- Авторы
- Файлы
Чеснокова Н.П
1
Брилль Г.Е.
1
Моррисон В.В.
1
Понукалина Е.В.
1
Полутова Н.В.
1
1 ФГБОУ ВО «Саратовский Государственный медицинский университет им. В.И. Разумовского Минздрава России»
3.1 Нервная регуляция дыхания
Дыхательный центр представляет собой совокупность нейронов продолговатого мозга, обладающих ритмической активностью и определяющих ритм дыхательных движений. Бульбарный дыхательный центр выполняет две основные функции:
1) регуляцию двигательной активности дыхательных мышц (двигательная функция);
2) гомеостатическую, связанную с изменением характера дыхания при сдвигах газового состава и кислотно-основного равновесия в крови и тканях.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна (длительности вдоха, выдоха, величины дыхательного объема).
Нейроны дыхательного центра расположены в дорсомедиальной и вентролатеральной областях продолговатого мозга, образуя так называемую дорсальную и вентральную дыхательные группы. В указанных дыхательных группах расположены следующие виды нейронов:
1) ранние инспираторные, максимальная частота разряда которых приходится на начало инспирации;
2) поздние инспираторные нейроны, максимальная частота разряда – в конце инспирации;
3) полные инспираторные нейроны, характеризующиеся постоянной активностью в течение фазы вдоха;
4) постинспираторные нейроны, максимальный разряд которых обнаруживается в течение выдоха;
5)экспираторные нейроны, активность которых возрастает во второй части выдоха;
6) преинспираторные нейроны, максимальный пик активности проявляют перед началом вдоха.
В структурах бульбарного дыхательного центра различают так называемые респираторно-связанные нейроны, активность которых совпадает с ритмом дыхания, но они не иннервируют дыхательные мышцы, а обеспечивают иннервацию верхних дыхательных путей.
В соответствии с локализацией нейронов бульбарного дыхательного центра, различают дорсальную дыхательную группу (ДДГ) и вентральную дыхательную группу (ВДГ). Нейроны дорсальной дыхательной группы получают афферентные сигналы от легочных рецепторов растяжения по волокнам n. Vagus. Только часть инспираторных нейронов дорсальной группы дыхательного центра связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контрлатеральной стороной.
Вентральная дыхательная группа расположена латеральнее обоюдного ядра продолговатого мозга, подразделяется на ростральную и каудальную части. Причем, ростральная часть вентральной дыхательной группы представлена ранними, поздними, полными инспираторными и постинспираторными нейронами.
Дорсальная и вентральная группы нейронов в правой и левой половинах продолговотого мозга взаимосвязаны как в пределах одной половины, так и с нейронами противоположной стороны. В синхронизации деятельности контрлатеральных нейронов бульбарного дыхательного центра участвуют проприобульбарные нейроны и экспираторные нейроны комплекса Бетцингера.
Касаясь функциональных особенностей отдельных нейронов бульбарного дыхательного центра, следует отметить, что ранние инспираторные нейроны (активируются в момент вдоха) называют еще проприобульбарными, так как не направляют свои аксоны за пределы дыхательного центра продолговатого мозга и контактируют только с другими типами дыхательных нейронов. Часть полных и поздних инспираторных нейронов направляет свои аксоны к дыхательным мотонейронам спинного мозга. Все экспираторные нейроны каудальной части вентральной дыхательной группы направляют аксоны в спинной мозг. При этом 40% экспираторных нейронов иннервируют внутренние межреберные мышцы, а 60% — мышцы брюшной стенки.
Таким образом, нейроны бульбарного дыхательного центра в зависимости от их значимости в регуляции внешнего дыхания разделяют на три группы:
1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях;
2) нейроны, синаптически связанные с мотонейронами спинного мозга и регулирующие активность мышц вдоха и выдоха;
3) проприобульбарные нейроны, участвующие в генерации дыхательного ритма, аксоны которых обеспечивают связь только с нейронами продолговатого мозга.
Подобно многим физиологическим системам контроля, система управления дыханием организована как контур отрицательной обратной связи.
Афферентация с различных рецепторных зон интегрируется в бульбарном дыхательном центре. Последний, в свою очередь, генерирует импульсацию к мотонейронам спинального отдела дыхательного центра, регулирующего сократительную активность дыхательной мускулатуры.
Важная роль в регуляции внешнего дыхания отводится центрам варолиева моста, в частности, пневмотаксическому центру. Последний включает медиальное, парабрахиальное ядро и ядро Келликера. В парабрахиальном ядре находятся преимущественно инспираторные, экспираторные и фазопереходные нейроны. Ядро Келликера содержит инспираторные нейроны.
Дыхательные нейроны моста участвуют в механизмах смены фаз дыхания, регулируют величину дыхательного объема.
Непосредственными регуляторами сократительной способности дыхательных мышц являются спинальные мотонейроны, получающие информацию по нисходящим ретикулоспинальным путям от бульбарного дыхательного центра.
Как известно, нейроны диафрагмального нерва расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Подавляющее количество волокон диафрагмального нерва являются аксонами α-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен диафрагмы, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны, иннервирующие межреберные мышцы, расположены в передних рогах спинного мозга на уровне TIV-TX, из них часть нейронов регулирует сокращения межреберных мышц, а другая часть – их позно-тоническую активность.
Обращает на себя внимание тот факт, что активность спинальных мотонейронов, обеспечивающих регуляцию двигательной активности межреберных мышц и диафрагмы, в свою очередь, находится под контролем инспираторных нейронов спинного мозга, расположенных на уровне СI-CII вблизи латерального края промежуточной зоны серого вещества.
В обеспечении дыхания, особенно в условиях патологии, участвуют мышцы брюшной стенки, получающие иннервацию от мотонейронов спинного мозга на уровне TIV-LIII.
Двум фазам внешнего дыхания (вдоху и выдоху) соответствуют три фазы активности бульбарного дыхательного центра: инспирация, пассивная контролируемая экспирация и активная экспирация. Во время фазы инспирации диафрагма и наружные межреберные мышцы увеличивают силу сокращения, активируются мышцы гортани, расширяется голосовая щель, снижается сопротивление потоку воздуха. В постинспираторную фазу дыхания происходит медленное расслабление диафрагмы, сокращение мышц гортани, выход воздуха в окружающую среду.
В фазе экспирации – экспираторный поток усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки.
Рефлекторная регуляция дыхания обеспечивается за счет афферентной импульсации в бульбарный дыхательный центр с различных рецепторных зон. Мощной рефлексогенной зоной является слизистая оболочка полости носа, где расположены различные типы механорецепторов, в том числе ирритантные, растяжения, а также болевой чувствительности, обоняния.
