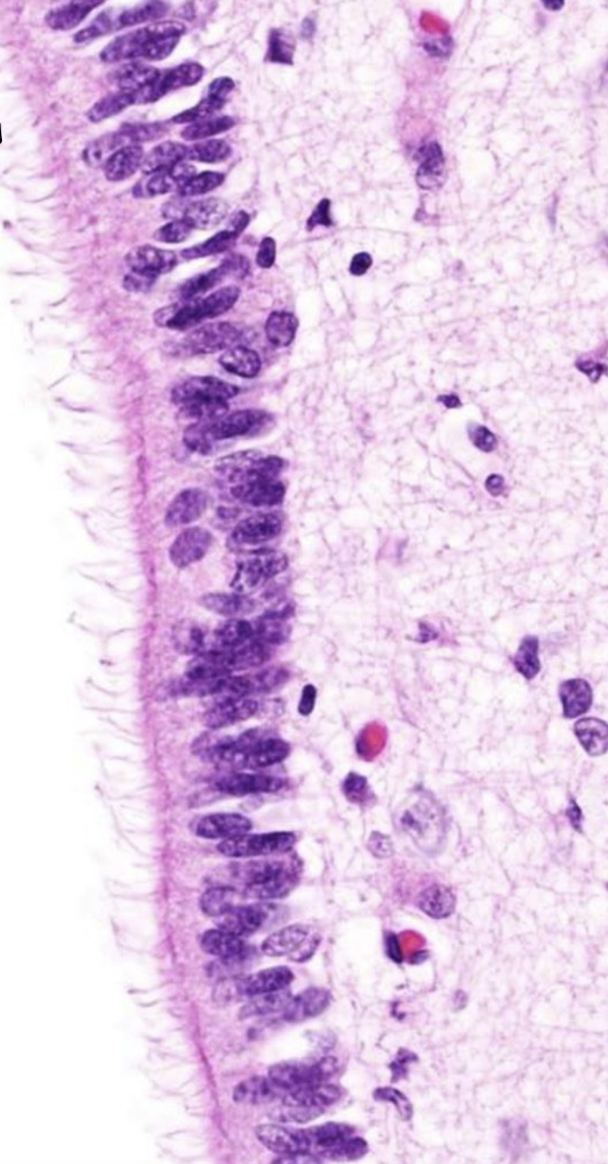
Они образуют эпендиму. Между соседними клетками плотные соединения отсутствуют. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости.
Танициты — клетки, базальная поверхность которых имеет длинный отросток, пронизывающий все вещество мозга и на его поверхности образующий отграничительную глиальную мембрану. Многочисленны в дне III желудочка, передают информацию о составе цереброспинальной жидкости на первичную капиллярную сеть воротной системы гипофиза.
Эпендимный эпителий сосудистых сплетений желудочков продуцирует цереброспинальную жидкость (ликвор)
Эпендимоциты
Волокнистые (фиброзные)
Локализуются в белом
веществе ЦНС
Имеют тонкие длинные
слабоветвящиеся отростки,
которые на концах
разветвляются и формируют
отграничительные мембраны.
Протоплазматические
Локализуются в сером
веществе ЦНС
Имеют многочисленные короткие
разветвления, широкие отростки,
часть которых окружает кровеносные
капилляры, участвуют в образовании
гематоэнцефалического барьера;
также отростки изолируют синапсы.
По отросткам переносятся из крови к
нейронам питательные вещества.
Функции: трофическая, защитная
(иммунобиологическая защита)
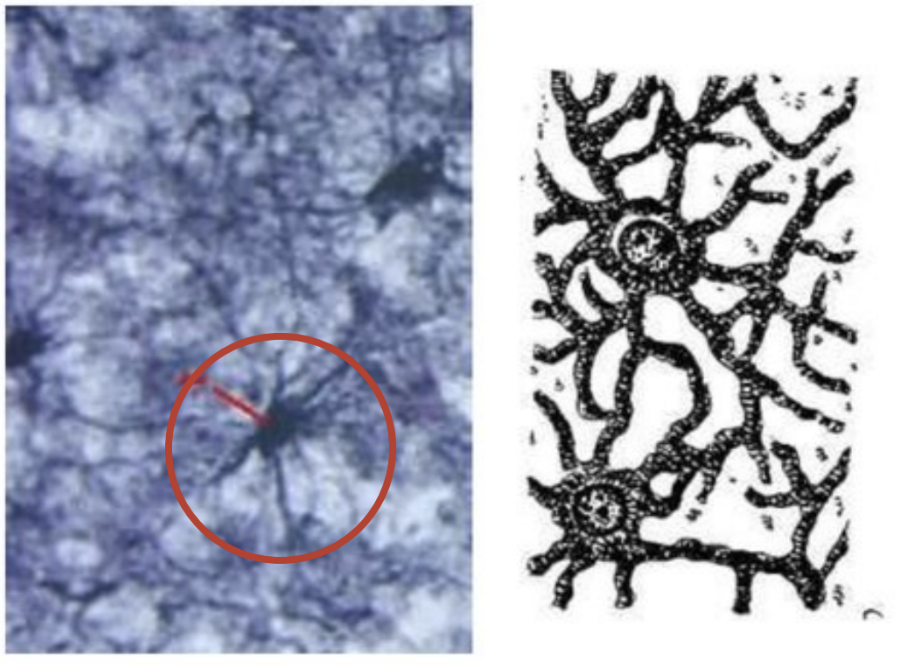
Астроциты
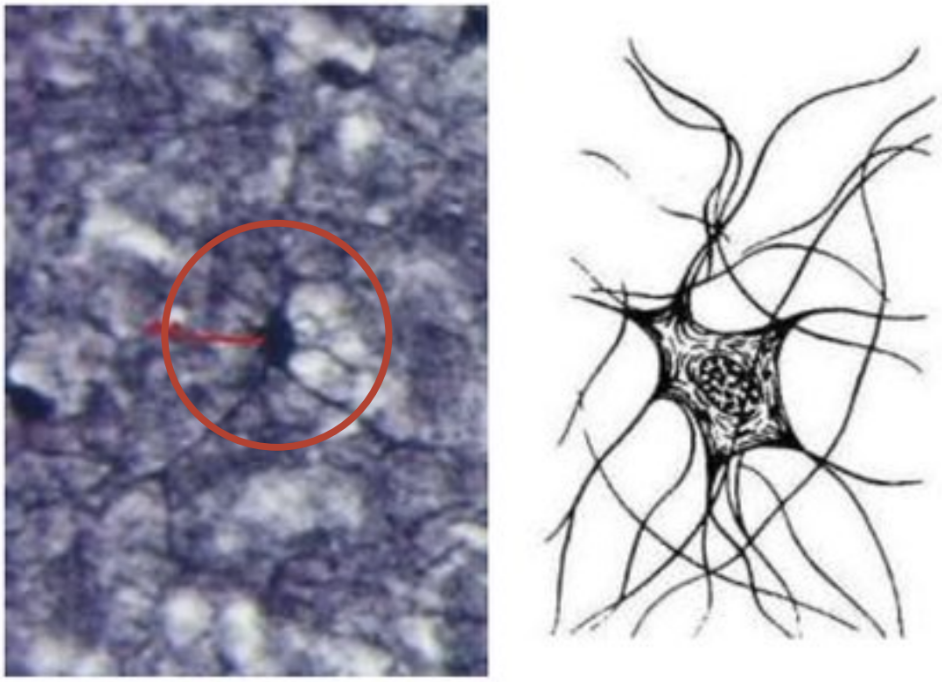
Олигодендроциты
Микроглия
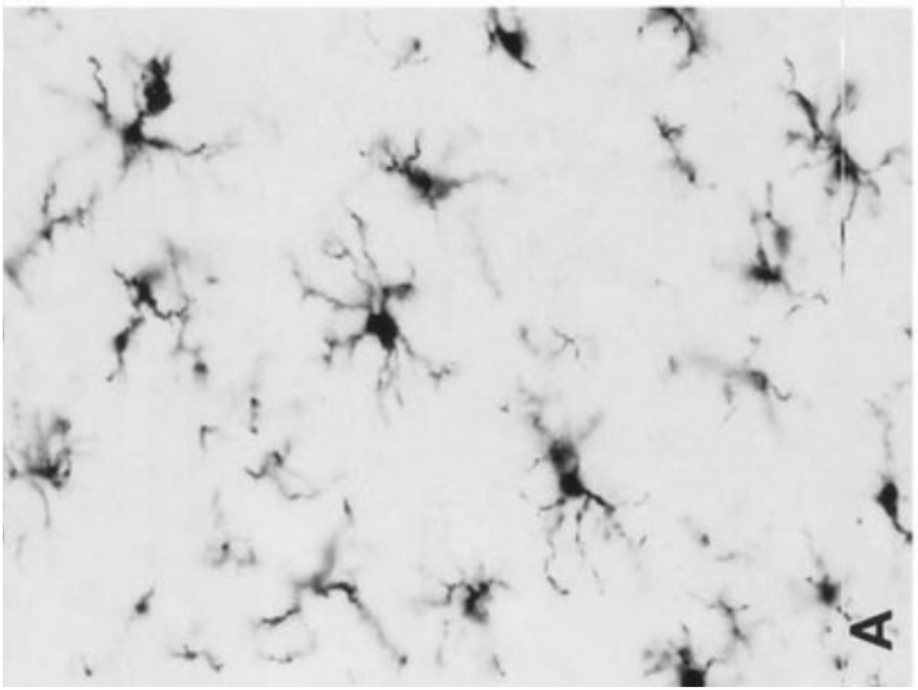
Представляет собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов. Клетки микроглии характеризуются небольшими размерами, тела их имеют продолговатую форму.
Имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах. В периферической нервной системе олигодендроциты представлены нейролеммоцитами, которые образуют оболочки вокруг отростков нейронов, и мантийными клетками, окружающими тела нейронов.
Функция: защита от инфекции и повреждения, удаление продуктов разрушения нервной ткани.
Ветвистая микроглия
Встречается как в сером, так и в
белом веществе центральной
нервной системы. В цитоплазме
клеток реактивной микроглии
присутствуют плотные тельца,
липидные включения, лизосомы.
Реактивная микроглия
Формируется вследствие
активации покоящейся
микроглии при травмах
центральной нервной системы.
Нейроны
Эффекторные
(эфферентные)
нейроны
Специализированные клетки нервной системы, ответственные за получение, обработку и передачу сигнала (на: другие нейроны, мышечные или секреторные клетки). Нейрон является морфологически и функционально самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами.
Тело клетки содержит крупное светлое ядро с I-2 ядрышками, в цитоплазме содержатся все органеллы, особенно канальцы гранулярной ЭПС. Рибосомы образуют скопления – глыбки базофильного вещества (нет в аксоне и аксональных холмиках) по всей цитоплазме, в них идет синтез всех необходимых веществ, которые от тела транспортируются по отросткам.
Дендриты представляют собой истинные выпячивания тела клетки. По дендритам распространяются импульсы к телу нейрона. Они содержат те же органеллы, что и тело клетки: глыбки хроматофильной субстанции, митохондрии, большое количество микротрубочек и нейрофиламентов.
Аксон – это отросток, по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а также гладкую эндоплазматическую сеть.
Рефлекторная дуга
В зависимости от функции различают три типа нейронов:
Ведущую роль в образовании и проведении нервного импульса выполняет плазмолемма нейронов. При действии раздражителя в зоне воздействия происходит волна деполяризации распространяется по плазмолемме.
Чувствительные
(афферентные)
нейроны
Образуют 1-ое звено рефлекторной дуги (спинномозговые узлы). Длинный дендрит идет на периферию и там заканчивается нервным окончанием, а короткий аксон в соматической рефлекторной дуге поступает в задние рога спинного мозга. Афферентный нейрон преобразует раздражение в нервный импульс.
Вставочные
нейроны
Располагаются в спинном и головном мозге; второе звено рефлекторной дуги, отвечает за передачу информации.
Передают информацию на рабочие клетки. Имеют короткие разветвленные дендриты и длинный аксон, который достигает скелетное мышечное волокно и через нервно-мышечный синапс передает нервный импульс.
![]()
![]()
![]()
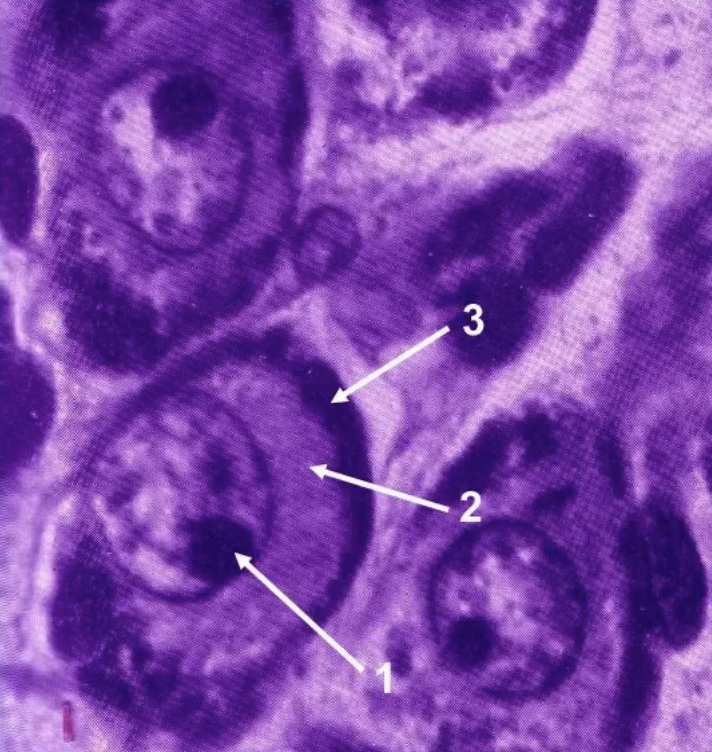
Функция: синтез и секретированные биологически активных веществ, в частности нейромедиаторов.
1 — ядро с эксцентричным ядрышком
2 — зона комплекса Гольджи и накопления нейросекрета (гранулы фиолетового цвета)
3 — хроматофильное в-во Ниссаля
Секреторные нейроны
В цитоплазме таких нейронов и в их аксонах находятся различной величины гранулы нейросекрета, содержащие белок, а в некоторых случаях липиды и полисахариды.
Гранулы нейросекрета выводятся непосредственно в кровь или в мозговую жидкость. Нейросекреты выполняют роль нейрорегуляторов, участвуя во взаимодействии нервной и гуморальной систем интеграции.