Возбуждение этих рецепторов возникает в момент каждого вдоха и приводит к формированию потока афферентной импульсации в ретикулярную формацию ствола мозга с последущей активацией бульбарного дыхательного центра, сосудодвигательного центра, гипоталамических и корковых структур мозга.
Раздражение ирритантных рецепторов слизистой оболочки носа приводит к рефлекторному сужению бронхов, голосовой щели, остановке дыхания на выдохе, развитию брадикардии, а в ряде случаев прекращению сердечных сокращений и другим изменениям (тормозной тригемино-вагусный рефлекс Кречмера ).
Слизистая трахеи и бронхов является слабой рефлексогенной зоной. В стенке крупных внелегочных бронхов и трахеи имеются высокопороговые, низкочувствительные медленноадаптирующиеся, быстроадаптирующиеся и промежуточные механорецепторы, в норме их роль в регуляции дыхания минимальна.
Чувствительность этих рецепторов возрастает при развитии воспалительного процесса в бронхолегочной системе инфекционной или аллергической природы, когда освобождаются медиаторы воспаления и аллергии: гистамин, кинины, лейкотриены, простагландины и др.. Возбудимость рецепторов трахеи и бронхов возрастает и в случае застойных явлений в малом кругу кровообращения, когда прежние объемы воздуха сильно растягивают стенки воздухоносных путей. Афферентация с рецепторов трахеи и бронхов направляется в бульбарный дыхательный центр по чувствительным волокнам n. Vagus, модулируя глубину и частоту дыхательных движений.
Мощной рефлексогенной зоной является паренхима легких, обеспечивающая не только альвеолярное дыхание, но и рефлекторную регуляцию внешнего дыхания.
Основные типы легочных вагусных афферентов включают: медленноадаптирующиеся рецепторы растяжения альвеол, быстроадаптирующиеся рецепторы, С-волокна.
Многочисленные быстроадаптирующиеся рецепторы (БАР) находятся в эпителии внутрилегочных бронхов и бронхиол. Эти рецепторы наиболее чувствительны к следующим типам раздражителей: ирритантным воздействиям, повреждению паренхимы и механическому раздражению дыхательных путей. Возбуждение БАР возникает также при глубоком дыхании, легочной эмболии и капиллярной гипертензии. Афферентация с этих рецепторов распространяется по чувствительным маломиелинизированным волокнам n. Vagus в ретикулярную формацию ствола мозга и бульбарный дыхательный центр, вызывая бронхоконстрикцию, тахипноэ, развитие кашля и тахикардии. Возбуждение этих рецепторов может быть клинически значимым в патогенезе бронхиальной астмы и нарушениях реактивности дыхательных путей.
По данным ряда авторов в паренхиме легких выделяют и БАР рецепторы спадения, реагирующие на спадение альвеол под воздействием внутрилегочных и внелегочных факторов. Афферентация с этих рецепторов поступает в бульбарный дыхательный центр по маломиелинизированным волокнам n. Vagus и обеспечивает развитие тахипноэ.
Медленноадаптирующиеся рецепторы растяжения – важная группа механорецепторов c вагусной афферентацией, расположенных в гладких мышцах воздухоносных путей. Частота импульсов с этих рецепторов возрастает по мере растяжения альвеол вдыхаемым воздухом и распространяется по толстым миелинизированным α-волокнам n. Vagus в бульбарный дыхательный отдел, обеспечивая формирование рефлекса Геринга-Брейера. Последний контролирует частоту и глубину дыхания, имеет физиологическое значение при дыхательных объемах превышающих 1 л (у взрослых при физической нагрузке). Рефлекс Геринга-Брейера более важен для регуляции дыхательного акта у новорожденных, а также в условиях патологии как один из механизмов реализации инспираторной, экспираторной и смешанной одышек.
Третьей группой легочных механорецепторов являются С-волокна – тонкие миелинизированные вагусные афференты. С – волокна оканчиваются в паренхиме легких, в бронхах и кровеносных сосудах, активируются экзогенными раздражителями и медиаторами альтерации. Активация С-волокон приводит к тахипноэ, брадикардии, гиперсекреции слизи. В состав С-волокон входят J-рецепторы, расположенные в альвеолярных перегородках в контакте с капиллярами (юкстакапиллярные рецепторы), чувствительные к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам. Активация J-рецепторов вызывает закрытие гортани и апноэ, за которыми следует частое поверхностное дыхание, гипотензия и брадикардия.
Важная роль в рефлекторной регуляции дыхания отводится проприорецепторам суставов грудной клетки, межреберных мышц, диафрагмы, сухожильным рецепторам. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которая через α-мотонейроны повышает активность α-мотонейронов и дозирует таким образом мышечное усилие.
В регуляции активности бульбарного дыхательного центра и внешнего дыхания принимает участие и афферентация с висцеральных рецепторов и рецепторов кожи, о чем свидетельствует развитие гипервентиляции легких при болевом и термическом раздражении.
3.2. Механизмы гуморальной регуляции дыхания
Важная роль в регуляции дыхания отводится хеморецепторам.
Изменения газового состава крови (РаО2, РаСО2) влияют на активность дыхательного центра путем возбуждения хеморецепторов каротидных и аортальных телец (периферические рецепторы), а также хеморецепторов вентральной зоны продолговатого мозга и дорсального дыхательного ядра (центральные рецепторы). Периферические хеморецепторы (рис.5) обеспечивают регуляцию частоты дыхательных движений. Адекватным раздражителем для них является уменьшение РО2 артериальной крови, в меньшей степени – увеличение РСО2 и снижение рН. Периферические хеморецепторы расположены у бифуркации общих сонных артерий на внутреннюю и наружнюю. Несмотря на свой миниатюрный размер, каротидные тельца интенсивно кровоснабжаются (1,4-2 л/мин на 100 г ткани). Этот орган особенно чувствителен к колебаниям кислорода в артериальной крови. При Ра О2 в пределах 60-80 мм рт. ст. наблюдается слабое усиление вентиляции, при Ра О2 ниже 50 мм рт. ст. возникает выраженная гипервентиляция легких. Ра СО2 и рН крови потенцируют эффекты гипоксемии на артериальные хеморецепторы и не являются адекватными раздражителями для этих рецепторов. После двустороннего удаления каротидных телец гипоксический вентиляторный ответ у человека исчезает. При отсутствии хеморецепторной стимуляции, например, при глубокой гипокапнии, повреждении синокаротидной зоны (опухоли, коллагенозы, травмы) ритмогенез дыхания снижается и полностью прекращается.