Отросток нервной клетки в нервном волокне называют осевым цилиндром, или аксоном, так как чаще всего (за исключением чувствительных нервов) в составе нервных волокон находятся именно аксоны. В ЦНС оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в ПНС — нейролеммоцитами.
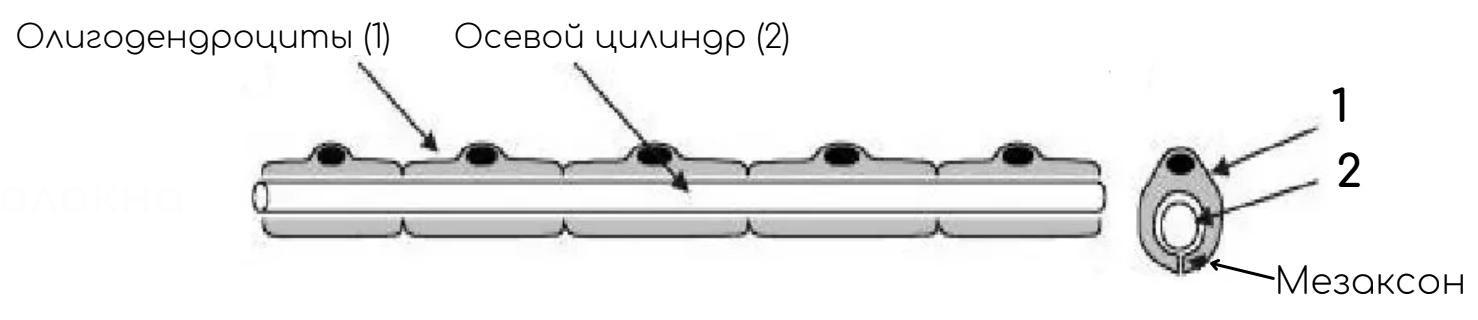
Нервные волокна
Безмиелиновые нервные волокна
Место нахождения: в составе автономной, или вегетативной, нервной системы.
Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров (волокна кабельного типа), принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее.
По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану – мезаксон, на которой как бы подвешен осевой цилиндр. Скорость проведения импульса 1-5 м/с.
Где встречается? в центральной и в периферической нервной системе
Они значительно толще безмиелиновых нервных волокон. Диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее.
Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. Через определенные интервалы (1-2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье.
В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Образуется двойная складка плазмолеммы нейролеммоцита – мезаксон, который удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону – миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой.
Оболочка аксона (аксолемма) обладает в области перехвата значительной электронной плотностью. Отрезок волокна между смежными перехватами называется межузловым сегментом. Скорость передачи импульса миелиновыми волокнами – 5-120 м/с.
Для миелиновых волокон характерно сальтаторное проведение возбуждения, т.е. прыжками. Между перехватами идет электрический ток, скорость которого выше, чем прохождение волны деполяризации по аксолемме.
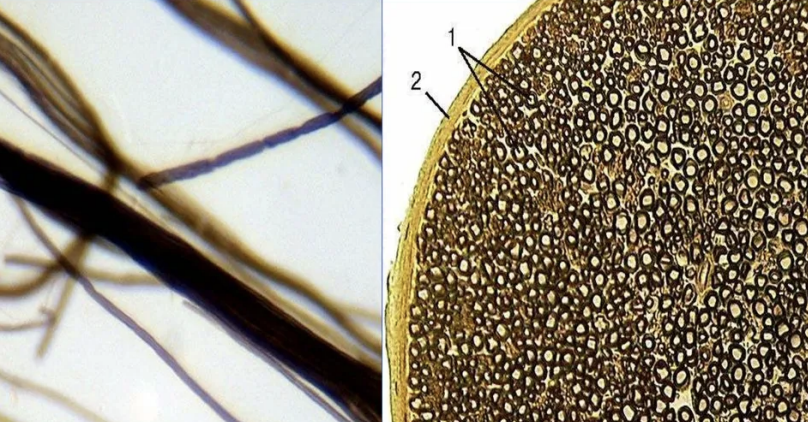
![]()
![]()
Миелиновые нервные волокна
Нервный импульс доходит до пресинаптической части и активирует синаптические пузырьки. Синаптический пузырек подходит к пресинаптической мембране, сливается с ней и нейромедиатор из синаптического пузырька попадает в синаптическую щель и действует на рецептор постсинаптической мембраны, что вызывает её деполяризацию, которая передается по центральному отростку следующего нейрона.
Аксо-аксональные
Межнейрональные контакты
![]()
![]()
Аксо-соматические
Аксо-дендритические
![]()
Синапсы – это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры.
Синапсы определяют направление проведения импульса. Нервные клетки соединены между собой посредством синапсов.
Эффекторные синапсы – синапсы, которые заканчиваются на рабочих клетках. Нервно-мышечные синапсы образуются на скелетном мышечном волокне; содержат пресинаптическую часть, которая образована конечным терминальным отделом аксона двигательного нейрона и внедряется в скелетное мышечное волокно. А прилежащий участок скелетного мышечного волокна образует постсинаптическую часть. В этой части отсутствуют миофибриллы, но в большом количестве располагаются ядра и митохондрии, а сарколемма формирует постсинаптическую мембрану
Постсинаптическая часть содержит постсинаптическую мембрану, которая содержит высокоспецифичные белковые рецепторы, реагирующие
только на конкретные медиаторы. Между пресинаптической и
постсинаптической частями находится синаптическая щель.
Двигательные нервные окончания – это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы.
При их участии нервный импульс передается на ткани рабочих органов.
Возбуждающие
Содержат возбуждающие
нейромедиаторы
(ацетилхолин, адреналин,
норадреналин,
глютаминовая кислота)
![]()
![]()
![]()
Адренергические
Передача импульса
совершается с помощью
медиатора адреналина
![]()
Холинергические
Передача импульса
совершается с помощью
медиатора ацетилхолина
Тормозные
Содержат тормозные
нейромедиаторы
(глицин, ГАМК — гамма
аминомасляная кислота)
СИНАПСЫ
и
Рецепторы
![]()
Экстерорецепторы
- слуховые
- зрительные
- обонятельные
- вкусовые
- осязательные
Интерорецепторы
- висцеро-рецепторы (сигнализирующие о состоянии внутренних органов)
- проприорецепторы (рецепторы опорно-двигательного аппарата)
![]()
Рецепторы рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов.
Соответственно выделяют две большие группы рецепторов:
В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания делят на механорецепторы, барорецепторы, хеморецепторы, терморецепторы и некоторые другие.