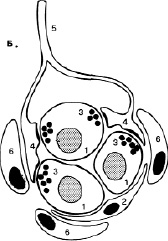
Рис. 5. Каротидное тельце: 1-хеморецепторные клетки; 2-поддерживающие клетки; 3-синаптические пузырьки; 4-чувствительные нервные окончания; 5-нервное волокно
Центральные хемочувствительные клетки реагируют на отклонения РСО2 и [H+] во внеклеточной жидкости внутримозгового интерстициального пространства, регулируют глубину вдоха. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Одной из причин высокой скорости вентиляторного ответа на гиперкапнию является легкость диффузии СО2 через барьерную систему кровь-головной мозг. Более того, повышенное РСО2 вызывает расширение сосудов, особенно церебральных, способствуя тем самым усилению диффузии СО2 через гемато-энцефалический барьер.
Библиографическая ссылка
Чеснокова Н.П, Брилль Г.Е., Моррисон В.В., Понукалина Е.В., Полутова Н.В. ЛЕКЦИЯ 3 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ ДЫХАНИЯ // Научное обозрение. Медицинские науки. – 2017. – № 2.
– С. 36-39;
URL: https://science-medicine.ru/ru/article/view?id=972 (дата обращения: 03.04.2023).
Предлагаем вашему вниманию журналы, издающиеся в издательстве «Академия Естествознания»
(Высокий импакт-фактор РИНЦ, тематика журналов охватывает все научные направления)
| Двигательный нейрон | |
|---|---|

Микрофотография из подъязычное ядро показывая мотонейроны с их характерными грубыми Субстанция Ниссля («тигроидная» цитоплазма). Окраска H & E-LFB. |
|
| подробности | |
| Расположение | Вентральный рог из спинной мозг, немного ядра черепных нервов |
| Форма | Проекционный нейрон |
| Функция | Возбуждающая проекция (на СМП ) |
| Нейротрансмиттер | UMN к LMN: глутамат; LMN к СМП: АЧ |
| Пресинаптические связи | Первичная моторная кора через Кортикоспинальный тракт |
| Постсинаптические связи | Мышечные волокна и другие нейроны |
| Идентификаторы | |
| MeSH | D009046 |
| НейроЛекс МНЕ БЫ | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Анатомические термины нейроанатомии
[редактировать в Викиданных ] |
А двигательный нейрон (или мотонейрон) это нейрон чья Тело клетки расположен в моторная кора, мозговой ствол или спинной мозг, и чья аксон (волокна) проецируются в спинной мозг или за пределы спинного мозга, чтобы прямо или косвенно управлять эффекторными органами, в основном мышцы и железы.[1] Есть два типа моторных нейронов — верхние двигательные нейроны и нижние двигательные нейроны. Аксоны из синапса верхних мотонейронов на интернейроны в спинном мозге и иногда прямо на нижние мотонейроны.[2] Аксоны нижних мотонейронов эфферентные нервные волокна которые передают сигналы от спинной мозг к эффекторам.[3] Типы нижних мотонейронов: альфа двигательные нейроны, бета мотонейроны, и гамма мотонейроны.
Один двигательный нейрон может иннервировать множество мышечные волокна и мышечное волокно может подвергнуться многим потенциалы действия за время, затраченное на сингл подергивание мышц. Иннервация происходит в нервно-мышечное соединение и подергивания могут накладываться друг на друга в результате суммирование или тетаническое сокращение. Отдельные подергивания могут стать неразличимыми, а напряжение плавно нарастает, в конечном итоге достигая плато.[4]
Развитие
Моторные нейроны начинают развиваться рано эмбриональное развитие, и двигательная функция продолжает развиваться в детстве.[5] в нервная трубка клетки специфицируются либо к рострально-каудальной оси, либо к вентрально-дорсальной оси. В аксоны мотонейронов начинают появляться на четвертой неделе развития из вентральной области вентрально-дорсальной оси ( базальная пластинка ).[6] Этот гомеодомен известен как домен предшественника моторных нейронов (pMN). Факторы транскрипции сюда включают Pax6, OLIG2, Nkx-6.1, и НКХ-6.2, которые регулируются звуковой еж (Тсс). Ген OLIG2 является наиболее важным из-за его роли в продвижении Выражение Ngn2, ген, который вызывает выход из клеточного цикла, а также способствует дальнейшим факторам транскрипции, связанным с развитием моторных нейронов.[7]
Дальнейшая спецификация мотонейронов происходит, когда ретиноевая кислота, фактор роста фибробластов, Wnts, и TGFb, интегрированы в различные Hox факторы транскрипции. Существует 13 факторов транскрипции Hox, которые вместе с сигналами определяют, будет ли мотонейрон более ростральным или каудальным по характеру. В позвоночнике Hox 4-11 сортируют мотонейроны в один из пяти моторных столбцов.[7]
| Моторная колонка | Расположение в спинном мозге | Цель |
| Средняя колонка двигателя | Присутствует вся длина | Осевые мышцы |
| Гипаксиальная моторная колонка | Грудной отдел | Мышцы стенки тела |
| Преганглионарная моторная колонка | Грудной отдел | Симпатический ганглий |
| Боковая моторная колонка | Плечевая и поясничная области (обе области далее делятся на медиальную и латеральную области) | Мышцы конечностей |
| Диафрагмальная моторная колонка | Шейный отдел | Диафрагма[9] |
Анатомия и психология

Тракты спинного мозга

Расположение нижних мотонейронов в спинном мозге
Верхние двигательные нейроны
Верхние мотонейроны происходят из моторная кора расположен в прецентральная извилина. Клетки, составляющие первичная моторная кора находятся Клетки Беца, которые являются разновидностью пирамидная ячейка. Аксоны этих клеток спускаются из коры, чтобы сформировать кортикоспинальный тракт.[10] Кортикомоторнейроны проецируются из первичной коры непосредственно на двигательные нейроны вентрального рога спинного мозга.[11][12] Их аксоны синапсы на спинных мотонейронах нескольких мышц, а также на спинномозговой интернейроны.[11][12] Они уникальны для приматов, и было высказано предположение, что их функция — адаптивное управление Руки включая относительно независимое управление отдельными пальцами.[12][13] Кортикомоторнейроны до сих пор были обнаружены только в первичной моторной коре, но не во вторичных моторных областях.[12]
Нервные тракты
Нервные тракты пучки аксонов как белое вещество, которые несут потенциалы действия к их эффекторам. В спинном мозге эти нисходящие пути переносят импульсы из разных регионов. Эти тракты также служат местом происхождения нижних мотонейронов. В спинном мозге можно обнаружить семь основных нисходящих моторных трактов:[14]
- Боковой кортикоспинальный тракт
- Руброспинальный тракт
- Боковой ретикулоспинальный тракт
- Вестибулоспинальный тракт
- Медиальный ретикулоспинальный тракт
- Тектоспинальный тракт
- Передний кортикоспинальный тракт
Нижние двигательные нейроны
Нижние мотонейроны — это те, которые берут начало в спинном мозге и прямо или косвенно иннервируют эффекторные мишени. Мишени этих нейронов различны, но в соматической нервной системе целью будет какое-то мышечное волокно. Существует три основных категории низших мотонейронов, которые можно разделить на подкатегории.[15]
В соответствии с их целями моторные нейроны делятся на три широкие категории:[16]
- Соматические двигательные нейроны
- Специальные висцеральные двигательные нейроны
- Общие висцеральные двигательные нейроны
Соматические двигательные нейроны
Соматические мотонейроны происходят из Центральная нервная система, проецировать их аксоны к скелетные мышцы [17] (например, мышцы конечностей, брюшного пресса и межреберные мышцы ), которые участвуют в движение. Три типа этих нейронов — это альфа эфферентные нейроны, бета эфферентные нейроны, и гамма эфферентные нейроны. Они называются эфферент для обозначения потока информации от Центральная нервная система (CNS) в периферия.