![]()
![]()
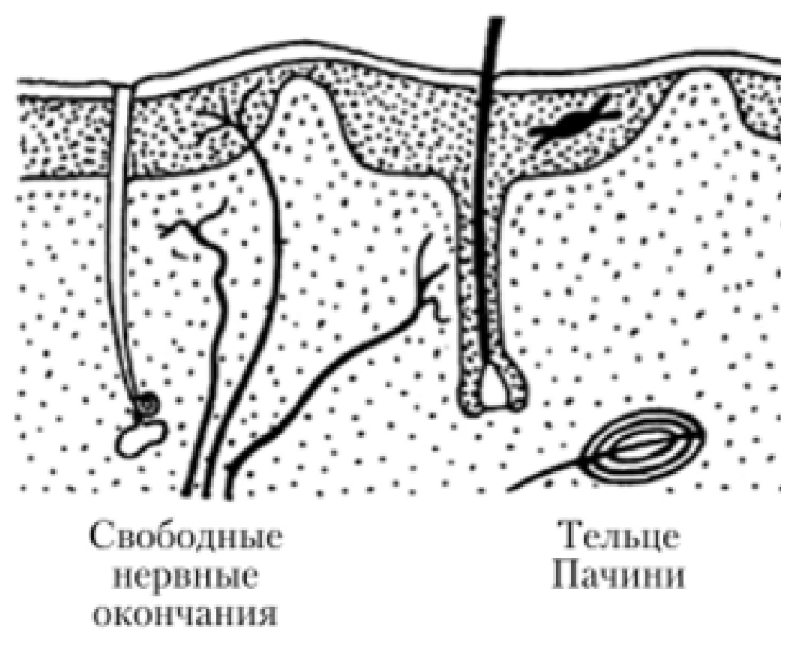
Инкапсулированные
- Покрытые соединительной тканной капсулой
- Тельца Фатера — Пачини
Неинкапсулированные
- Не покрыты соединительной тканной капсулой
- Тельца Мейснера
По особенностям строения чувствительные окончания подразделяют на:
Свободные нервные окончания
Состоящие только из конечных ветвлений осевого цилиндра
Воспринимают холод, тепло и боль. Такие окончания характерны для эпителия. В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви.
Несвободные нервные окончания
Содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии
Регенерация
Нервная клетка сохраняет способность к регенерации при условии сохранения тела нейрона, а отростки и нервные волокна регенерируют примерно со скоростью 1-2 мм в сутки.
Регенерация зависит от места травмы. Как в центральной, так и в
периферической нервной системе погибшие нейроны не восстанавливаются.
Полноценной регенерации нервных волокон в центральной нервной системе
обычно не происходит, но нервные волокна в составе периферических нервов
обычно хорошо регенерируют.
Поврежденные нервные волокна головного и спинного мозга не регенерируют.
Однако при малых травмах центральной нервной системы возможно
частичное восстановление ее функций, обусловленное пластичностью
нервной ткани.
Лекция № 8
НЕРВНАЯ ТКАНЬ. НЕРВНЫЕ ВОЛОКНА. НЕРВНЫЕ
ОКОНЧАНИЯ
План:
1.Общая морфофункциональная
характеристика нервной ткани.Гистогенез.
2.Морфологическая и функциональная
классификация нейроцитов.
3.Микро -и субмикроскопическое строение
нейроцита.Общие и
специальные органеллы.
4.Классификация, гистологическая стуктура и функции нейроглии.
5.Нервные волокна.Классификация.
6.Микроскопическое и
субмикроскопическое строение миелинового нервного волокна.
7.Гистофизиология безмиелинового
нервного волокна.
8.Морфофункциональная характеристика
нервных окончаний.
9.Рецепторы, их классификация и
структура.
10.Эффекторы, классификация и
структура.
11.Межнейронные синапсы, их
классификация и морфология.
12.Строение простой рефлекторной дуги.Роль синапсов.
Нервная ткань (нейроциты, нейроглия)
Нервная ткань (textus nervosus)
принадлежит к специальным тканям, ее элементы способны воспринимать
раздражение, трансформировать это раздражение в нервный импульс, быстро его
передавать, хранить информацию, продуцировать биологически активные вещества,
благодаря чему нервная ткань обеспечивает согласованную деятельность органов и
систем организма и его адаптацию к условиям внешней среды. Нервная ткань
построена из нервных клеток (нейронов, нейроцитов) и
вспомогательных элементов, которые объединяются под названием нейроглии.
Нейроны.
Нейроны являются морфологическими и
функциональными единицами нервной ткани.
Состоят из тела (перикариона) и отростков.
Наличие последних является самым характерным признаком нервных клеток. Именно
отростки обеспечивают проведение нервного импульса часто на достаточно длинное
расстояние, поэтому длина их колеблется от нескольких микрометров до 1.,,1,5м.
Нейроны не способны к митотическому делению, имеют длинный жизненный цикл. Срок
их жизни совпадает со сроком жизни индивида. Размеры перикариона нейронов очень разнообразны — от 5… 8 мкм
(клетки-зерна мозжечка) до 120 мкм (гигантопирамидные)
нейроны коры головного мозга). Среди отростков нервных клеток различают
аксоны и дендриты
Аксон (нейрит) — это длинный отросток, длина которого может достигать 1,5 м. Название его походит от греческого аксис — ось. Он всегда в клетке лишь один.
Диаметр аксона по всей длине является неизменным, он не ветвится, но может
давать коллатерали, которые имеют другое направление. Заканчивается аксон
терминальным разветвлением. Это отросток, который проводит нервный импульс в
направлении от тела клетки. Дендриты — это чаще всего короткие деревовидные
отростки, которые ветвятся (название их происходит от греческого дендрон — дерево); основы дендритов имеют коническое
расширение. Нервный импульс эти отростки передают по направлению к телу
клетки.
Считается общепринятым, что синтез
белка в нейроне происходит только в перикарионе и
дендритах. Однако в последнее время появились достаточно убедительные
доказательства синтеза белка в аксоне. Многочисленные белки аксоплазмы
синтезируются в перикарионе и перемещаются в составе
медленного аксонного транспорта.
Они содержат рибосомы, компоненты гранулярной
и гладкой эндоплазматической сети, элементы комплекса Гольджи.
Дендриты интегрально образуют до 95% всей рецепторной поверхности (рецептивного
поля) нейрона.Дендриты
являются наиболее чувствительными индикаторами функционального состояния
нервных связей и любых изменений в нервной системе. Дендриты, в отличие от
аксонов, способны генерировать не только потенциалы действия, возникающие по
закону всё или ничего, но и градуальные генераторные
потенциалы.
ЦИТОСКЕЛЕТ
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламенты) и микрофиламентов.
Микротрубочки наиболее крупные
элементы цитоскелета, их диаметр 24 нм. С ними
связывают внутриклеточный, в том числе аксонный
транспорт. От перикариона по отросткам перемещаются
различные вещества (белки, нейромедиаторы и т.д.),
органеллы (митохондрии, элементы цитоскелета,
везикулы и т.д.). Микротрубочки в перикарионе и
дендритах (в отличие от аксона) не имеют направленной ориентации. Большинство
микротрубочек аксона одним концом направлено к терминали,
а другим- к перикариону.
Характер ориентации микротрубочек имеет важное значение
для распределения по отросткам различных органелл. К 1 концу перемещаются
митохондрии и секреторные пузырьки, а ко 11 концу рибосомы,
мультивезикулярные тельца, элементы комплекса Гольджи.
Нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим
связям, характеру ветвления дендритов, электрофизиологическим характеристикам,
химии нейромедиаторов, позиции в функциональных сетях
и множеству других характеристик.
Нервные клетки содержат в центре перикариона одно большое круглое светлое ядро с малым количеством гетерохроматина, одним или несколькими ядрышками. В
нейронах некоторых ганглиев вегетативной нервной системы насчитывается до 15
ядер.