- Альфа двигательные нейроны иннервировать экстрафузальные мышечные волокна, которые являются основным компонентом мышцы, генерирующим силу. Их клеточные тела находятся в брюшной рог спинного мозга, и их иногда называют клетки вентрального рога. Один мотонейрон может синапсировать в среднем со 150 мышечными волокнами.[18] Моторный нейрон и все мышечные волокна, с которыми он соединяется, представляют собой моторный блок. Моторные агрегаты делятся на 3 категории:[19] Основная статья: Моторный блок
- Медленные (S) двигательные единицы стимулируют мелкие мышечные волокна, которые сокращаются очень медленно и дают небольшое количество энергии, но очень устойчивы к утомлению, поэтому они используются для поддержания мышечного сокращения, например, для поддержания тела в вертикальном положении. Они получают энергию с помощью окислительных средств и, следовательно, нуждаются в кислороде. Их еще называют красными волокнами.[19]
- Двигательные единицы с быстрым утомлением (FF) стимулируют большие группы мышц, которые прикладывают большое количество силы, но очень быстро утомляются. Они используются для задач, требующих больших коротких всплесков энергии, таких как прыжки или бег. Они получают энергию с помощью гликолитических средств и, следовательно, не нуждаются в кислороде. Их называют белыми волокнами.[19]
- Быстрые устойчивые к утомлению двигательные единицы стимулируют группы мышц среднего размера, которые не реагируют так быстро, как двигательные единицы FF, но могут удерживаться намного дольше (как следует из названия) и обеспечивают большую силу, чем двигательные единицы S. Они используют как окислительные, так и гликолитические средства для получения энергии.[19]
Помимо произвольного сокращения скелетных мышц, альфа-мотонейроны также способствуют мышечный тонус, непрерывная сила, создаваемая несокращающейся мышцей, препятствующая растяжению. Когда мышца растягивается, сенсорные нейроны в пределах мышечное веретено определить степень растяжения и отправить сигнал в ЦНС. ЦНС активирует альфа-двигательные нейроны в спинном мозге, которые заставляют экстрафузионные мышечные волокна сокращаться и тем самым сопротивляться дальнейшему растяжению. Этот процесс также называют рефлекс растяжения.
- Бета мотонейроны иннервировать интрафузальные мышечные волокна из мышечные веретена, с коллатералями к экстрафузальным волокнам. Есть два типа бета-мотонейронов: Медленно сокращающиеся — они иннервируют экстрафузальные волокна. Быстрое сокращение — они иннервируют интрафузальные волокна.[20]
- Гамма двигательные нейроны иннервируют интрафузальные мышечные волокна в мышечном веретене. Они регулируют чувствительность веретена к растяжению мышц. При активации гамма-нейронов интрафузальные мышечные волокна сокращаются, так что требуется лишь небольшое растяжение для активации сенсорных нейронов веретена и рефлекса растяжения. Существует два типа гамма-мотонейронов: динамические — они фокусируются на волокнах Bag1 и повышают динамическую чувствительность. Статические — они фокусируются на волокнах Bag2 и повышают чувствительность к растяжению.[20]
- Регулирующие факторы нижних мотонейронов
- Принцип размера — это относится к соме мотонейрона. Это ограничивает более крупные нейроны для получения большего возбуждающего сигнала для стимуляции мышечных волокон, которые они иннервируют. Уменьшая ненужное задействование мышечных волокон, организм может оптимизировать потребление энергии.[20]
- Постоянный входящий ток (PIC) — недавние исследования на животных показали, что постоянный поток ионов, таких как кальций и натрий, через каналы в соме и дендритах влияет на синаптический вход. Другой способ думать об этом заключается в том, что постсинаптический нейрон активируется перед получением импульса.[20]
- После Гиперполяризация (AHP) — Обнаружена тенденция, согласно которой медленные двигательные нейроны имеют более интенсивные AHP в течение более длительного времени. Один из способов запомнить это — то, что медленные мышечные волокна могут сокращаться дольше, поэтому имеет смысл, что соответствующие им двигательные нейроны активизируются с меньшей скоростью.[20]
Специальные висцеральные двигательные нейроны
Они также известны как жаберные двигательные нейроны, которые участвуют в мимике, жевании, фонации и глотании. Связанные черепные нервы — это глазодвигательный, похищает, трохлеарный, и подъязычный нервы.[16]
| Филиал НС | Должность | Нейротрансмиттер |
|---|---|---|
| Соматический | н / д | Ацетилхолин |
| Парасимпатический | Преганглионарный | Ацетилхолин |
| Парасимпатический | Ганглионарный | Ацетилхолин |
| Симпатичный | Преганглионарный | Ацетилхолин |
| Симпатичный | Ганглионарный | Норэпинефрин * |
| * За исключением волокон до потовые железы и некоторые кровеносный сосуд Нейромедиаторы моторных нейронов |
Общие висцеральные двигательные нейроны
Эти двигательные нейроны косвенно иннервируют сердечная мышца и гладкие мышцы из внутренности (мышцы артерии ): Oни синапс на нейроны, расположенные в ганглии из автономная нервная система (симпатичный и парасимпатический ), расположенный в периферическая нервная система (ПНС), которые непосредственно иннервируют висцеральные мышцы (а также некоторые клетки железы).