Цитоплазма
нервной клетки (нейроплазма) содержит три типа организованных структур: общие
органеллы, включения и специальные органеллы. Включениями нейроплазмы
могут быть углеводы (гликоген), пигментные вещества (липофусцин, меланин) и
разнообразные секреты (в нейросекреторных клетках). Специальными органеллами
нейронов является хроматофильная субстанция и
нейрофибриллы.
Под световым микроскопом хроматофильная
субстанция имеет вид разных по размерам и форме комочков и зерен, которые
окрашиваются базофильно, локализованные в перикарионе и дендритах нейронов и никогда не оказываются в
аксонах и начальных сегментах последних. Хроматофильное
вещество впервые описал Ф. Ниссль в 1889 г., в связи с чем оно носило его имя (субстанция Ниссля). Й.Леношек (в 1845 г.) дал ей название тигроид. Хроматофильную
субстанцию также называют базофильным веществом.
Под электронным микроскопом эта
структура оказывается гранулярной эндоплазматической сетью с параллельным
расположением ее сплющенных цистерн (так называемая эргастоплазма), где
интенсивно синтезируется белок, что характерно для нервной клетки. Хроматофильная субстанция является показателем
функционального состояния нейрона. Она может исчезать при истощении нервной
клетки (так называемый хроматолиз, или тигролиз), а
затем возобновляться.
В аксонах, которые не содержат органелл белкового синтеза, цитоплазма
постоянно перемещается от перикариона к терминалам со
скоростью 1…3 мм на сутки. Это так называемый медленный аксонный
транспорт, за счет которого происходит доставка белков (например, ферментов,
необходимых для синтеза медиаторов в синаптических
окончаниях). Кроме того, существует быстрый аксонный
транспорт (5…10 мм/час), который переносит, главным образом, вещества,
необходимые для синаптической функции, дендритный
транспорт (скорость 3 мм/час) и ретроградный поток, с помощью которого ряд
компонентов цитоплазмы возвращается из окончаний в тело клетки. Транспорт
веществ по отросткам нейронов обеспечивают такие органеллы: эндоплазматическая
сеть, микротрубочки, актомиозиновая система цитоскелета.
Нейрофибриллы можно обнаружить в цитоплазме при импрегнации серебром.
Они имеют вид тонких нитей диаметром 0,3… 0,5 мкм, образуют плотную сеть в перикарионе и имеют параллельную ориентацию в составе
дендритов и нейритов, включая их самые тонкие конечные разветвления. Методом
электронной микроскопии обнаружено, что нейрофибриллам отвечают пучки нейрофиламентов (микрофиламентов)
диаметром 6…10 нм и нейротубулы. (микротрубочки)
диаметром 20… З0 нм. Микрофиламенты
и микротрубочки принадлежат к системе цитоскелета
нейронов. Последний построен главным образом из белка спектрина.
Морфологическая
классификация нейронов
Эта
классификация нейронов базируется на количестве отростков. По этому признаку нервные клетки разделяют на такие разновидности:
1) униполярные (имеют единственный отросток, который является аксоном); 2)
биполярные (имеют два отростка — аксон и дендрит); 3) псевдоуниполярные
(имеют один отросток, который на определенном расстоянии от тела клетки
разделяется на аксон и дендрит, так что фактически клетка имеет два отростка,
как и биполярная); 4) мультиполярные (имеют
много отростков, один из которых является аксоном, а все другие дендритами. В
организме человека подавляющее большинство нейронов являюся
мультиполярными; биполярные клетки: лишь в сетчатке
глаза и в спиральном ганглии завитки, а псевдоуниполярные
— в спинномозговых узлах. Униполярные клетки в теле человека не обнаружены.
Такое строение (с одним отростком — аксоном) имеют лишь нейробласты.
Функциональная
классификация нейронов
Функциональная классификация нейронов
базируется на положении нервной клетки в составе рефлекторной дуги. Согласно с
этой классификацией различают такие виды нейронов: 1) афферентные (рецепторные,
чувствительные) воспринимают раздражение и трансформируют его в нервный
импульс; 2) ассоциативные (вставные) передают нервный импульс между нейронами;
3) эфферентные (моторные, двигательные) обеспечивают передачу нервного импульса
на рабочую структуру.
Рефлекторная
дуга — это цепочка нервных
клеток, которые передают нервный импульс от чувствительного нервного окончания
(рецептор) к двигательному нервному окончанию (эффектор), который расположен в
рабочем органе.
Самая простая рефлекторная дуга состоит из двух нейронов: афферентного, дендрит которого заканчивается рецептором, а аксон передает
импульс на дендрит эфферентного нейрона; эфферентного, который своим аксоном
передает импульс к эффектору в рабочем органе.
Сложные рефлекторные дуги имеют между афферентными и эфферентными
нейронами несколько ассоциативных нервных клеток. Нервное возбуждение по
рефлекторной дуге передается лишь в одном направлении, которое имеет название
физиологичной (или динамичной) поляризации нейронов.
Изолированный нейрон, как показал А. И.
Бабухин, способный проводить импульс в любом направлении. Однонаправленность
передачи импульса в пределах рефлекторной дуги предопределена
структурой межнейронного контакта, который имеет название синапса.
Нейроглия
Термин нейроглия ввёл немецкий патолог
Рудольф Вирхов в 1846 г.,
означает в буквальном переводе нервный клей, а в действительности это среда,
которая окружает нейроны для описания
связующих элементов между нейронами. Эти клетки составляют почти половину
объёма мозга. Нейроны существуют в тесной генетической, структурной и
функциональной связи с нейроглией. Построена нейроглия из клеток. Ее функции:
опорная, разграничительная, трофическая, секреторная, защитная. Все клетки
нейроглии разделяют на два генетических вида: глиоцити
(макроглия) и глиальные
макрофаги (микроглия), В свою очередь, среди глиоцитов различают эпендимоциты,
астроциты и олигодендроциты
Макроглия походит, как и нейроны, из нервной трубки,
а микроглия — из моноцитов и принадлежит к макрофагической системе. В последнее время, появились
данные, что микроглия не имеет моноцитарного
генезиса.
Астроциты звёздчатые клетки, их отростки отходят от тела клетки в
разных направлениях, оплетают нейроны, сосуды, клетки (эпендимы) желудочков мозга,
образуя расширения в виде концевой ножки.Астроциты
имеют адренорецепторы и рецепторы многих нейромедиаторов. Астроциты образуют
опорный аппарат центральной нервной системы. Различают протоплазматичиские
и волокнистые (фибриллярные) астроциты; существуют
также и переходные формы астроцитов
(волокнисто-протоплазматические). Протоплазматические астроциты локализируются преимущественно в сером веществе
мозга. Размеры их 15…25 мкм. Отростки короткие и толстые, сильно разветвлены.
На импрегнированных металлами препаратах эти клетки напоминают заросли
кустарника. Волокнистые астроциты
преимущественно расположены в белом веществе мозга. Отростки их длинные,
прямые, слабо или совсем не разветвленные, на поперечном разрезе круглой или
овальной формы.
Отростки астроцитов
оканчиваются на сосудах, нейронах, базальной мембране, которая отделяет
мозговую ткань от мягкой мозговой оболочки. В цитоплазме астроцитов
содержатся фибриллы, которые состоят из филаментов.
Функции астроглии многочисленны.
* В гистогенезе проводящие
пути для миграции недифференцированных нейронов в коре мозжечка и для врастания
аксонов в зрительный нерв.