Как следствие, моторная команда скелетный и жаберные мышцы моносинаптический с участием только одного мотонейрона, либо соматический или жаберный, который прикрепляется к мышце. Для сравнения, командование висцеральные мышцы является дисинаптический с участием двух нейронов: общий висцеральный мотонейрон, расположенный в ЦНС, синапсирует с ганглиозным нейроном, расположенным в ПНС, который синапсирует с мышцей.
Все двигательные нейроны позвоночных холинергический, то есть они выпускают нейротрансмиттер ацетилхолин. Парасимпатические ганглиозные нейроны также являются холинэргическими, тогда как большинство симпатических ганглиозных нейронов являются холинергическими. норадренергический, то есть они высвобождают нейромедиатор норадреналин. (см. таблицу)
Нервно-мышечные соединения
Один двигательный нейрон может иннервировать множество мышечные волокна и мышечное волокно может подвергнуться многим потенциалы действия за время, затраченное на сингл подергивание мышц. В результате, если потенциал действия возникает до того, как подергивание завершилось, подергивания могут накладываться друг на друга либо через суммирование или тетаническое сокращение. В итоге, мышца стимулируется повторно, так что дополнительные потенциалы действия, исходящие от соматическая нервная система прибыть до окончания подергивания. Таким образом, подергивания накладываются друг на друга, что приводит к большей силе, чем при одиночном подергивании. Тетаническое сокращение вызывается постоянной, очень высокочастотной стимуляцией — потенциалы действия возникают с такой высокой скоростью, что отдельные подергивания неразличимы, а напряжение плавно нарастает, в конечном итоге достигая плато.[4]
Интерфейс между двигательным нейроном и мышечным волокном — это специализированный синапс называется нервно-мышечное соединение. При адекватной стимуляции мотонейрон выпускает поток ацетилхолина (Ach). нейротрансмиттеры от окончаний аксона из синаптических пузырьков связываются с плазматической мембраной. Молекулы ацетилхолина связываются с постсинаптический рецепторы находится в концевой пластине двигателя. После связывания двух рецепторов ацетилхолина открывается ионный канал, и ионы натрия могут проникать в клетку. Приток натрия в клетку вызывает деполяризацию и запускает мышечный потенциал действия. Затем стимулируются Т-канальцы сарколеммы, чтобы вызвать высвобождение ионов кальция из саркоплазматического ретикулума. Именно это химическое высвобождение заставляет целевое мышечное волокно сокращаться.[18]
В беспозвоночные В зависимости от высвобождаемого нейротрансмиттера и типа рецептора, который он связывает, реакция мышечного волокна может быть либо возбуждающей, либо тормозящей. Для позвоночные Однако ответ мышечного волокна на нейротрансмиттер может быть только возбуждающим, другими словами, сократительным. Расслабление мышц и подавление мышечных сокращений у позвоночных достигается только путем подавления самого двигательного нейрона. Вот как миорелаксанты работают, воздействуя на двигательные нейроны, которые иннервируют мышцы (уменьшая их электрофизиологический активность) или на холинергический нервно-мышечные соединения, а не сами мышцы.
Смотрите также
- Ячейка Беца
- Центральный хроматолиз
- Двигательная дисфункция
- Заболевание двигательных нейронов
- Нерв
использованная литература
- ^ Тортора, Жерар; Дерриксон, Брайан (2014). Принципы анатомии и физиологии (14-е изд.). Нью-Джерси: John Wiley & Sons, Inc., стр.406, 502, 541. ISBN 978-1-118-34500-9.
- ^ Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Оксфорд: Издательство Оксфордского университета. С. 151–153. ISBN 978-0-19-856878-0.
- ^ Шактер Д.Л., Гилберт Д.Т., Вегнер Д.М. (2011) Психология второе издание. Нью-Йорк, Нью-Йорк: стоит
- ^ а б Рассел, Питер (2013). Биология — изучение разнообразия жизни. Торонто: образование Нельсона. п. 946. ISBN 978-0-17-665133-6.
- ^ Тортора, Жерар; Дерриксон, Брайан (2011). Принципы анатомической физиологии (14-е изд.). Нью-Джерси: John Wiley & Sons, Inc., стр.1090–1099. ISBN 978-1-118-34500-9.
- ^ Сэдлер, Т. (2010). Медицинская эмбриология Лангмана (11-е изд.). Филадельфия: Липпинкотт Уильям и Уилкинс. С. 299–301. ISBN 978-0-7817-9069-7.
- ^ а б Дэвис-Дузенбери, Б.Н.; Уильямс, Луизиана; Клим, младший; Эгган, К. (февраль 2014 г.). «Как сделать спинномозговые мотонейроны». Развитие. 141 (3): 491–501. Дои:10.1242 / дев.097410. PMID 24449832.
- ^ Эдгар Р., Мазор Й, Ринон А., Блюменталь Дж., Голан Й, Бужор Э, Ливнат I, Бен-Ари С, Лидер I, Шитрит А, Гильбоа Й, Бен-Иегуда А, Эдри О, Шрага Н., Богоч Й, Лешанский Л., Ахарони С., Западный доктор медицины, Варшавский Д., Штрихман Р. (2013). «LifeMap Discovery ™: портал исследований эмбрионального развития, стволовых клеток и регенеративной медицины». PLoS ONE. 8 (7): e66629. Bibcode:2013PLoSO … 866629E. Дои:10.1371 / journal.pone.0066629. ISSN 1932-6203. ЧВК 3714290. PMID 23874394.
- ^ Филиппиду, Поликсени; Уолш, Кэролайн; Обен, Жозе; Жаннотта, Люси; Дасен, Джереми С. (2012). «Устойчивая активность гена Hox5 необходима для развития дыхательных моторных нейронов». Природа Неврология. 15 (12): 1636–1644. Дои:10.1038 / нн.3242. ISSN 1097-6256. ЧВК 3676175. PMID 23103965.
- ^ Фитцпатрик, Д. (2001) Первичная моторная кора: верхние моторные нейроны, которые инициируют сложные произвольные движения. В D. Purves, G.J. Августин, Д. Фитцпатрик и др. (Ред.), Неврология. Полученное из «Архивная копия». В архиве из оригинала 2018-06-05. Получено 2017-11-30.CS1 maint: заархивированная копия как заголовок (ссылка на сайт)
- ^ а б Мак, Сара; Кандел, Эрик Р .; Джесселл, Томас М .; Шварц, Джеймс Х .; Siegelbaum, Steven A .; Хадспет, А. Дж. (2013). Принципы нейронауки. Кандел, Эрик Р. (5-е изд.). Нью-Йорк. ISBN 9780071390118. OCLC 795553723.
- ^ а б c d Лимон, Роджер Н. (4 апреля 2008 г.). «Нисходящие пути в управлении моторикой». Ежегодный обзор нейробиологии. 31 (1): 195–218. Дои:10.1146 / annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853.
- ^ Иса, Т. (апрель 2007 г.). «Прямые и непрямые кортико-мотонейрональные пути и контроль движений кисти / руки». Физиология. 22 (2): 145–152. Дои:10.1152 / Physiol.00045.2006. PMID 17420305.
- ^ Тортора, Дж. Дж., Дерриксон, Б. (2011). Спинной мозг и спинномозговые нервы. В B. Roesch, L. Elfers, K. Trost, et al. (Ред.), Основы анатомии и физиологии (стр. 443-468). Нью-Джерси: John Wiley & Sons, Inc.
- ^ Фитцпатрик, Д. (2001) Цепи нижних моторных нейронов и управление моторами: Обзор. В D. Purves, G.J. Августин, Д. Фитцпатрик и др. (Ред.), Неврология. Полученное из «Архивная копия». В архиве из оригинала 2018-06-05. Получено 2017-11-30.CS1 maint: заархивированная копия как заголовок (ссылка на сайт)
- ^ а б «ГЛАВА ДЕВЯТАЯ». www.unc.edu. В архиве из оригинала на 2017-11-05. Получено 2017-12-08.
- ^ Сильверторн, Ди Англауб (2010). Физиология человека: комплексный подход. Пирсон. п. 398. ISBN 978-0-321-55980-7.
- ^ а б Тортора, Дж. Дж., Дерриксон, Б. (2011). Мышечная ткань. В B. Roesch, L. Elfers, K. Trost, et al. (Ред.), Основы анатомии и физиологии (стр. 305-307, 311). Нью-Джерси: John Wiley & Sons, Inc.
- ^ а б c d Purves D, Augustine GJ, Fitzpatrick D и др., Редакторы: Neuroscience. 2-е издание, 2001 г. «Архивная копия». В архиве из оригинала 2018-06-05. Получено 2017-09-05.CS1 maint: заархивированная копия как заголовок (ссылка на сайт)
- ^ а б c d е Мануэль, Марин; Житницкий, Даниэль (2011). «Альфа, бета и гамма мотонейроны: функциональное разнообразие на последнем пути двигательной системы». Журнал интегративной неврологии. 10 (3): 243–276. Дои:10.1142 / S0219635211002786. ISSN 0219-6352. PMID 21960303.
Источники
- Шервуд, Л. (2001). Физиология человека: от клеток к системам (4-е изд.). Пасифик Гроув, Калифорния: Брукс-Коул. ISBN 0-534-37254-6.
- Marieb, E.N .; Маллатт, Дж. (1997). Человеческая анатомия (2-е изд.). Менло-Парк, Калифорния: Бенджамин / Каммингс. ISBN 0-8053-4068-8.
From Wikipedia, the free encyclopedia
| Motor neurons | |
|---|---|
|
Micrograph of the hypoglossal nucleus showing motor neurons with their characteristic coarse Nissl substance («tigroid» cytoplasm). H&E-LFB stain. |
|
| Details | |
| Location | Ventral horn of the spinal cord, some cranial nerve nuclei |
| Shape | Projection neuron |
| Function | Excitatory projection (to NMJ) |
| Neurotransmitter | UMN to LMN: glutamate; LMN to NMJ: ACh |
| Presynaptic connections | Primary motor cortex via the Corticospinal tract |
| Postsynaptic connections | Muscle fibers and other neurons |
| Identifiers | |
| MeSH | D009046 |
| NeuroLex ID | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A motor neuron (or motoneuron or efferent neuron[1]) is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands.[2] There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons.[3] The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors.[4] Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. Innervation takes place at a neuromuscular junction and twitches can become superimposed as a result of summation or a tetanic contraction. Individual twitches can become indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
Development[edit]
Motor neurons begin to develop early in embryonic development, and motor function continues to develop well into childhood.[6] In the neural tube cells are specified to either the rostral-caudal axis or ventral-dorsal axis. The axons of motor neurons begin to appear in the fourth week of development from the ventral region of the ventral-dorsal axis (the basal plate).[7] This homeodomain is known as the motor neural progenitor domain (pMN). Transcription factors here include Pax6, OLIG2, Nkx-6.1, and Nkx-6.2, which are regulated by sonic hedgehog (Shh). The OLIG2 gene being the most important due to its role in promoting Ngn2 expression, a gene that causes cell cycle exiting as well as promoting further transcription factors associated with motor neuron development.[8]
Further specification of motor neurons occurs when retinoic acid, fibroblast growth factor, Wnts, and TGFb, are integrated into the various Hox transcription factors. There are 13 Hox transcription factors and along with the signals, determine whether a motor neuron will be more rostral or caudal in character. In the spinal column, Hox 4-11 sort motor neurons to one of the five motor columns.[8]
| Motor column | Location in spinal cord | Target |
| Median motor column | Present entire length | Axial muscles |
| Hypaxial motor column | Thoracic region | Body wall muscles |
| Preganglionic motor column | Thoracic region | Sympathetic ganglion |
| Lateral motor column | Brachial and lumbar region (both regions are further divided into medial and lateral domains) | Muscles of the limbs |
| Phrenic motor column | Cervical region | Diaphragm[10] |
Anatomy and physiology[edit]
Location of lower motor neurons in spinal cord
Upper motor neurons[edit]
Upper motor neurons originate in the motor cortex located in the precentral gyrus. The cells that make up the primary motor cortex are Betz cells, which are giant pyramidal cells. The axons of these cells descend from the cortex to form the corticospinal tract.[11] Corticomotorneurons project from the primary cortex directly onto motor neurons in the ventral horn of the spinal cord.[12][13] Their axons synapse on the spinal motor neurons of multiple muscles as well as on spinal interneurons.[12][13] They are unique to primates and it has been suggested that their function is the adaptive control of the hands including the relatively independent control of individual fingers.[13][14] Corticomotorneurons have so far only been found in the primary motor cortex and not in secondary motor areas.[13]
Nerve tracts[edit]
Nerve tracts are bundles of axons as white matter, that carry action potentials to their effectors. In the spinal cord these descending tracts carry impulses from different regions. These tracts also serve as the place of origin for lower motor neurons. There are seven major descending motor tracts to be found in the spinal cord:[15]
- Lateral corticospinal tract
- Rubrospinal tract
- Lateral reticulospinal tract
- Vestibulospinal tract
- Medial reticulospinal tract
- Tectospinal tract
- Anterior corticospinal tract
Lower motor neurons[edit]
Lower motor neurons are those that originate in the spinal cord and directly or indirectly innervate effector targets. The target of these neurons varies, but in the somatic nervous system the target will be some sort of muscle fiber. There are three primary categories of lower motor neurons, which can be further divided in sub-categories.[16]
According to their targets, motor neurons are classified into three broad categories:[17]
- Somatic motor neurons
- Special visceral motor neurons
- General visceral motor neurons
Somatic motor neurons[edit]
Somatic motor neurons originate in the central nervous system, project their axons to skeletal muscles[18] (such as the muscles of the limbs, abdominal, and intercostal muscles), which are involved in locomotion. The three types of these neurons are the alpha efferent neurons, beta efferent neurons, and gamma efferent neurons. They are called efferent to indicate the flow of information from the central nervous system (CNS) to the periphery.