* Транспорт метаболитов из
капилляров мозга в нервную ткань. Астроцитарные ножки
почти полностью покрывают капилляры мозга.
* Регуляция химического состава
межклеточной жидкости. Астроциты участвуют в
метаболизме глутаминовой и аминомасляной
кислот, соответственно возбуждающего и тормозного нейромедиаторов
ЦНС. После высвобождения этих нейромедиаторов
в синаптическую щель часть молекул поступает в астроциты, где превращается в глутамин.
* Астроциты
изолируют рецептивные поверхности нейронов.
* Участие в патологических
процессах пролиферация и замещение погибших нейронов.
* Фагоцитоз и экспрессия Аг MHC II.
* Астроциты
выделяют ряд веществ, способствующих росту аксонов.
Эпендимоциты образуют плотный, эпителиообразный
пласт клеток, которые выстелают спинномозговой канал
и все желудочки мозга. Эпендимоциты возникают первыми
в процессе гистогенеза нервной ткани из глиобластов
нервной трубки. На этой стадии развития они выполняют разграничительную и
опорную функции. На поверхности клеток, обращённой в полость канала нервной
трубки, образуются реснички, которых может быть до 40 на одну клетку. Возможно,
реснички способствуют движению жидкости в полостях мозга. От базального конца эпендимоцита отходят длинные отростки, которые
разветвляются и пересекают всю нервную трубку, образовывая ее опорный аппарат.
На внешней поверхности трубки эти отростки образуют поверхностную глиальную пограничную мембрану, которая отграничивает
нервную трубку от других тканей.
После рождения эпендимоциты
выполняют лишь функцию выстелания полостей мозга.
Некоторые эпендимоциты выполняют секреторную функцию.
Например, эпендимоциты субкомиссурального
органа продуцируют секрет, который, возможно, принимает участие в регуляции
водного обмена. Особое строение имеют эпендимоциты,
что выстелают сосудистые сплетения желудочков мозга.
Цитоплазма базального полюса этих клеток образует многочисленные глубокие
складки, содержит большие митохондрии и разные включения. Существует мнение,
что эти эпендимоциты активно участвуют в образовании
цереброспинальной жидкости и регуляции ее состава.
Олигодендроциты-это самая многочисленная группа глиоцитов.
Они отличаются небольшими размерами, наличием коротких, очень тонких отростков.
Тела их имеют многоугольную или овальную форму. Олигодендроциты
окружают тела нейронов и их отростки по всей длине, локализуются как в
центральной, так и периферической нервной системе.
Цитоплазма олигодендроцитов
не содержит нейрофиламентов. Функции этих клеток
очень разнообразны: трофическая, изолирующая, участие в водно-солевом обмене,
процессах дегенерации и регенерации нервных волокон. Олигодендроцити,
которые образуют оболочки вокруг отростков нервных клеток, имеют название нейролеммоцитов (шванновских
клеток).
Олигодендро(глио)циты, как правило, более мелкие клетки, чем астроциты, но в этих миелинобразующих
клетках ЦНС высока плотность органелл. В сером
веществе мозга олигодендроциты находятся в
непосредственном контакте с перикарионами и
отростками нейронов. В белом веществе олигодендроциты
расположены рядами между нервными волокнами. Именно
миелин придает белому веществу характерный цвет, отличающий его от серого
вещества. Шванновские клетки входят в состав миелиновых
и безмиелиновых периферических нервных волокон,
синтезируют белки P0, P1, P2, образуют миелин и рассматриваются как аналоги олигодендроцитов. Каждая шванновская
клетка миелинизирует один аксон. Шванновские
клетки образуют щелевые контакты.
Микроглия — это совокупность маленьких клеток с двумя-тремя
отростками, которые имеют на своей поверхности короткие вторичные и третичные
разветвления. Ядра клеток вытянутой или треугольной формы, богатые на гетерохроматин. При раздражениях нервной ткани (воспаление,
рана) клетки микроглии изменяются — увеличивается
объем ядра и цитоплазмы, клетки становятся круглыми, подвижными, втягивают свои
отростки. Подобно другим макрофагам микроглиоцити
наполняются фагоцитованным материалом. В таком виде
их называют зернистыми шарами. В последнее время показана способность микроглии принимать участие в
синтезе белков-иммуноглобулинов (антител).
Нервные
волокна — это отростки нервных
клеток, покрытые оболочками. В зависимости от строения оболочки они разделяются
на две основных группы — миелиновые и безмиелиновые.
И те, и другие построены из осевого цилиндра, который является отростком
нервной клетки и оболочки, образованной клетками олигодендроглии
( н е й р
о л е м м о ц и т а м и, ш в а н н о в с к и м и клетками).
Миелиновые нервные волокна
имеют достаточно сложное строение. Они встречаются как в центральной, так и в
периферической нервной системе, то есть в составе головного и спинного мозга, а
также в составе периферических нервов. Это толстые волокна, диаметр их
поперечного сечения колеблется от 1 до 20 мкм. Они построены из осевого
цилиндра, миелиновой оболочки, нейролеммы и базальной
мембраны. Осевой цилиндр — это отросток нервной клетки, которым чаще бывает
аксон, но может быть и дендрит. Он состоит из нейроплазмы,
которая содержит продольно ориентированные нейрофиламенты и нейротубулы, а
также митохондрии. Осевой цилиндр покрыт аксолеммой
(продолжением клеточной мембраны) которая обеспечивает проведение нервного
импульса.
Миелиновая оболочка — это трубка толщиной от 0,3 до 15…20 мкм, которая
одевает осевой цилиндр. Она отсутствует в месте выходження отростка от перикариона,
в участке терминальных разветвлений аксона и в участках, которые имеют название
узловых перехватов. Участок волокна между двумя соседними перехватами
называется межузловым сегментом. Длина последнего —
от нескольких микрометров до нескольких миллиметров.
Миелиновое нервное волокно состоит из осевого цилиндра, вокруг которого шванновские клетки образуют миелин за счёт удлинения и
концентрического наслаивания мембран мезаксона.
Каждая шванновская клетка миелинизирует
небольшой сегмент только одного аксона. В миелине периферических нервов присутствуют
небольшие просветления насечки миелина. Снаружи от миелина располагаются
тонкий слой цитоплазмы шванновской клетки и её ядро.
Узловые перехваты. На границе между
соседними шванновскими клетками участок
плазматической мембраны аксона (аксолемма) не прикрыт
миелином. Здесь шванновские клетки образуют
многочисленные переплетающиеся отростки. Аксолемма
перехватов содержит множество потенциалозависимых Na+ -каналов, необходимых для поддержания импульсной
активности. Узловой перехват имеет размеры 0,25.. 1 мкм.
Скачкообразное проведение нервных импульсов
в миелиновых волокнах, когда сигнал перескакивает от одного перехвата к
другому, как раз и обеспечивают Na+-каналы
перехватов. По этой же причине в миелиновых волокнах (в отличие от не имеющих
перехватов безмиелиновых волокон) скорость проведения
выше.
Насечки миелина участки
расслоения миелина, образовавшиеся при миелинизации;
в них присутствует цитоплазма шванновских клеток.
Функция насечек неясна.
Миелиновая оболочка содержит липиды и
потому окрашивается в черный цвет при обработке осмиевой кислотой. С помощью
электронного микроскопа было обнаружено, что миелиновая оболочка имеет
пластинчатое строение. Окончательно понять строение миелиновой оболочки помогли
исследования процесса развития миелиновых нервных волокон.