- Alpha motor neurons innervate extrafusal muscle fibers, which are the main force-generating component of a muscle. Their cell bodies are in the ventral horn of the spinal cord and they are sometimes called ventral horn cells. A single motor neuron may synapse with 150 muscle fibers on average.[19] The motor neuron and all of the muscle fibers to which it connects is a motor unit. Motor units are split up into 3 categories:[20]
- Slow (S) motor units stimulate small muscle fibers, which contract very slowly and provide small amounts of energy but are very resistant to fatigue, so they are used to sustain muscular contraction, such as keeping the body upright. They gain their energy via oxidative means and hence require oxygen. They are also called red fibers.[20]
- Fast fatiguing (FF) motor units stimulate larger muscle groups, which apply large amounts of force but fatigue very quickly. They are used for tasks that require large brief bursts of energy, such as jumping or running. They gain their energy via glycolytic means and hence don’t require oxygen. They are called white fibers.[20]
- Fast fatigue-resistant motor units stimulate moderate-sized muscles groups that don’t react as fast as the FF motor units, but can be sustained much longer (as implied by the name) and provide more force than S motor units. These use both oxidative and glycolytic means to gain energy.[20]
In addition to voluntary skeletal muscle contraction, alpha motor neurons also contribute to muscle tone, the continuous force generated by noncontracting muscle to oppose stretching. When a muscle is stretched, sensory neurons within the muscle spindle detect the degree of stretch and send a signal to the CNS. The CNS activates alpha motor neurons in the spinal cord, which cause extrafusal muscle fibers to contract and thereby resist further stretching. This process is also called the stretch reflex.
- Beta motor neurons innervate intrafusal muscle fibers of muscle spindles, with collaterals to extrafusal fibres. There are two types of beta motor neurons: Slow Contracting- These innervate extrafusal fibers. Fast Contracting- These innervate intrafusal fibers.[21]
- Gamma motor neurons innervate intrafusal muscle fibers found within the muscle spindle. They regulate the sensitivity of the spindle to muscle stretching. With activation of gamma neurons, intrafusal muscle fibers contract so that only a small stretch is required to activate spindle sensory neurons and the stretch reflex. There are two types of gamma motor neurons: Dynamic- These focus on Bag1 fibers and enhance dynamic sensitivity. Static- These focus on Bag2 fibers and enhance stretch sensitivity.[21]
- Regulatory factors of lower motor neurons
- Size Principle – this relates to the soma of the motor neuron. This restricts larger neurons to receive a larger excitatory signal in order to stimulate the muscle fibers it innervates. By reducing unnecessary muscle fiber recruitment, the body is able to optimize energy consumption.[21]
- Persistent Inward Current (PIC) – recent animal study research has shown that constant flow of ions such as calcium and sodium through channels in the soma and dendrites influence the synaptic input. An alternate way to think of this is that the post-synaptic neuron is being primed before receiving an impulse.[21]
- After Hyper-polarization (AHP) – A trend has been identified that shows slow motor neurons to have more intense AHPs for a longer duration. One way to remember this is that slow muscle fibers can contract for longer, so it makes sense that their corresponding motor neurons fire at a slower rate.[21]
Special visceral motor neurons[edit]
These are also known as branchial motor neurons, which are involved in facial expression, mastication, phonation, and swallowing. Associated cranial nerves are the oculomotor, abducens, trochlear, and hypoglossal nerves.[17]
| Branch of NS | Position | Neurotransmitter |
|---|---|---|
| Somatic | n/a | Acetylcholine |
| Parasympathetic | Preganglionic | Acetylcholine |
| Parasympathetic | Ganglionic | Acetylcholine |
| Sympathetic | Preganglionic | Acetylcholine |
| Sympathetic | Ganglionic | Norepinephrine* |
| *Except fibers to sweat glands and certain blood vessels Motor neuron neurotransmitters |
General visceral motor neurons[edit]
These motor neurons indirectly innervate cardiac muscle and smooth muscles of the viscera ( the muscles of the arteries): they synapse onto neurons located in ganglia of the autonomic nervous system (sympathetic and parasympathetic), located in the peripheral nervous system (PNS), which themselves directly innervate visceral muscles (and also some gland cells).
In consequence, the motor command of skeletal and branchial muscles is monosynaptic involving only one motor neuron, either somatic or branchial, which synapses onto the muscle. Comparatively, the command of visceral muscles is disynaptic involving two neurons: the general visceral motor neuron, located in the CNS, synapses onto a ganglionic neuron, located in the PNS, which synapses onto the muscle.