В процессе развития миелинового волокна
осевой цилиндр погружается в нейролеммоцит, вгибая
его оболочку и образовывая глубокую складку. Эта двойная складка (дупликатура) плазмолеммы нейролеммоцита
получила название мезаксона. В процессе последующего
развития шванновская клетка медленно вращается вокруг
осевого цилиндра, в результате чего мезаксон
многократно окутывает его. Цитоплазма леммоцита и его
ядро остаются на периферии, образовывая нейролемму.
Таким образом, миелиновая оболочка образуется из плотно, концентрически наслоенных вокруг осевого цилиндра.
На продольном разрезе миелинового
нервного волокна вблизи узловой перетяжки есть участок, в котором завитки мезаксона последовательно контактируют с осевым цилиндром.
Места прикрепления самых глубоких завитков более удалены от перехватове
следующие — постепенно приближаются к нему. Это объясняется тем
что мезаксон наслаивается в процессе роста и осевого
цилиндра и нейролеммоцитов, образуют пальцеобразные
выросты диаметром 50 нм. Длина выростов разная. Вместе они имеют характерный
вид пышного воротничка.
Н е й р о л е м м а — тонкая, светлая при
обработке осмиевой кислотой оболочка нервного волокна, расположенная внешне от
миелинового слоя. Нейролемма образована
цитоплазматичними частями нейролеммоцитов
и ядрами. Базальная мембрана, укрывая внешне нервное волокно, соединяется с коллагеновыми волокнами эндоневрию
(соединительной тканью, которая окружает нервные волокна).
Вышеописанное строение имеют
периферические миелиновые нервные волокна. Миелиновые волокна
центральной нервной системы имеют ряд особенностей строения: их оболочку,
вместо нейролеммоцитов, образуют типичные олигодендроцити (в последних меньше цитоплазмы, они
мельче); отсутствуют насечки миелина и базальная мембрана; узловые перехваты
имеют большие размеры, а межузловые сегменты короче.
Безмиелиновые
нервные волокна являются типичными
для автономного отдела нервной системы. Диаметр волокон 1 — 4 мкм, то есть они
тоньше от миелиновых волокон. Строение их значительно проще. Безмиелиновые волокна состоят из осевого цилиндра, нейролеммы и базальной мембраны. Нейролемма
образована тяжем нейролеммоцитов, которые плотно прилегают друг к другу.
Прогибая оболочку нейролеммоцитов, осевой цилиндр
глубоко погружается в этот тяж, а глиальная клетка,
как муфта, одевает отросток. Если тяж леммоцитов
охватывает не один осевой цилиндр, а несколько (10-20), то такие безмиелиновые волокна называют полиаксонными,
или волокнами кабельного типа. Внешне безмиелиновое
нервное волокно, как и миелиновое, покрыто базальной мембраной.
Скорость передачи нервного импульса
миелиновыми нервными волокнами значительно высшая (5… 120м/с), чем безмиелиновыми (1…2 м/с). Это
объясняется тем, что в безмиелиновом волокне волна
деполяризации двигается по всей плазмолемме не прерываясь, а в миелиновом — она
идет сальтаторно, то есть прыжками, возникая лишь в
участках перехватов.
Нервные окончания (terminationes nervorum) разделяют на рецепторы, эффекторы и межнейронные
синапсы.
Рецепторы- чувствительные
окончания дендритов нервных клеток, приспособленные к восприятию раздражений,
которые поступают к организму. Различают э к с т е р о р е ц
е п т о р ы, которые воспринимают раздражение из внешней среды, и н т е р о р
е ц е п т о р ы, раздражения к которым
поступают от собственных тканей организма. Разновидностью интерорецепторов
является п р о п р
и о р е ц е п т о р ы
— чувствительные нервные окончания в мышцах и сухожилиях, которые принимают
участие в регуляции движений и положения тела в пространстве. В зависимости от
природы раздражений, которые вызывают возбуждения чувствительных нервных
окончаний, последние разделяют на терморецепторы (воспринимают изменения
температуры), механорецепторы (воспринимают действие
механических раздражителей), барорецепторы
(воспринимают изменения давления), хеморецепторы (воспринимают действие
химических раздражителей), ноцирецепторы
(воспринимают болевые раздражения) и др.
В зависимости от строения существуют
свободные и несвободные нервные окончания.
Свободные нервные окончания состоят лишь из разветвлений осевого цилиндра.
Несвободные рецепторы, кроме осевого цилиндра, включают также клетки
нейроглии. Если несвободные нервные окончания окружает соединительнотканная
капсула, они получают название инкапсулированных; те
несвободные рецепторы, которые не имеют соединительнотканной капсулы, называют неинкапсулированными. Рецепторные окончания в составе
эпителиальной, соединительной и мишечной тканей имеют
ряд особенностей строения, которые рассматриваются ниже.
Для эпителиев характерные свободные нервные окончания. Функция свободных
рецепторов, например, эпидермиса, связанная с восприятием болевых и
температурных раздражений. Свободные нервные окончания могут в виде корзины
оплетать волосяные фолликулы. Регистрируя смещение в пространстве отдельных
волосков, они играют роль механорецепторов.
В составе многослойных эпителиев локализованы одиночные чувствительные
эпителиальные клетки, так называемые осязательные эпителиоциты
Меркеля. Они специализированы на восприятии
механических раздражений. К базальной части клеток Меркеля
в виде дисков прилегают чувствительные нервные окончания. При этом образуются
так называемые осязательные мениски (диски М е р к е л я), которые выполняют функцию механорецепции.
Чувствительные нервные окончания в
составе соединительной ткани разделяются на несвободные неинкапсулированные
и инкапсуллированные рецепторы, а также
нервно-сухожильные веретена.
В инкапсулированных тельцах нервные окончания, как правило, окруженные нейролеммоцитами и вспомогательными элементами
соединительнотканного происхождения. Среди инкапсулированных нервных телец в
зависимости от строения различают пластинчатые тельца (Фатер-Паччини),
лукообразные тельца (Гольджи-Маццони), осязательные
тельца (Мейснера), конечные колбы (Краузе) .
Тельце Фатер-Паччини это образование овальной формы размерами около
0.52 мм.
Вокруг разветвлений нервного окончания, которое потеряло
миелиновую оболочку скопления видоизмененных нейролеммоцитов
образует внутреннюю колбу. Вокруг колбы концентрические наслоения коллагеновых волокон формируют так называемые пластинки,
между которыми залегают фибробласты. В своей совокупности пластинки и
фибробласты образуют внешнюю колбу, которая составляет основную массу
пластинчатого тельца. Телец Фатер-Паччини много в
соединительной ткани всех внутренних органов, а также в глубоких слоях дермы.
Они воспринимают изменения давления. Тельца Г о л ь д жи -М
а ц ц о н
и меньше телец Фатер-Паччини, имеют более тонкую
капсулу и относительно большую внутреннюю колбу. Встречаются в коже, серозных и
слизистых оболочках, выполняют функции барорецепции.
Осязательные тельца Мейснера находятся в
соединительной ткани кожи, в частности, в сосковом слое дермы. Это овальные образования
с размерами около 50-100 мкм. Внутри тельца Мейснера
перпендикулярно к поверхности кожи размещены нервные окончания. Осязательные
тельца являются рецепторами тактильной чувствительности.