All vertebrate motor neurons are cholinergic, that is, they release the neurotransmitter acetylcholine. Parasympathetic ganglionic neurons are also cholinergic, whereas most sympathetic ganglionic neurons are noradrenergic, that is, they release the neurotransmitter noradrenaline. (see Table)
Neuromuscular junctions[edit]
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. As a result, if an action potential arrives before a twitch has completed, the twitches can superimpose on one another, either through summation or a tetanic contraction. In summation, the muscle is stimulated repetitively such that additional action potentials coming from the somatic nervous system arrive before the end of the twitch. The twitches thus superimpose on one another, leading to a force greater than that of a single twitch. A tetanic contraction is caused by constant, very high frequency stimulation — the action potentials come at such a rapid rate that individual twitches are indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
The interface between a motor neuron and muscle fiber is a specialized synapse called the neuromuscular junction. Upon adequate stimulation, the motor neuron releases a flood of acetylcholine (Ach) neurotransmitters from the axon terminals from synaptic vesicles bind with the plasma membrane. The acetylcholine molecules bind to postsynaptic receptors found within the motor end plate. Once two acetylcholine receptors have been bound, an ion channel is opened and sodium ions are allowed to flow into the cell. The influx of sodium into the cell causes depolarization and triggers a muscle action potential. T tubules of the sarcolemma are then stimulated to elicit calcium ion release from the sarcoplasmic reticulum. It is this chemical release that causes the target muscle fiber to contract.[19]
In invertebrates, depending on the neurotransmitter released and the type of receptor it binds, the response in the muscle fiber could be either excitatory or inhibitory. For vertebrates, however, the response of a muscle fiber to a neurotransmitter can only be excitatory, in other words, contractile. Muscle relaxation and inhibition of muscle contraction in vertebrates is obtained only by inhibition of the motor neuron itself. This is how muscle relaxants work by acting on the motor neurons that innervate muscles (by decreasing their electrophysiological activity) or on cholinergic neuromuscular junctions, rather than on the muscles themselves.
See also[edit]
- Betz cell
- Central chromatolysis
- Motor dysfunction
- Motor neuron disease
- Nerve
- Efferent nerve fiber
- Motor nerve
References[edit]
- ^ «Afferent vs. Efferent: AP® Psych Crash Course Review | Albert.io». Albert Resources. 2019-12-02. Retrieved 2021-04-25.
- ^ Tortora, Gerard; Derrickson, Bryan (2014). Principles of Anatomy & Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 406, 502, 541. ISBN 978-1-118-34500-9.
- ^ Pocock, Gillian; Richards, Christopher D. (2006). Human physiology : the basis of medicine (3rd ed.). Oxford: Oxford University Press. pp. 151–153. ISBN 978-0-19-856878-0.
- ^ Schacter D.L., Gilbert D.T., and Wegner D.M. (2011) Psychology second edition. New York, NY: Worth
- ^ a b Russell, Peter (2013). Biology — Exploring the Diversity of Life. Toronto: Nelson Education. p. 946. ISBN 978-0-17-665133-6.
- ^ Tortora, Gerard; Derrickson, Bryan (2011). Principles of Anatomy Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 1090–1099. ISBN 978-1-118-34500-9.
- ^ Sadler, T. (2010). Langman’s medical embryology (11th ed.). Philadelphia: Lippincott William & Wilkins. pp. 299–301. ISBN 978-0-7817-9069-7.
- ^ a b Davis-Dusenbery, BN; Williams, LA; Klim, JR; Eggan, K (February 2014). «How to make spinal motor neurons». Development. 141 (3): 491–501. doi:10.1242/dev.097410. PMID 24449832.
- ^ Edgar R, Mazor Y, Rinon A, Blumenthal J, Golan Y, Buzhor E, Livnat I, Ben-Ari S, Lieder I, Shitrit A, Gilboa Y, Ben-Yehudah A, Edri O, Shraga N, Bogoch Y, Leshansky L, Aharoni S, West MD, Warshawsky D, Shtrichman R (2013). «LifeMap Discovery™: The Embryonic Development, Stem Cells, and Regenerative Medicine Research Portal». PLOS ONE. 8 (7): e66629. Bibcode:2013PLoSO…866629E. doi:10.1371/journal.pone.0066629. ISSN 1932-6203. PMC 3714290. PMID 23874394.
- ^ Philippidou, Polyxeni; Walsh, Carolyn; Aubin, Josée; Jeannotte, Lucie; Dasen, Jeremy S. (2012). «Sustained Hox5 Gene Activity is Required for Respiratory Motor Neuron Development». Nature Neuroscience. 15 (12): 1636–1644. doi:10.1038/nn.3242. ISSN 1097-6256. PMC 3676175. PMID 23103965.
- ^ Fitzpatrick, D. (2001) The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b Mack, Sarah; Kandel, Eric R.; Jessell, Thomas M.; Schwartz, James H.; Siegelbaum, Steven A.; Hudspeth, A. J. (2013). Principles of neural science. Kandel, Eric R. (5th ed.). New York. ISBN 9780071390118. OCLC 795553723.
- ^ a b c d Lemon, Roger N. (April 4, 2008). «Descending Pathways in Motor Control». Annual Review of Neuroscience. 31 (1): 195–218. doi:10.1146/annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853. S2CID 16139768.
- ^ Isa, T (April 2007). «Direct and indirect cortico-motoneuronal pathways and control of hand/arm movements». Physiology. 22 (2): 145–152. doi:10.1152/physiol.00045.2006. PMID 17420305.
- ^ Tortora, G. J., Derrickson, B. (2011). The Spinal Cord and Spinal Nerves. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 443-468). New Jersey: John Wiley & Sons, Inc.
- ^ Fitzpatrick, D. (2001) Lower Motor Neuron Circuits and Motor Control: Overview. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «Lower Motor Neuron Circuits and Motor Control — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b «CHAPTER NINE». www.unc.edu. Archived from the original on 2017-11-05. Retrieved 2017-12-08.
- ^ Silverthorn, Dee Unglaub (2010). Human Physiology: An Integrated Approach. Pearson. p. 398. ISBN 978-0-321-55980-7.
- ^ a b Tortora, G. J., Derrickson, B. (2011). Muscular Tissue. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 305-307, 311). New Jersey: John Wiley & Sons, Inc.
- ^ a b c d Purves D, Augustine GJ, Fitzpatrick D, et al., editors: Neuroscience. 2nd edition, 2001 «The Motor Unit — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-09-05.
- ^ a b c d e Manuel, Marin; Zytnicki, Daniel (2011). «Alpha, Beta, and Gamma Motoneurons: Functional Diversity in the Motor System’s Final Pathway». Journal of Integrative Neuroscience. 10 (3): 243–276. doi:10.1142/S0219635211002786. ISSN 0219-6352. PMID 21960303. S2CID 21582283.
Sources[edit]
- Sherwood, L. (2001). Human Physiology: From Cells to Systems (4th ed.). Pacific Grove, CA: Brooks-Cole. ISBN 0-534-37254-6.
- Marieb, E. N.; Mallatt, J. (1997). Human Anatomy (2nd ed.). Menlo Park, CA: Benjamin/Cummings. ISBN 0-8053-4068-8.