Конечные колбы Краузе встречаются в
конъюнктиве глаза, соединительной ткани языка и наружных половых органов.
Характерная особенность колб Краузе — очень тонкая
соединительнотканная капсула. Считают, что колбы Краузе
являются механорецепторами.
Нервно-сухожильные веретена
(сухожильные органы Гольджи) образованы толстыми
(диаметром около 15 мкм) миелиновыми волокнами, которые, подходя к коллагеновым волокнам сухожилия, теряют миелиновую оболочку
и дают многочисленные разветвления, которые оплетают сухожильные щепотки.
Подобное строение имеют также тельца Руффини,
которые залегают в глубоких слоях дермы и подкожной жировой ткани. Тельца Руффини особенно многочисленны в участке подошвы стопы.
Нервно-сухожильные веретена, как и тельца Руффини,
считают механорецепторами, которые воспринимают
взаимное смещение коллагеновых волокон и изменение их
положения относительно окружающих тканей.
В мышечной ткани чувствительные нервные окончания образуют
нервно-мышечные веретена, которые воспринимают изменение длины мышечного
волокна и скорость этого изменения. Каждое веретено состоит из 10-12 тонких коротких
поперечно-исчерченых мышечных волокон, окруженных
соединительнотканной внутренней капсулой.
Среди внутриверетенных мышечных волокон есть
волокна с ядерной сумкой и с ядерной цепочкой. Волокна с ядерной сумкой в своей
центральной части содержат большое количество ядер. Волокна с ядерной цепочкой
вдвое тоньше и более короче от волокон с ядерной сумкой, их ядра размещены в
виде цепочки вдоль рецепторного участка.
Эффекторы образованы окончаниями аксонов нейроцитов
и бывают двух типов — двигательные и секреторные. Нервно-мышечные окончания в
скелетных мышцах образованы терминалями аксонов нейроцитов двигательных ядер передних рогов спинного мозга
или двигательных ядер головного мозга. Двигательные нервные окончания в
гладкой мышечной ткани имеют более простое строение: отдельные нервные
окончания образуют на поверхности гладких миоцитов
характерные расширения (варикозы), где в составе синаптических пузырьков накапливается ацетилхолин или
адреналин.
Межнейронные синапсы — особенная форма межклеточных связей, характерная для
нервной ткани. В составе синапса есть две части — пресинаптическая
и постсинаптическая, между которыми есть синаптическая
щель. Пресинаптическая часть (или полюс) образована
терминальной веточкой аксона той нервной клетки, которая передает импульс. Она
по большей части расширена в виде пуговицы, покрытая пресинаптической
мембраной. В этом полюсе содержатся митохондрии и синаптические
пузырьки, которые покрыты мембраной и имеют определенные химические вещества,
так называемые медиаторы. Последние способствуют
передаче нервного импульса на постсинаптическую часть
Синаптические пузырьки бывают разными по размерам и строению:
маленькие прозрачные (ЗО…60 нм), большие
электронно-плотные (80…150 нм), прозрачные, что содержат плотную гранулу
(50…90 нм). Медиаторами могут быть ацетилхолин (холинергичные
синапсы), норадреналин и адреналин (адренергические синапсы), а также другие
вещества —серотонин, вещество Р, глутаминовая кислота, энкефалин, нейротензин, ангиотензин II, вазоактивный интестинальний пептид, дофамин, глицин, гамааминомасляная
кислота. Три последних является тормозными медиаторами. Постсинаптическая
мембрана содержит особенный белок — рецептор медиатора, чем предопределено
действие последнего на постсинаптическую часть.
Синаптическая щель имеет размеры 20…30 нм, заполненная тканевой
жидкостью. Она может содержать электронноплотные
частицы (двойной слой электронноплотного материала,
разделенный просветом шириной 2 нм), или нитевидные структуры, которые
расположены на поверхности обоих синаптических
мембран вроде щетины в щетке. Возможно, такая структура служит для содержания
пре- и постеинаптических
мембран вместе.
При поступлении нервного импульса к окончанию пресинаптического
нейрона синаптичные пузырьки сливаются с пресинаптической мембраной, их содержание выливается в синаптическую щель и медиатор действует на
постсинаптический нейрон. Мембрана самих пузырьков используется повторно.
Функционально различают два вида
синапсов — возбудительные и тормозные.
Морфологические типы синапсов различают в зависимости от того, какие
части нейронов контактируют между собой: аксодендритные
(аксон первого нейрона передает импульс на дендрит второго); аксосоматические
(аксон первого нейрона передает импульс на тело второго); аксоаксонные
(терминале аксона первого нейрона заканчиваются на аксоне второго). Очевидно, аксоаксонные синапсы выполняют тормозную функцию. Кроме
того, между некоторыми нейронами найдены дендродендритные,
а также дендросоматичные синапсы. Таким образом,
любая часть нейрона может
образовывать синапс с любой частью
другого нейрона.
Большинство нейромедиаторов аминокислоты
и их производные. Одни нейроны модифицируют аминокислоты с образованием аминов
(норадреналин, серотонин, ацетилхолин), другие нейромедиаторов пептидной природы (эндорфины,
энкефалины). Лишь небольшое количество нейромедиаторов образовано не аминокислотами. Нейроны могут
синтезировать более одного нейромедиатора.
Нервная ткань развивается из нервной пластинки, которая является
утолщением эктодерми на спинной стороне зародыша.
Нервная пластинка последовательно превращается в нервный желобок и нервную
трубку, которая, отделяется от кожной эктодермы.
Источники информации:
а) основные:
1. Гистология, цитология и эмбриология
/ [Афанасьев Ю. И., Юрина Н. А., Котовский Е. Ф. и др.] ;
под ред. Ю. И. Афанасьева, Н. А. Юриной. [5-е изд., перераб.
и доп.]. М. :
Медицина. 2001. С. 268-301.
2. Данилов Р. К. Гистология.
Эмбриология. Цитология. : [учебник для студентов медицинских вузов] / Р. К.
Данилов М. :
ООО Медицинское информационное агенство, 2006. С. 178-192.
3. Волков К.С. Ультраструктура клеток и
тканей : учебное пособие-атлас / К.С. Волков, Н.В. Пасечка. Тернополь : Укрмедкнига, 2004. С. 82-93.
4. Материалы для подготовки студентов к
практическим занятиям по теме «Общие принципы организации тканей. Нервные ткани.» (Интранет).
б)
дополнительные:
1. Гистология :
[учебник] / под ред. Э. Г. Улумбекова, Ю. А. Чельшева. [3-е изд., перераб. и
доп.]. М. : ГЕОТАРМедиа, 2007. С.325-345 .
2. Кузнецов С. Л. Атлас по гистологии,
цитологии и эмбриологии / С. Л. Кузнецов, Н. Н. Мушкамбаров,
В. Л. Горячкина.
М.: Медицинское информационное агенство, 2002.
С.99-113 .
3. Практикум по гистологии, цитологии и
эмбриологии; под редакцией Н. А. Юриной, А. И. Радостиной.
М.: Изд-во УДН,
1989. С.116-129 .
компакт-диски:
http://intranet.tdmu.edu.ua/index.phpdir_name=cd&file_name=index.php#3
Другие гистологические сайты:
http://en.wikipedia.org/wiki/Histology